茶树种质资源花器官微形态特征观察
2021-08-02樊晓静于文涛蔡春平王泽涵林浥张琛叶乃兴
樊晓静 于文涛 蔡春平 王泽涵 林浥 张琛 叶乃兴


摘要:【目的】对不同茶树种质资源花器官的微形态特征进行观察与分析,为种质资源鉴定评价提供参考依据。【方法】利用真空冷冻干燥和冷场发射扫描电镜技术,对11份茶树花器官的花柄、花托、萼片、花瓣、子房、花柱、柱头和花丝的表皮纹饰、气孔和茸毛纹饰等微形态特征进行系统观察,并进行变异系数和主坐标分析。【结果】茶树花柄和花托的表皮纹饰较相似,为细长条纹形,且在部分种质的花柄和花托上发现茸毛和气孔;萼片的内表皮光滑,可分为表皮细胞凹凸不平、凹陷和饱满3种类型,在其表面具平滑型茸毛;萼片外表皮光滑具条纹纹饰,在其表面分布着无规则气孔器且不同茶树种质气孔器特征不同;花瓣表皮细胞形状主要为不规则形、五边形、六边形和近圆形,其表皮分布着波状、条纹状、辐射状等纹饰;花丝表皮细胞为不规则多边形,排列紧密,具波状、丝状、条状的表皮纹饰,气孔主要分布在花丝的中下部;花柱表皮细胞排列整齊,其细胞形状可分为梭形、长条纹形和多边形3种类型;子房壁表皮细胞凹凸程度不同,满被平滑型茸毛。对参试茶树种质花器官的全部气孔数量性状进行测量统计,花柄气孔器大小为142.99~431.66 μm2,气孔开度为0.19~0.92;花托气孔器大小为201.48~642.17 μm2,气孔开度为0.26~0.62;萼片气孔器大小为219.74~563.32 μm2,气孔开度为0.37~0.52;花瓣气孔器大小为401.80~1322.07 μm2,气孔开度为0.38~0.66;花丝气孔器大小为257.90~706.74,气孔开度为0.32~0.73。对茶树花器官气孔数量性状变异分析,其变异系数平均值为18.5%;以40个微形态性状指标和16个微形态质量性状指标对11份种质进行主坐标分析,结果发现仅用质量性状时可有效区分种质。变异分析和主坐标分析表明茶树花器官的气孔相关数量性状种质内变异较大,而表面纹饰等质量性状具有较强的遗传稳定性。【结论】在茶树分类鉴定中,可适当考虑花器官的质量性状,并优先选择花器官的萼片、子房壁等纹饰特征作为识别不同茶树品种的依据。
关键词: 茶树;种质资源;花器官;微形态;扫描电镜
中图分类号: S571.1 文献标志码: A 文章编号:2095-1191(2021)03-0700-11
Floral micromorphological trait of tea germplasm resources
FAN Xiao-jing1, YU Wen-tao2*, CAI Chun-ping2, WANG Ze-han1, LIN Yi1,
ZHANG Chen1, YE Nai-xing1*
(1College of Horticulture, Fujian Agriculture and Forestry University/Key Laboratory of Tea Science in Universities of Fujian Province, Fuzhou 350002, China; 2 Technology Centre of Fuzhou Customs/Fujian Key Laboratory for
Technology Research of Inspection and Quarantine, Fuzhou 350001, China)
Abstract:【Objective】The microform of different tea floral structures was observed and analyzed,which would provide reference for the identification and utilization of tea tree germplasm. 【Method】The vacuum freeze drying and field emission scanning electron microscopy(FE-SEM) observations were applied to investigate floral micromorphological traits of 11 tea, Camellia sinensis(L.) O. Kuntze, germplasms, and the coefficient of variation and principal coordinates were analyzed. The epidermal micromorphological traits(epidermises ornamentation, stomata and trichome ornamentation) of tea floral organs, such as peduncles, receptacles, sepal, petal, ovary, style, stigma and filament, were reported and analyzed. 【Result】The peduncle and receptacle epidermises were similar in stripe shape. And trichomes and stomata were found in some germplasms. The inner epidermises of tea sepals were smooth, and could be divided into three types: uneven, concave and plump epidermal cells. There were smooth trichomes on the inner epidermises of the sepals. The ou-ter epidermises of sepals were smooth and striated. There were irregular stomata on the surface, and different tea germplasms had different stomatal characteristics. The shapes of petal epidermal cells could be divided into four types: irregular, pentagon, hexagon and nearly round. On the petal epidermises, the wavy, striate and radial ornamentation were distributed. The filament epidermal cells were irregular polygons with stomates. The epidermises were closely arranged with wavy, filiform and striped epidermal ornamentation. Stomata were distributed in the middle and lower segments of filament. The epidermal cells of styles were neatly arranged, and the cell shapes could be divided into three types: fusiform, elongated and polygonal. The epidermal cells of ovary were different in the degree of smoothness, and were covered with smooth type trichomes. The stomatal quantitative characters in flower organs of tea germplasm were measured, the peduncle stomach size were 142.99-431.66 μm2, and the stomatal aperture were 0.19-0.92. The receptacles stomach size were 201.48-642.17 μm2, and the stomatal aperture were 0.26-0.62. The sepal stomach size were 219.74-563.32 μm2, and the stomatal aperture were 0.37-0.52. The petal stomach size were 401.80-1322.07 μm2, and the stomatal aperture were 0.38-0.66. The filament stomach size were 257.90-706.74 μm2, and the stomatal aperture were 0.32-0.73. The variations of all stomatal quantitative characters were analyzed, and the average coefficient of variation was 18.5%. The principal coordinate analysis of 11 germplasms were carried out with all 40 micromorphological traits and 16 qualitative micromorphological traits, the results showed that only the quality traits could be used to distinguish the germplasms. It indicated that the quantitative stomatal traits had large variations intro-germplasm, while qualitative traits had strong genetic stability. 【Conclusion】In the classification and identification of tea plants, the quality characters of flower organs can be properly consi-dered, and the ornamentation characteristics of sepals and ovary walls of flower organs can be preferentially selected as the basis for identifying different tea varieties.
Key words: Camellia sinensis(L.) O. Kuntze; germplasm resources; floral organ; micromorphology; scanning electron microscope(SEM)
Foundation item: Fujian Province “2011 Collaborative Innovation Center” Chinese Oolong Tea Industry Innovation Center(Cultivation)Special Project(Minkejiao〔2015〕75);Fujian Agriculture and Forestry University Construction Pro-ject for Technological Innovation and Service System of Tea Industry Chain(2020-01);Special Fund for Science and Technology Innovation of Fujian Zhang Tianfu Tea Development Foundation(FJZTF01);Scientific Research Project of Gene-ral Administration of Customs of China(2020HK187)
0 引言
【研究意义】茶树[Camellia sinensis(L.) O. Kuntze]属于山茶科(Theaceae)山茶属(Camellia)植物,是我国广泛栽培的主要经济作物之一。植物的开花及结果过程有助于保持植物的遗传多样性(Akagi et al.,2018)。花作为植物的繁殖器官,受环境影响较小,具有较强的遗传稳定性(Caseys,2018)。花器官外部形态特征是研究茶树种质资源演化的主要依据,也是茶树品种分类的主要依据(叶乃兴等,2005;陈常颂和余文权,2016;Konarska,2017),而显微形态结构是茶树外部形态结构的微观体现,因此对茶树花器官的微形态结构的研究尤为重要。【前人研究进展】目前常采用扫描电镜对植物微形态进行观察。扫描电镜在揭示不同植物物种及品种间的微形态差异方面应用越来越广泛,成为探讨植物系统发育关系的重要手段(钟秋生等,2012;Yu et al.,2012;Fortini et al.,2013;石力匀等,2020)。国内外已有较多学者对植物花器官微形态进行了报道。曹清河等(2010)对10种花粉形态进行微形态观察,结果发现其外壁纹饰类型间存在差异,各自具有特异性。莫爱琼等(2015)对花器官的微形态特征观察,结果显示柱头和花柱的表面纹饰具明显差异,可为品种鉴定提供参考。张永芳等(2016)利用扫描电镜对柿属植物花粉形态进行观察和分析,结果表明柿属种间的花粉形态存在异同,可为柿属分类学提供依据。Millner和Baldwin(2016)研究花萼及愈伤组织等微形态,认为Restrepia代表一个特殊的兰花属,且这种传粉机制可能与该属的繁殖系统直接相关。de Costa等(2017)研究表明花瓣圆锥细胞的2个微形态特征对于物种分组具有较大影响,且圆锥形细胞顶点之间的距离是与传粉媒介相互作用的重要参数。宋科等(2018)通过扫描电镜对4个蓝莓品种花器官的超微结构进行比较,结果表明蓝莓品种间花丝表皮毛、柱头表面形态等花部结构存在较大差异,可作为品种识别的依据。王非等(2018)通过扫描电镜对褐毛铁线莲的花部结构进行观察,发现花丝被毛,花药无毛,且雄蕊和雌蕊的相间分布更有利于铁线莲的授粉。张雪娟等(2019)对沙棘花形态结构的研究结果表明,花表皮的多样性在抵御寒冬、保护生殖器官方面起到重要作用。方仁等(2020)对8个番茄品种的花粉形态观察结果显示,花粉形状、大小等性状差异可为番茄的亲缘关系和分类鉴定提供依据。王茜等(2020)对凤仙花组植物的花粉進行扫描电镜观察,结果表明花粉的网眼及网脊等微形态特征可为凤仙花组组下划分及种水平的界定提供依据。【本研究切入点】目前,关于茶树花器官微形态,仅见花粉微形态有相对较多的描述和研究(彭艾,2009;魏兆兆等,2012;谢微微等,2018;樊晓静等,2019),未见关于茶树花柄、花托、花萼、子房、花柱、柱头和花丝等微形态观察的研究报道。【拟解决的关键问题】以11份茶树种质资源为研究对象,利用扫描电镜技术对茶树的花柄、花托、萼片、花瓣、子房、花柱、柱头及花丝等结构进行系统的微形态观察与分析,全面、清晰了解茶树花器官微形态特征,探讨茶树种质资源微形态特征,以期为茶树种质资源的鉴定评价工作提供理论及试验依据。
1 材料与方法
1. 1 试验材料
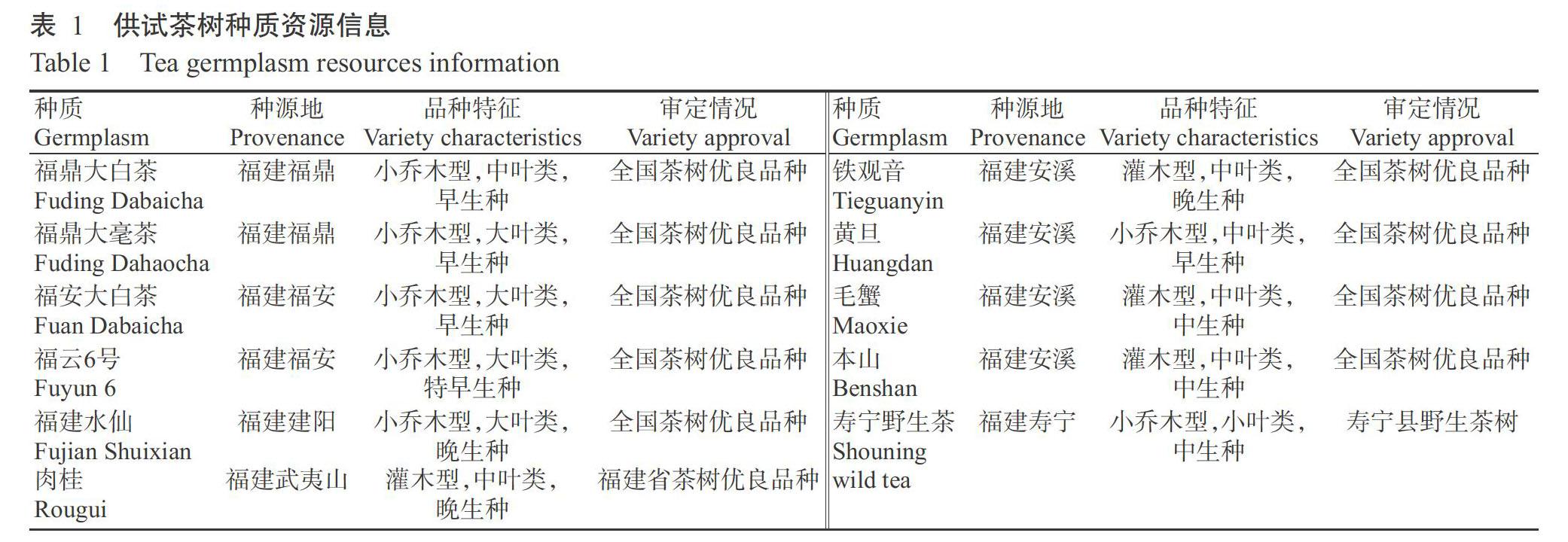
于2018和2019年的11—12月采摘茶树盛花期花朵,取样地点为福建农林大学茶树品种资源园。供试材料为福鼎大白茶等11份茶树种质(表1),每份种质资源分别从3棵茶树上取样,其中寿宁野生茶的花取自单株上。本研究所取茶树花器官分别为花柄、花托、萼片、花瓣、子房、花柱、柱头和花丝,如图1所示。
1. 2 试验方法
解剖茶树花器官,在体视显微镜下观察拍照。扫描电镜前处理方法(杨国一等,2018a)如下:将解剖新鲜茶树花器官于2.5%戊二醛溶液中固定,置于4 ℃冰箱中5 h;然后依次用50%、70%和90%浓度的乙醇进行梯度脱水,每次15 min,再用100%乙醇脱水3~5次,每次15 min;最后将花器官材料置于100%叔丁醇中浸泡15 min后,放入真空冷冻干燥机进行干燥。用双面导电胶将茶树花器官固定于圆柱形样品台,经E-1010型离子溅射镀膜仪(日本日立公司)喷金80 s,在SU-8010型冷场发射扫描电镜(日本日立公司)下观察茶树花器官的表皮细胞、纹饰、气孔器及表皮毛特征,同时分别在400~8000的放大倍率下观察拍照。
用Image J(美国National Institutes of Health)对花器官的内气孔长、内气孔宽、外气孔长和外气孔宽进行数据测量,各性状测量均进行15次重复。计算气孔器大小(外气孔长×外气孔宽)和气孔开度(内气孔宽/内气孔长)。性状量化方法为定量性状直接取其数值,定性性状则按不同形态进行编号。
1. 3 统计分析
采用Excel 2010对调查数据进行计算与汇总,利用MVSP v3.13n(英国Kovach Computing Services公司)进行主坐标分析和聚类分析。
2 结果与分析
2. 1 茶树花柄、花托微形态性状
通过扫描电镜观察,11份茶树种质资源的花柄和花托表皮纹饰颇为相似(图2-A1和图2-A2),细胞形状皆为矩形,表皮纹饰为规则的细长条纹形,表面具丝状体附着。福云6号、福鼎大毫茶和肉桂的花柄表皮上有少量气孔存在,其气孔器大小为142.99~431.66 μm2,气孔开度为0.19~0.92,其他种质花柄未见气孔;福云6号、肉桂、毛蟹和本山花柄表皮上具少量茸毛(图2-A3),且茸毛纹饰均为平滑型。供试茶树种质花托表皮均具气孔(图2-A4),气孔器大小为201.48~642.17 μm2,气孔开度为0.26~0.62(表2)。
2. 2 茶树萼片、花瓣微形态性状
由扫描电镜观察结果(表2和表3)可知,各茶树种质资源的萼片内表皮细胞为近长方形或多边形,排列紧凑,表面较光滑,且在其中部或边缘具平滑型茸毛(图2-B1);萼片外表皮细胞为不规则状,表面光滑具条纹纹饰,并在中间部位分布着无规则气孔器,气孔类型皆为凸起型。其中,福鼎大白茶、福云6号和福鼎大毫茶的花萼内表皮细胞凹凸不平(图2-B2);福安大白茶和肉桂花萼内表皮细胞稍平展但细胞凹陷较多(图2-B3);铁观音、毛蟹、本山和寿宁野生茶的花萼内表皮细胞平展,均较饱满(图2-B4)。每份茶树种质的萼片外表皮的气孔器特征有所不同,福鼎大白茶的花萼气孔器明显凸出于表皮(图2-B5),其他种质均稍突出于表皮(图2-B6);供试茶树种质气孔的外拱盖均较平滑;福鼎大白茶、福云6号、福鼎大毫茶和寿宁野生茶的花萼外缘角质层呈脊状增宽;除本山的花萼外缘角质层呈辐射状纹饰环绕(图2-B7),其他种质的外缘角质层呈条状纹饰环绕(图2-B8)。各品种茶树的萼片气孔器大小为219.74~563.32 μm2,气孔开度为0.37~0.52(表2)。
由图2-C1~图2-C4及表4可知,茶树的花瓣表皮细胞形状主要呈不规则形、五边形、六边形和近圆形,其中分布着波状、条纹状、辐射状等纹饰。福鼎大白茶、福鼎大毫茶、肉桂、铁观音和本山的花瓣表皮微形态相对较多样,其他种质的花瓣表皮微形态则相对较简单。除福云6号外,其他种质的花瓣表皮均有气孔且气孔呈近圆形;福鼎大白茶气孔器稍内陷于表皮(图2-C5),其他种质资源与表皮持平。铁观音的气孔外拱盖被明显条状(图2-C6),而其他种质资源均较平滑。福安大白茶和铁观音气孔器外缘为辐射状纹饰(图2-C7),其他种质为条状纹饰(图2-C8)。各品种茶树的花瓣气孔器大小为401.80~1322.07 μm2,气孔开度为0.38~0.66。
2. 3 茶树雄蕊微形态性状
通过扫描电镜观察,茶树花丝表皮细胞排列紧密,纹饰为平行于花丝的波状、丝状、条状。气孔主要集中在花丝的中下部(图2-D1),气孔类型为近圆形,与花瓣相似,未发现茸毛。其中,福鼎大白茶、福云6号和福安大白茶的表皮纹饰主要为紧密的波状(图2-D2);肉桂、黄旦和铁观音纹饰主要为卷曲的丝状(图2-D3);福鼎大毫茶、福建水仙、毛蟹、本山和寿宁野生茶主要为稍曲的条状(图2-D4)。福云6号的花丝气孔器较大(706.74 μm2),铁观音相对最小(257.90 μm2);福云6号和本山的花丝气孔开度相对较大(0.73),寿宁野生茶和福安大白茶则相对较小(0.32~0.38)(表2)。
2. 4 茶树雌蕊微形态性状
通过扫描电镜观察,茶树花柱表皮具乳突状细胞,并有花粉粒附着,表明授粉过程正在柱头上进行。柱头上均具分泌液,属于湿柱头。花柱整体呈长圆柱形(图2-E1),顶端渐细,表皮细胞排列整齐。花柱细胞形状可分为3种类型,福鼎大白茶、福鼎大毫茶、肉桂、福建水仙和本山为梭形(图2-E2),福云6号、福安大白茶、铁观音和寿宁野生茶为长条纹形(图2-E3),黄旦和毛蟹则为多边形(图2-E4)。茶树子房壁表皮细胞呈不规则多边形,满被茸毛(图2-F1),茸毛纹饰为平滑型。福鼎大白茶、黃旦、铁观音、毛蟹表皮有似圆形细胞凸起(图2-F2),纹饰光滑,凸起程度从大到小顺序为福鼎大白茶>铁观音>毛蟹>黄旦。福鼎大毫茶、肉桂、本山和寿宁野生茶子房表皮细胞内陷(图2-F3),福云6号和福安大白茶的表皮纹饰则较平展(图2-F4)。
2. 5 茶树花器官气孔相关数量性状的变异分析
对参试茶树种质花器官全部气孔相关数量性状进行变异分析,其变异系数为0.4%~94.6%,平均为18.5%。种质内气孔相关数量性状的变异系数平均值为8.18%~27.61%,均值为17.99%;种质间气孔相关数量性状的变异系数为12.22%~23.92%,均值为18.07%。可见,茶树花器官气孔相关数量性状在种质内和种质间的变异系数均较大。
2. 6 基于茶树微形态性状的主坐标分析
对包括气孔相关数量性状在内的40个茶树花器官微形态性状进行主坐标分析,结果(图3)显示,参试种质材料中,福云6号、福安大白茶、黄旦、铁观音、毛蟹和本山可明显与其他种质资源区分开,而福鼎大毫茶、肉桂、寿宁野生茶、福鼎大白茶和福建水仙等种质间存在交叉和重叠现象,尤其是福鼎大白茶和福建水仙在主坐标图中分布距离较远。对除气孔相关数量性状外的16个茶树花器官微形态质量性状进行主坐标分析,结果(图4)显示,11份种质均能各自聚在一起,且与其他种质明显区分开。主坐标分析结果(表5)显示,前5个主成分的累积贡献率达83.886%,表明其可代表所选用性状的绝大部分信息。第1主成分的贡献率为28.491%,主要由萼片气孔外缘角质层和子房壁纹饰决定;第2主成分的贡献率为19.592%,主要由花瓣纹饰形状和萼片细包凹凸决定;第3主成分的贡献率为16.072%,主要由花瓣表皮是否平展和花丝纹饰决定;第4主成分的贡献率为9.913%,主要由萼片外表皮纹饰和花丝纹饰决定;第5主成分的贡献率为9.817%,主要由萼片的外表皮纹饰及其气孔外缘环绕类型、花瓣气孔外拱盖被披纹饰类型决定。对40个花器官微形态性状和16个微形态质量性状(未含24个气孔相关数量性状)的综合分析表明,当包括气孔器数量性状时,未能明显区分各种质,存在交叉及重叠现象;而仅包含质量性状时,可有效区分各种质。
2. 7 基于茶树微形态性状的聚类分析
基于11份供试材料,对40个微形态性状进行UPGMA法聚类分析,结果(图5)表明,在欧式距离为6.3时,聚类可分为2支,福鼎大白茶与福云6号聚为一支,这2份种质的微形态特点是萼片内表皮细胞凹凸不平,茸毛纹饰为平滑型,气孔外缘角质层有脊状增宽且外缘有条状纹饰环绕,同时花瓣表皮细胞形状为不规则形,纹饰类型为波状。福鼎大毫茶与福安大白茶等9份种质聚为另一支,这些种质的共同特点是萼片内表皮细胞形状为长方形、多边形,茸毛纹饰为平滑型,花瓣气孔器与表皮持平。
3 讨论
植物的表皮纹饰特征具有重要的分类学价值(Ullah et al.,2018)。本研究中电镜观察结果表明,茶树花器官的相同部位微形态性状表现稳定,不同部位微形态性状呈现差异性。如花柱纹饰类型为长条纹形、梭形和多边形3类,子房壁表皮纹饰光滑,细胞呈凸起、平展或凹陷3种纹饰。本研究观察发现茶树花器官的萼片上有茸毛,花丝的表皮上无茸毛,与李远志(1986)对花器官的观察结果一致。表皮毛的纹饰特征是研究植物分类学的重要手段(王虹等,2014)。本研究观察发现花柄、花托、萼片和子房的茸毛纹饰均为平滑型,说明茶树花器官的茸毛纹饰特征较统一。
气孔存在于植物的茎、叶和花等器官表面,控制着水分流失和气体交换。在茶树中,关于叶片气孔的研究较多(杨国一等,2018b)。本研究报道了茶树花柄、花托、花瓣、萼片和花丝的表皮也具气孔。除此之外,在观察中还发现茶树萼片上气孔分布较多,花柄、花托和花丝气孔较少,花瓣则居于中间。植物气孔性状是环境和基因共同影响的结果,气孔器类型具有一定的遗传稳定性,而气孔大小受环境影响很大(严学成,1990;李密密等,2012)。本研究观察发现,茶树花柄、花托和萼片气孔器类型均为长卵形,花瓣和花丝的气孔器类型均为近圆形。与杨国一等(2018b)研究结果相似,本研究对茶树花器官气孔的变异系数分析也发现,气孔微形态性状在种质内和种质间皆具较高的变异系数。
对于形态学特征分析,往往有很多有一定相关性的变量,因此进行多元分析就相对复杂。主坐标分析是一种非约束性的数据降维分析方法,可将多个变量化为少数几个指标,从而更好地描述材料构成特征(赵梦然等,2012;王海平等,2014)。本研究分别对所有40个花器官微形态性状和16个微形态质量性状(未含24个气孔相关数量性状)进行主坐标分析,结果表明花器官气孔器数量性状不具有遗传稳定性,不能作为茶树花器官微形态分类鉴定的依据,而茶树花器官的萼片表皮纹饰、子房壁纹饰、花瓣表皮纹饰、花丝纹饰等微形态质量性状作为稳定的性状,在茶树品种鉴定及分类等研究中有较高的价值。
4 结论
11份茶树花器官气孔数量性状变异较大,花器官质量性状较种质鉴定的稳定。因此,在茶树分类鉴定中,可适当考虑花器官的质量性状,并可优先选择花器官的萼片、子房壁等纹饰特征作为识别不同茶树品种的依据。
参考文献:
曹清河,张安,李强,李秀英,王欣,王路,马代夫. 2010. 甘薯属10种植物花粉形态扫描电镜观察[J]. 西北植物学报,30(3): 530-534. [Cao Q H,Zhang A,Li Q,Li X Y,Wang X,Wang L,Ma D F. 2010. Pollen morphology of 10 species of Ipomoea by scanning electron microscope(SEM)[J]. Acta Botanica Boreali-Occidentalia Sinica,30(3): 530-534.]
陈常颂,余文权. 2016. 福建省茶树品种图志[M]. 北京:中国农业科学技术出版社. [Chen C S,Yu W Q. 2016. Records of tea varieties in Fujian Province[M]. Beijing: China Agricultural Science and Technology Press.]
樊晓静,于文涛,刘登勇,卢明基,郑洁,陈晓岚,魏明秀,林浥,叶乃兴. 2019. 福建寿宁野生茶树种质资源叶片和花粉微形态观察[J]. 福建农业学报,34(3): 298-305. doi:10.19303/j.issn.1008-0384.2019.03.007. [Fan X J,Yu W T,Liu D Y,Lu M J,Zheng J,Chen X L,Wei M X,Lin Y,Ye N X. 2019. Micromorphology of leaves and pollens of wild tea(Camellia sinensis) germplasms from Shou-ning,Fujian[J]. Fujian Journal of Agricultural Sciences,34(3): 298-305.]
方仁,安振宇,黄伟雄,白先进,尧金燕,龙兴,周双云,张继. 2020. 8个番荔枝栽培品种的花粉形态扫描电镜观察[J]. 南方农业学报,51(7): 1553-1559. doi:10.3969/j.issn. 2095-1191.2020.07.006. [Fang R,An Z Y,Huang W X,Bai X J,Yao J Y,Long X,Zhou S Y,Zhang J. 2020. Morphology of pollens of eight Annona squamosa L. varieties by scanning electron microscope[J]. Journal of Southern Agriculture,51(7): 1553-1559.]
李密密,舒璞,周義锋,郭建林,孙小芹,杭悦宇. 2012. 中国薯蓣属植物叶表皮微形态特征及其分类学意义[J]. 西北植物学报,32(11): 2232-2242. doi:10.3969/j.issn.1000-4025.2012.11.014. [Li M M,Shu P,Zhou Y F,Guo J L,Sun X Q,Hang Y Y. 2012. Micro-morphological characteristics of leaf epidermis and its taxonomic significance in Dioscorea from China[J]. Acta Botanica Boreali-Occidentalia Sinica,32(11): 2232-2242.]
李远志. 1986. 国内外几个茶树大叶品种的花器官形态分析[J]. 福建茶叶,(4): 15-22. [Li Y Z. 1986. Morphological analysis of flower organs of several large leaf tea cultivars at home and abroad[J]. Tea in Fujian,(4): 15-22.]
莫愛琼,胡晓颖,高丽霞. 2015. 两个蓝莓品种花器官的形态特征观察[J]. 热带亚热带植物学报,23(3): 295-300. doi:10.11926/j.issn.1005-3395.2015.03.010. [Mo A Q,Hu X Y,Gao L X. 2015. Morphological characteristics of flowers of two cultivars of blueberry[J]. Journal of Tropical and Subtropical Botany,23(3): 295-300.]
彭艾. 2009. 闽台主要茶树品种遗传多样性及亲缘关系研究[D]. 福州:福建农林大学. doi:10.7666/d.y1515091. [Peng A. 2009. Studies on the genetic diversity and phylogentic relationship of main tea plant cultivars from Fujian and Taiwan Provinces[D]. Fuzhou: Fujian Agriculture and Fores-try University.]
石力匀,王政,何松林,李玲达,王胜楠,孙畅,杨博. 2020. 切花月季弯梗现象的形态解剖学观察[J]. 河南农业科学,49(9): 120-128. doi:10.15933/j.cnki.1004-3268.2020.09. 015. [Shi L Y,Wang Z,He S L,Li L D,Wang S N,Sun C,Yang B. 2020. Studies on morphological characteristics and anatomical structure of bent peduncle phenomenon in rosa hybrida[J]. Journal of Henan Agricultural Sciences,49(9): 120-128.]
宋科,杨静慧,夏凯丽,刘艳军,梁发辉,张卫华. 2018. 不同品种蓝莓花器官超微结构差异[J]. 天津农学院学报,25(2): 20-23. doi:10.19640/j.cnki.jtau.2018.02.006. [Song K,Yang J H,Xia K L,Liu Y J,Liang F H,Zhang W H. 2018. Differences in ultrastructure of floral organs between different blueberry varieties[J]. Journal of Tianjin Agricultural University,25(2): 20-23.]
王非,郭艳平,王竞红,张永胜,李强,郑瑞. 2018. 褐毛铁线莲花部结构的扫描电镜观察[J]. 江苏农业科学,46(9): 140-144. doi:10.15889/j.issn.1002-1302.2018.09.032. [Wang F,Guo Y P,Wang J H,Zhang Y S,Li Q,Zheng R. 2018. Scanning electron microscopic observation on flower structure of Clematis fusca[J]. Jiangsu Agricultural Sciences,46(9): 140-144.]
王海平,李锡香,沈镝,邱杨,宋江萍,张晓辉,Philipp W.Simon. 2014. 基于表型性状的中国大蒜资源遗传多样性分析[J]. 植物遗传资源学报,15(1): 24-31. doi:10.13430/ j.cnki.jpgr.2014.01.004. [Wang H P,Li X X,Shen D,Qiu Y,Song J P,Zhang X H,Simon P W. 2014. Diversity evaluation of garlic (Allium sativum L.) clones from China based on morphological characteristics[J]. Journal of Plant Genetic Resources,15(1): 24-31.]
王虹,王磊,范林仙,刘翛然,王咏星. 2014. 新疆14种青兰属植物叶表皮微形态结构研究[J]. 西北植物学报,34(10): 2004-2019. doi:10.7606/j.issn.1000-4025.2014.10.2004. [Wang H,Wang L,Fan L X,Liu X R,Wang Y X. 2014. Leaf epidermal micro-morphology structures of fourteen Dracocephalum L. species from Xinjiang[J]. Acta Bota-nica Boreali-Occidentalia Sinica. 34(10): 2004-2019.]
王茜,杨旭东,夏常英,李瑾,于胜祥. 2020. 凤仙花组(Impatiens sect. Impatiens)植物花粉形态研究[J]. 园艺学报,47(5): 893-906. doi:10.16420/j.issn.0513-353x.2019-0426. [Wang Q,Yang X D,Xia C Y,Li J,Yu S X. 2020. Pollen morphology of Impatiens sect. Impatiens(Balsaminaceae)[J]. Acta Horticulturae Sinica,47(5): 893-906.]
魏兆兆,谢云,孟辉,郑玉梅,吴窈窈. 2012. 3种类型浙江红山茶的花粉形态学研究[J]. 浙江农林大学学报,29(4): 634-638. doi:10.3969/j.issn.2095-0756.2012.04.024. [Wei Z Z,Xie Y,Meng H,Zheng Y M,Wu Y Y. 2012. Pollen morphology for three types of Camellia chekiangoleosa[J]. Journal of Zhejiang A & F University, 29(4): 634-638.]
谢微微,于文涛,杨国一,陈静,潘玉华,叶乃兴. 2018. 14个茶树品种的花粉微形态观察[J]. 南方农业学报,49(9): 1698-1704. doi:10.3969/j.issn.2095-1191.2018.09.03. [Xie W W,Yu W T,Yang G Y,Chen J,Pan Y H,Ye N X. 2018. Micromorphological observation on pollen of 14 cultivars of tea tree (Camellia sinensis)[J]. Journal of Sou-thern Agriculture,49(9): 1698-1704.]
严学成. 1990. 茶树形态结构与品质鉴定[M]. 北京: 农业出版社. [Yan X C. 1990. Morphological structure and qua-lity identification of tea[M]. Beijing: Agriculture Press.]
杨国一,于文涛,蔡春平,陈笛,谢微微,王鹏杰,叶乃兴. 2018a. 茶树叶片扫描电镜样品制备方法的比较研究[J]. 江苏农业科学,46(3): 95-98. doi:10.15889/j.issn.1002-1302.2018.03.024. [Yang G Y,Yu W T,Cai C P,Chen D,Xie W W,Wang P J,Ye N X. 2018a. Comparative study on the preparation methods of tea leaf SEM samples[J]. Jiangsu Agricultural Sciences,46(3): 95-98.]
杨国一,于文涛,郑晶,陈静,谢微微,叶乃兴. 2018b. 乌龙茶种质叶片微形态特征的扫描电镜观察[J]. 南方农业学报,49(10): 2020-2027. doi:10.3969/j.issn.2095-1191.2018. 10.18. [Yang G Y,Yu W T,Zheng J,Chen J,Xie W W,Ye N X. 2018b. Scanning electron microscopy observation on foliar micromorphology of Oolong tea germplasms[J]. Journal of Southern Agriculture,49(10): 2020-2027.]
叶乃兴,杨江帆,邬龄盛,王振康. 2005. 茶树花主要形态性状和生化成分的多样性分析[J]. 亚热带农业研究,1(4): 32-35. doi:10.13321/j.cnki.subtrop.agric.res.2005.04.008. [Ye N X,Yang J F,Wu L S,Wang K Z. 2005. Diversity analysis of the main morphologyical traits of tea plant flower and its biochemical components[J]. Subtropical Agriculture Research,1(4): 32-35.]
張雪娟,张欣,王小莉,刘庆华,郭海利,张小民. 2019. 中国沙棘花形态结构的扫描电镜观察[J]. 山西农业大学学报(自然科学版),39(3):7-13. doi:10.13842/j.cnki.issn1671-8151.201810047. [Zhang X J,Zhang X,Wang X L,Liu Q H,Guo H L,Zhang X M. 2019. Blossom anatomy of Hippophae rhamnoides subsp. sinensis Rousi[J]. Journal of Shanxi Agricultural University(Natural Science Edition),39(3): 7-13.]
张永芳,胡超琼,杨勇,朱仁胜,郭静,王仁梓. 2016. 柿属8种植物花粉形态观察[J]. 园艺学报,43(6): 1167-1174. doi:10.16420/j.issn.0513-353x.2015-0741. [Zhang Y F,Hu C Q,Yang Y,Zhu R S,Guo J,Wang R Z. 2016. Pollen morphology observation of eight resources in Diospyros[J]. Acta Horticulturae Sinica,43(6): 1167-1174.]
赵梦然,陈强,黄晨阳,张金霞,邬向丽. 2012. 中国野生白灵菇遗传多样性的SCoT分析[J]. 园艺学报,39(12): 2475-2482. doi:10.16420/j.issn.0513-353x.2015-0741. [Zhao M R,Chen Q,Huang C Y,Zhang J X,Wu X L. 2012. SCoT Analysis of genetic polymorphism for wild germplasm of Pleurotus eryngii var. tuoliensis from China[J]. Acta Horticulturae Sinica,39(12): 2475-2482.]
钟秋生,陈常颂,林郑和,阮其春,彭艾. 2012. 铁观音等5个茶树种质的花器性状初步分析[J]. 茶叶科学技术,(3): 23-25. doi:10.3969/j.issn.1007-4872.2012.03.006. [Zhong Q S,Chen C S,Lin Z H,Ruan Q C,Peng A. 2012. A preliminary analysis of flower morphological traits of five cultivars such as Tie-guanyin[J]. Tea Science and Technology,(3): 23-25.]
Akagi T,Henry I M,Ohtani H,Morimoto T,Beppu K,Kataoka I,Tao R. 2018. A Y-encoded suppressor of feminization arose via lineage-specific duplication of a cytokinin response regulator in kiwifruit[J]. The Plant Cell,30(4): 780-795. doi:10.1105/tpc.17.00787.
Caseys C. 2018. Shy girl gives kiwifruit male flowers[J]. The Plant Cell,30(4): 739-740. doi:10.1105/tpc.18.00278.
de Costa V B S,de Mendon?a Pimentel R M,das Chagas M D G S,Alves G D,Castro C C D. 2017. Petal micromorphology and its relationship to pollination[J]. Plant Bio-logy,19(2): 115-122. doi:10.1111/plb.12523.
Fortini P,Antonecchia G,Di M P,Maiuro L,Viscosi V. 2013. Role of micromorphological leaf traits and molecular data in taxonomy of three sympatric white oak species and their hybrids (Quercus L.)[J]. Plant Biosystems,149(3): 546-558. doi:10.1080/11263504.2013.868374.
Konarska A. 2017. Comparative micromorphology and anatomy of flowers and floral secretory structures in two Viburnum species[J]. Protoplasma,254(1): 523-537. doi:10.1007/s00709-016-0972-0.
Millner H J,Baldwin T C. 2016. Floral micromorphology of the genus Restrepia (Orchidaceae) and the potential consequences for pollination[J]. Flora-Morphology,Distribution,Functional Ecology of Plants,225:10-19. doi:10. 1016/j.flora.2016.09.007.
Ullah F,Papini A,Shah S N,Zaman W,Sohail A,Iqbal M. 2018. Seed micromorphology and its taxonomic evidence in subfamily Alsinoideae(Caryophyllaceae)[J]. Microscopy Research and Technique,84(4):250-259. doi:10. 1002/jemt.23167.
Yu W T,Jacques F M B,Chen S T,Zhou Z K. 2012. Nutlet micro-morphology of the genus Microula(Boraginaceae) from the Qinghai-Tibetan Plateau,and its systematic implications[J]. Nordic Journal of Botany,30:596-612. doi: 10.1111/j.1756-1051.2011.01336.x.
(責任编辑 邓慧灵)
