铜和镁离子对卵圆卡盾藻生长、光合色素和溶血活性的影响
2021-08-17苟偲钰叶志林江天久
苟偲钰,叶志林,江天久
(暨南大学赤潮与海洋生物学研究中心、水体富营养化与赤潮防治广东省高校重点实验室,广东 广州 510632)
卵圆卡盾藻(Chattonellaovata)是鱼毒性赤潮原因种之一。2001年在我国大鹏湾首次记载了卵圆卡盾藻赤潮[1],此后,日本和墨西哥相继报道了该藻引发的赤潮会导致养殖鱼类大量死亡,海产养殖业损失巨大[2-3]。研究表明,鱼毒性赤潮藻产生的溶血毒素是引起鱼类死亡的重要原因之一,但其具体的致毒机理和毒素的化学结构成分尚不清楚[4]。藻细胞叶绿素三维荧光技术可以准确识别鱼毒性藻类和非鱼毒性藻类并判别鱼毒性藻类毒性的高低[5],提示藻类的光合作用系统可能在鱼毒性藻类产毒中具有重要作用,为此,需进一步探讨藻类的光合色素与藻类溶血活性间的关系。铜和镁离子在藻类叶绿素合成或光合电子传递中具有重要作用,本研究通过调控培养液中上述两种离子的浓度,研究其对卵圆卡盾藻生长、光合色素含量及其溶血活性的影响,进一步探讨溶血毒素与光合色素的关系,旨在为卵圆卡盾藻溶血毒素的产生机制研究提供参考。
1 材料与方法
1.1 实验试剂
1.1.1 溶血毒素提取缓冲液 分别称取0.238 g KCl、8.775 g NaCl、0.551 g CaCl2·2H2O、0.150 g MgSO4和1.476 g TRIS定容至1 dm3高纯水中,用0.1 mol/dm3HCl调pH至7.4,储存于4 ℃条件下备用。
1.1.2 兔血红细胞缓冲液 分别称取0.200 g KCl、8.000 g NaCl、 0.215 g KH2PO4和3.580 g Na2HPO4·12H2O定容至1 dm3高纯水中,用0.1 mol/dm3HCl调pH至7.4,储存于4 ℃条件下备用。
1.1.3 0.5%兔血红细胞溶液 取新西兰兔静脉血在1 000 r/min下离心15 min后弃去上层血清和白细胞,下层红细胞加入适量兔血红细胞缓冲液充分混匀,再次离心弃上清。如此重复3次,最后用兔血红细胞缓冲液配制成体积分数为0.5%的兔血红细胞溶液,储存于4 ℃条件下备用。
1.2 实验方法
1.2.1 藻种培养和实验浓度确定 实验用卵圆卡盾藻来自暨南大学赤潮与海洋生物学研究中心藻种室,分离自大亚湾。卵圆卡盾藻的培养采用人工海水[6],用f/2培养基改良配方配制培养液,接种对数期的藻液后,置于温度为25 ℃,光照强度为3 000 lx,光暗循环 L∶D=12 h∶12 h的光照培养箱中进行培养。
预实验发现,卵圆卡盾藻在经人工海水培养驯化2代之后,当铜和镁金属离子浓度为0.5倍f/2培养基(f/2培养基的铜离子浓度为9.80 μg/dm3,不添加镁离子;镁离子浓度为18.79 g/dm3,不添加铜离子)时即铜离子浓度为4.90 μg/dm3、镁离子浓度为9.40 g/dm3时,只能维持生存而不出现增长现象,在不影响卵圆卡盾藻生长的基础上,探讨光合色素与溶血活性的关系。实验设计金属离子浓度如下:铜离子分别为3.92、5.88、7.84、9.80 μg/dm3,镁离子分别为7.52、11.27、15.03、18.79 g/dm3,即0.4、0.6、0.8、1.0倍f/2培养基。
1.2.2 色素的提取与高效液相色谱检测 色素提取参考Zapata等(1991)和Buffan-Dubau等(2000)的方法[7-8]。取50 cm3藻液进行抽滤,将滤膜剪碎放入离心管中,加入3 cm3体积分数为95%的甲醇在冰浴条件下超声破碎8 min后3 000 r/min离心5 min,取离心后的上清液用针头过滤器过滤2 cm3保存于-80 ℃条件下备用。
采用高效液相色谱法(High Performance Liquid Chromatography, HPLC)对色素进行分析,色谱柱为C8(150.0 nm×4.6 nm, 3.5 μm粒径),柱温为25 ℃,流速为1 cm3/min,洗脱时间为45 min,DAD紫外检测器的检测波长为430、440、470 nm。流动相A为超纯水,B为乙腈,C为甲醇-乙腈-丙酮(体积比为1∶1∶3)混合液,D为甲醇、乙腈-醋酸吡啶混合液。HPLC分析的进样量为100 mm3,为避免色素峰的变形,将200 mm3的提取液和67 mm3的超纯水混合后立即进样,以防色素损失。每次分析时,用30 mm3混合色素标准液、90 mm3甲醇和40 mm3超纯水混匀,作为标准工作液。以上所有的操作均在弱光环境下进行,以免光照破坏色素。
1.2.3 溶血毒素的提取和溶血活性的检测 溶血毒素的提取参照Eschbach等(2001)的方法[9]。取10 cm3藻液在11 000 r/min、4 ℃下离心15 min,弃上清液收集藻细胞,用1 cm3藻细胞毒素提取缓冲液重悬浮,在功率为30%的冰浴条件下超声破碎1 min,即得溶血毒素提取液。取0.5%兔血红细胞溶液200 cm3加入毒素提取液200 mm3。在25 ℃下反应2 h。离心取上清液至96孔板,于414 nm波长下检测其吸光值。溶血活性大小按下列公式计算:
(1)
式(1)中:H为溶血百分数;As为相同体积的溶血毒素提取液与兔血红细胞反应后上清液的吸光值;Aa为阴性对照1,为同体积的提取溶血毒素的缓冲液与红细胞置于相同条件的上清液吸光值;Ab为阴性对照2,为同体积的溶血毒素提取液与红细胞缓冲液置于相同条件的上清液吸光值;Ac为阳性对照,为全溶红细胞上清液的吸光值。
1.2.4 统计分析 实验中所有数据均为3个平行样的平均值±标准差,组间差异通过SPSS 20.0软件的单因素ANOVA统计检验。
2 结果与讨论
2.1 铜和镁离子对卵圆卡盾藻生长的影响
从图1(a)可以看出,卵圆卡盾藻在铜离子浓度为3.92~9.80 μg/dm3范围内,在培养的前8天,铜离子浓度对藻细胞密度无显著影响。第8天后,铜离子浓度为3.92 μg/dm3时,卵圆卡盾藻到达最大藻密度(2.19×104个/cm3)后迅速进入衰退期,且藻细胞密度低于9.80 μg/dm3组(2.87×104个/cm3)。但处于藻类指数生长期时的各组生长速率并无明显差异。铜离子的添加对卵圆卡盾藻的生长至关重要,低浓度的铜离子会使卵圆卡盾藻提前进入衰亡期。周海东等(2016)证明铜的缺失会干扰色素和脂质的合成,铜有助于稳定光系统PSII复合物附近的脂质环境,是光合电子传递链中电子传递体质蓝素的重要元素之一[10]。但高浓度的铜也会抑制光合作用,Perales等(2007)的研究发现栅藻(Scenedesmusincrassatulus)对高浓度的铜离子更为敏感[11]。
如图1(b)所示,在镁离子浓度为7.52~18.79 g/dm3范围内,随着镁离子浓度的增加,卵圆卡盾藻的细胞密度也逐渐增加。镁离子浓度为7.52 g/dm3时,卵圆卡盾藻的细胞密度最低,且在培养的第10天进入衰亡期。Urek等(2019)研究表明钝顶节旋藻(Arthrospiraplatensis)在镁离子不足的条件下生物量降低[12],说明低浓度的镁离子不利于藻类的生长。Hoyoung等(2019)研究不同浓度的镁离子在低光条件下对铜绿微囊藻(Microcystisaeruginosa)的生长影响表明,一定浓度的镁离子促进了铜绿微囊藻的细胞分裂[13],本研究也发现高浓度组镁离子的最大细胞密度明显高于其他组。

图1 不同浓度的铜和镁离子对卵圆卡盾藻生长的影响
2.2 铜和镁离子对卵圆卡盾藻光合色素含量的影响
高效液相色谱分析显示,卵圆卡盾藻的主要光合色素成分包括Mg-2, 4-二乙烯基脱镁卟啉a5单甲基酯(MgDVP)、叶绿素c2、硅甲藻黄素、紫黄质、岩藻黄素和叶绿素a。
在藻类光合系统中,铜离子主要作为光化学系统PSⅡ上电子供体辅基的超氧化物歧化酶参与电子传递,同时也会参与叶绿体光合膜脂的合成[14]。Rodríguez等(2018)研究发现低浓度的铜离子可诱导藻类的光合作用和类胡萝卜素水平的增加,而当铜离子浓度高于最佳水平时也会干扰各种代谢途径,从而抑制光合作用[15]。在本研究中,铜离子浓度范围为3.92~9.80 μg/dm3(图2),卵圆卡盾藻单位藻细胞的MgDVP和叶绿素c2平均含量随浓度的增加呈现先上升后下降的趋势,高浓度和低浓度组的MgDVP平均含量变化显著(p<0.05),其中当铜离子浓度为7.84 μg/dm3时,MgDVP和叶绿素c2平均含量最高。岩藻黄素和硅甲藻黄素的平均含量总体上随着铜离子的浓度增加呈现减少的趋势,组间差异显著(p<0.05),高浓度的铜离子可能抑制了这两种色素的积累。而随着铜离子浓度的增加,叶绿素a的平均含量变化不显著(p>0.05),表明在铜离子浓度为3.92~9.80 μg/dm3的范围内,卵圆卡盾藻的其他光合色素比叶绿素a对铜离子更为敏感。而郭宏实等(2019)研究发现,杜氏盐藻(Dunaliellasalina)的叶绿素a比类胡萝卜素对铜离子敏感[16],可能不同的藻类对铜离子的激性响应不同。
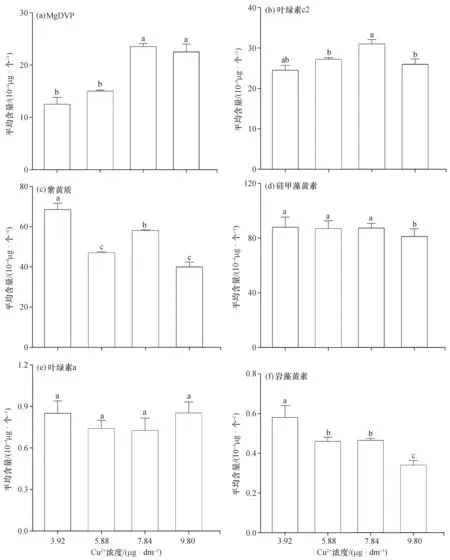
图2 不同浓度的铜离子对卵圆卡盾藻光合色素含量的影响
镁是藻类光合作用的重要元素之一,除了作为多种酶的活化剂参与光合作用之外,镁直接参与叶绿素的合成,是叶绿素中心原子的重要组成,保持天线色素、反应中心及电子载体间的联系,促进光能被高效的吸收、转化和传递[13]。另外,适当的镁也促进类胡萝卜素的合成[14]。在镁离子浓度为7.52~18.79 g/dm3的范围内(图3),卵圆卡盾藻单位藻细胞的MgDVP和叶绿素a的平均含量总体上随着镁离子浓度的增加而减少,高浓度组与低浓度组镁离子平均含量的差异显著(p<0.05),说明高浓度的镁离子抑制了MgDVP和叶绿素a的合成;Urek等也发现钝顶节旋藻(Arthrospiraplatensis)在较高镁离子的条件下叶绿素含量会降低[12]。随着镁离子浓度的增加,叶绿素c2和硅甲藻黄素的平均含量总体上增加,高浓度组与低浓度组相比差异显著(p<0.05),而紫黄质、岩藻黄素平均含量变化不显著(p>0.05)。
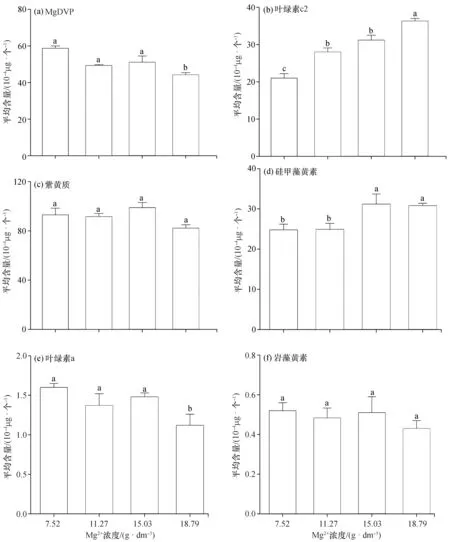
图3 不同浓度的镁离子对卵圆卡盾藻光合色素含量的影响
2.3 铜和镁离子对卵圆卡盾藻溶血活性的影响
溶血毒素是鱼毒性藻类产生的一种可致使动物血红细胞破损的次生代谢产物[17],其产生受多种环境因素的影响。本研究结果显示(图4),铜和镁离子浓度分别为3.92~9.80 μg/dm3和7.52~18.79 g/dm3时,卵圆卡盾藻的溶血活性随着离子浓度增加而增加,铜离子和镁离子在高浓度和低浓度时的溶血活性差异显著(p<0.05)。宋秀凯等(2006)也发现一定浓度的镁离子能够显著增加多变鱼腥藻(Anabaenavariabilis)的溶血活性[18],说明不同金属离子对藻类的溶血活性存在着显著的影响,但其中的机制仍然不清楚。
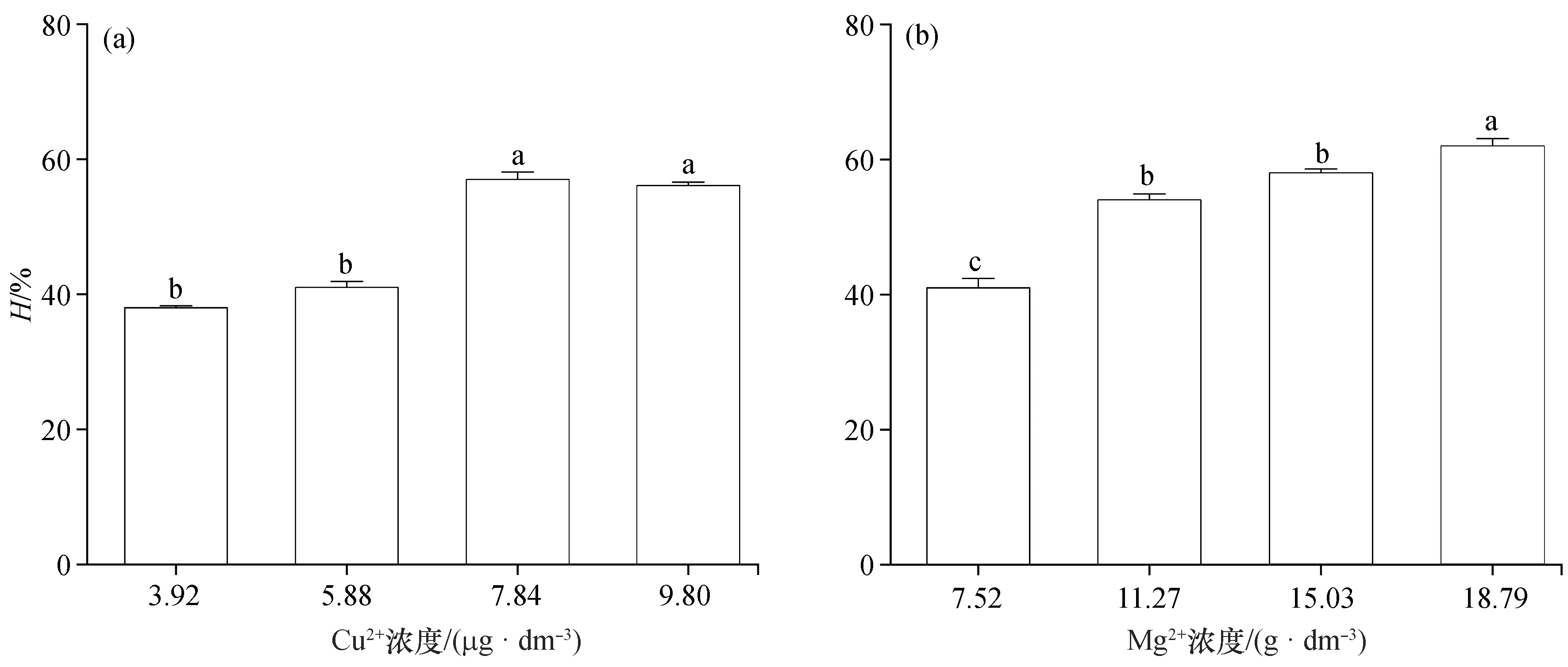
图4 不同浓度的铜和镁离子对卵圆卡盾藻溶血活性的影响
2.4 卵圆卡盾藻的溶血活性与光合色素的关系
为了进一步证明溶血活性与光合色素的关系,对不同浓度铜离子和镁离子条件下的卵圆卡盾藻6种主要色素MgDVP、叶绿素c2、岩藻黄素、紫黄质、硅甲藻黄素和叶绿素a与溶血活性进行因子分析,结果如图5所示。卵圆卡盾藻溶血活性与叶绿素c2在因子1上载荷较高,因子1主要由溶血活性和叶绿素c2决定。因此,相比于其它光合色素卵圆卡盾藻的溶血活性与叶绿素c2关系密切,溶血毒素可能是叶绿素c2的类似物。目前,已知的鱼毒性藻类溶血毒素组分主要有卟啉类、不饱和脂肪酸类、糖脂类及脂肪酸酰胺类类似物。就毒素成分与光合色素而言,Kuroda等(2005)研究表明,纯化的海洋卡盾藻(Chattonellamarina)溶血毒素与叶绿素c在446、583、635 nm处的吸收峰极为相似,可能是叶绿素c的衍生物[19]。Miyazaki等(2005)对褐藻Eiseniabicyclis的提取物分析表明其溶血毒素是含有吡咯环的卟啉衍生物,其结构与脱镁叶绿酸甲酯类似[20],还有学者发现许多叶绿素的衍生物或卟啉类物质都是光活毒素,如Henrikson等(2010)从环状异帽藻(Heterocapsacircularisquama)中提取的叶绿素c2 类似物就具有一定的细胞毒性[21]。赤潮异弯藻(Heterosigmaakashiwo)和海洋卡盾藻的溶血毒素提取物也具有光依赖性[22]。除卟啉衍生物外,由糖脂类和多不饱和脂肪酸类组成的溶血毒素也可能作为光合作用的产物。糖脂是含有糖基的脂质,是藻类光合作用的次级产物,也是植物光合膜脂的主要成分,受光合作用调控。在光合作用(PS II)阶段,可能作为电子的供体或受体存在[23]。有学者证实小定鞭藻(Prymnesiumparvum)溶血毒素为糖脂类化合物,海洋卡盾藻的溶血毒素是糖脂类和多不饱和脂肪酸类;针胞藻(Fibrocapsajaponica)产生的4种多不饱和脂肪酸均具有溶血活性[24]。
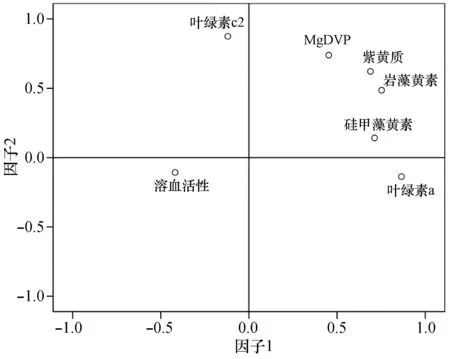
图5 卵圆卡盾藻的溶血活性与光合色素的关系
本研究结果发现,不同浓度的镁离子对卵圆卡盾藻的溶血活性和叶绿素c2含量的影响趋势变化一致, 且组间差异显著,可能镁离子通过参与叶绿素c衍生物的合成来影响溶血活性,卵圆卡盾藻的溶血毒素很可能就是叶绿素c2或其类似物。而不同浓度的铜离子可能是通过调控多不饱和脂肪酸的合成,参与光合膜脂的合成显著影响其溶血活性。对比不同金属离子浓度下的光合色素及溶血毒素的变化可以发现,鱼毒性藻类卵圆卡盾藻的溶血毒素与藻类的光合作用系统直接相关,提示溶血毒素可能就是藻细胞叶绿素及其衍生物或参与光合作用的代谢产物。
3 结论
(1)铜和镁离子均会对卵圆卡盾藻的生长、光合色素以及溶血毒素产生影响。随着铜和镁离子浓度的增加,卵圆卡盾藻的细胞密度增加,低浓度铜和镁离子对藻类生长有一定限制作用。高浓度的铜离子可能抑制卵圆卡盾藻的岩藻黄质和硅甲藻黄素的积累,镁离子浓度增加,叶绿素c2和硅甲藻黄素会随之增加。高浓度的铜离子及镁离子也会使卵圆卡盾藻的溶血活性增加,说明过量或不足的金属离子均可能影响藻类的生长、光合色素以及溶血毒素的合成。
(2)不同浓度的镁离子对卵圆卡盾藻的溶血活性和叶绿素c2含量的影响趋势变化一致;卵圆卡盾藻的叶绿素与溶血活性因子分析进一步提示其溶血活性和叶绿素c2关系密切,溶血毒素可能是叶绿素c2或其类似物。
