2种昆虫病原真菌对草地贪夜蛾体表侵染模式的扫描电镜观察
2021-08-02雷妍圆官昭瑛ShoaibFreedSyedMuhammadZakaAbidHussin章玉苹
雷妍圆 官昭瑛 Shoaib Freed Syed Muhammad Zaka Abid Hussin 章玉苹

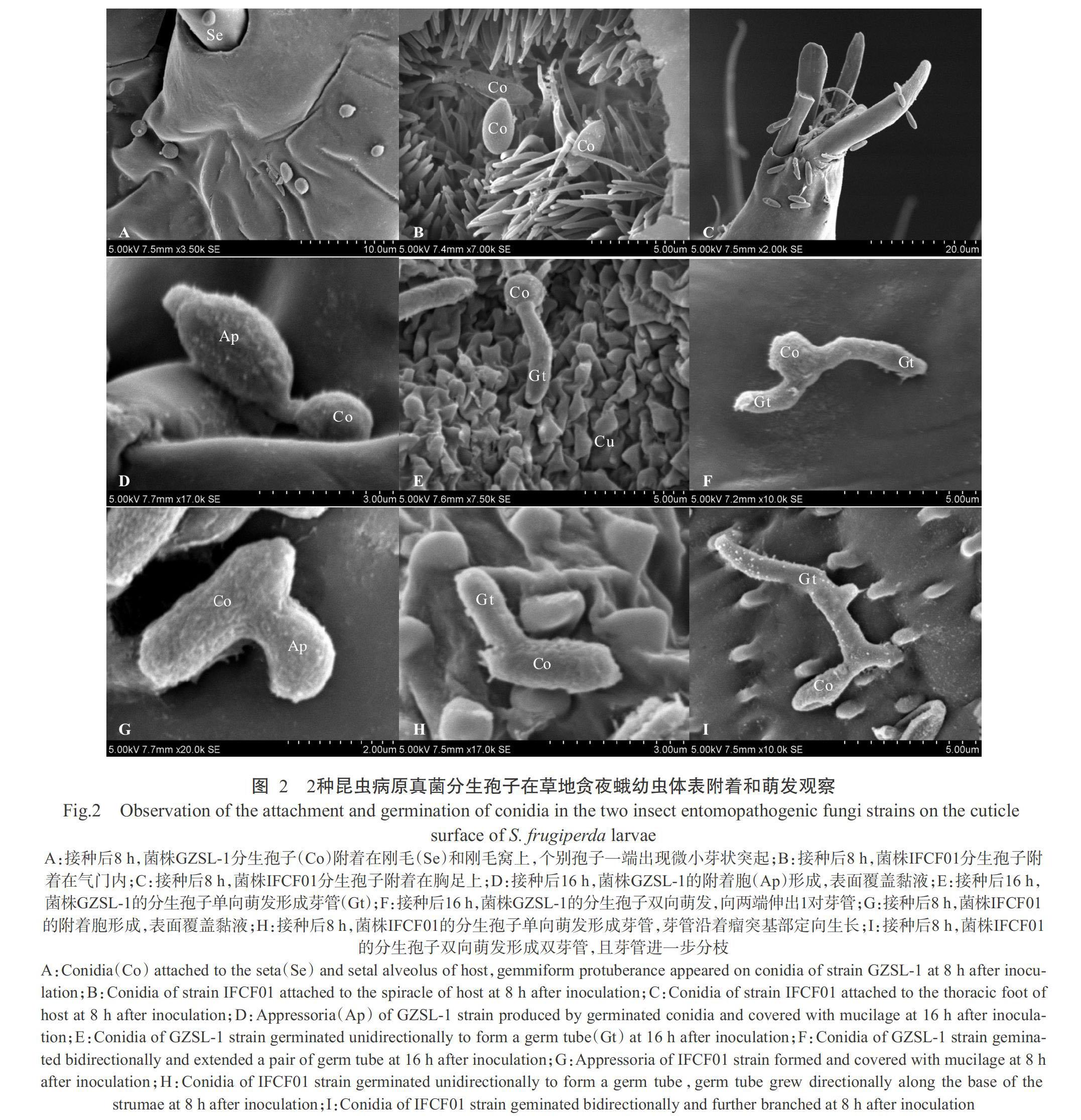
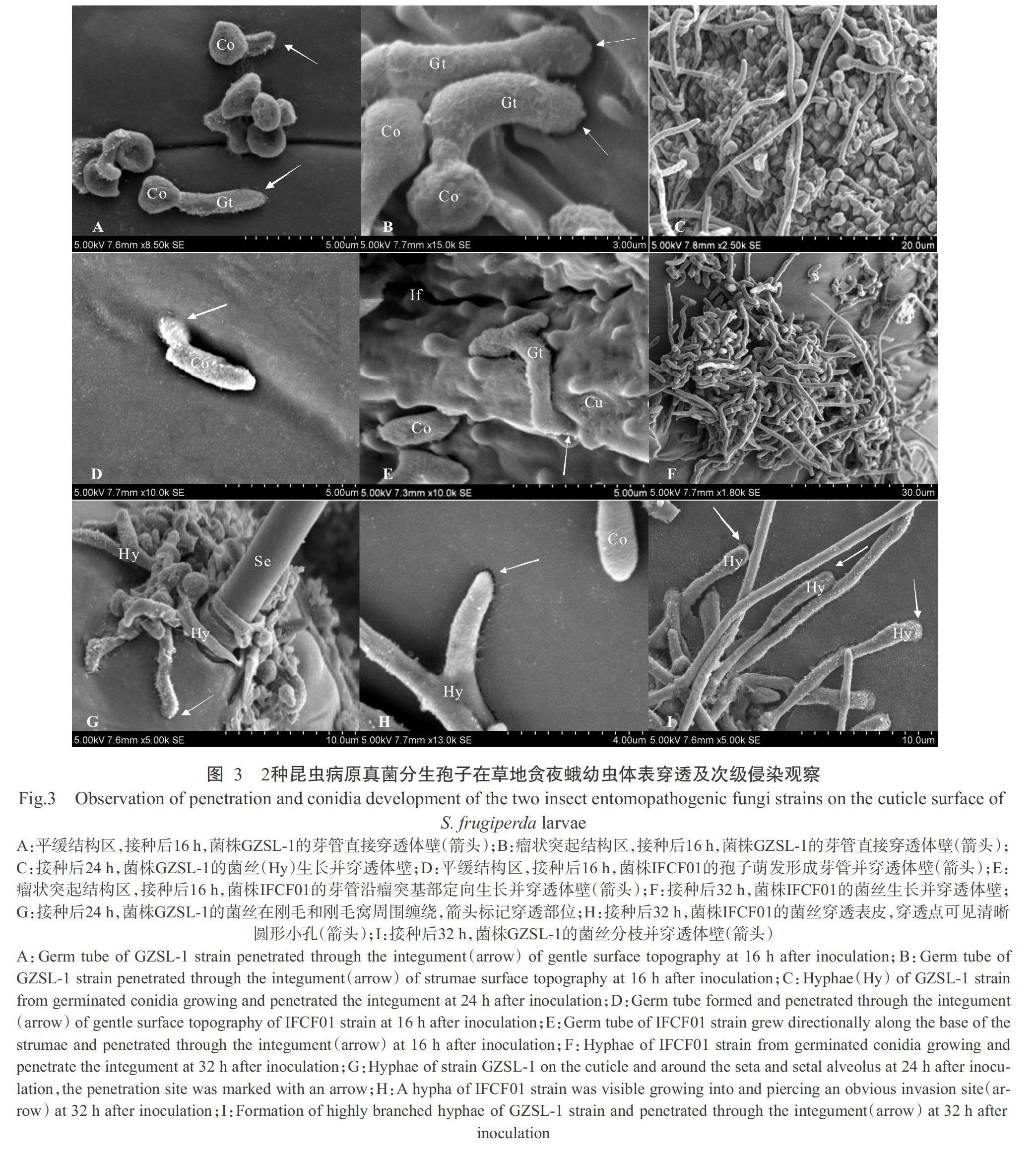
摘要:【目的】在前期篩选获得对草地贪夜蛾高效致病真菌球孢白僵菌(Beauveria bassiana)菌株GZSL-1和玫烟色虫草(Cordyceps fumosorosea)菌株IFCF01的基础上,进一步明确2株菌株在草地贪夜蛾体表的侵染模式,为阐明菌株致病机理,提高昆虫病原真菌对草地贪夜蛾的防效提供理论依据。【方法】利用扫描电镜观察草地贪夜蛾2龄幼虫分别接种菌株GZSL-1和IFCF01(孢子悬浮液浓度为1.0×107孢子/mL)后分生孢子在幼虫体表附着、萌发、穿透和增殖的过程。【结果】菌株GZSL-1和IFCF01萌发后均可形成附着胞,草地贪夜蛾幼虫体表结构对2株菌株分生孢子萌发、附着和穿透芽管长度有不同程度的影响。2株菌株在各结构区均有较高的萌发率(>90.00%)和附着胞率(>80.00%),在平缓结构区比瘤状突起结构区观察到更短的穿透芽管。在各结构区,菌株GZSL-1形成的穿透芽管长度均短于菌株IFCF01相应的芽管长度,在平缓结构区,菌株GZSL-1的穿透芽管长度显著短于菌株IFCF01(P<0.05)。菌株GZSL-1和IFCF01分生孢子接种后分别在16和8 h内萌发,在24和32 h内普遍出现穿透结构穿透草地贪夜蛾幼虫体壁。2株菌株的初级和次级分生孢子分别在接种后72和96 h内出现。【结论】扫描电镜观察证实球孢白僵菌菌株GZSL-1和玫烟色虫草菌株IFCF01对草地贪夜蛾具有高致病性,揭示了病原真菌分生孢子在草地贪夜蛾虫体上的侵染过程,结果为后续2株菌株的改良及田间应用打下理论基础。
关键词: 球孢白僵菌;玫烟色虫草;草地贪夜蛾;体表侵染;扫描电镜
中图分类号: S433.4;Q965.9 文献标志码: A 文章编号:2095-1191(2021)03-0578-11
Mode of infection on the cuticle of Spodoptera frugiperda (Lepidoptera: Noctuidae) by two entomopathogenic fungi species revealed by scanning electron microscopy
LEI Yan-yuan1, GUAN Zhao-ying2, Shoaib Freed3, Syed Muhammad Zaka3,
Abid Hussin4, ZHANG Yu-ping1*
(1 Institute of Plant Protection, Guangdong Academy of Agricultural Sciences/Guangdong Provincial Key Laboratory of High Technology for Plant Protection, Guangzhou 510640, China; 2 Shenzhen Institute of Technology, Shenzhen, Guangdong 518116, China; 3Faculty of Agricultural Sciences and Technology, Bahauddin Zakariya University, Multan 60800, Pakistan; 4College of Agricultural and Food Sciences, King Faisal University, Hofuf 31982, Saudi Arabia)
Abstract:【Objective】In this study,based on the previous screening of the high pathogenic Beauveria bassiana strain GZSL-1 and Cordyceps fumosorosea strain IFCF01 of Spodoptera frugiperda, the purpose of this study was to better understand the mode of action of these entomopathogenic fungi on the cuticle of S. frugiperda, and provide a theoretical basis for elucidating the pathogenicity mechanism of these two strains in order to achieve a constant and high control efficacy. 【Method】The attachment, germination, penetration and conidial reproduction of B. bassiana and C. fumosorosea to 2nd instar larvae of S. frugiperda infected with strains GZSL-1 and IFCF01(1.0×107 conidia/mL) were observed using scanning electron microscope(SEM). 【Result】Germinated conidia of both strains formed appressoria, and surface topography of S. frugiperda larvae affected germination of conidia, attachment and the length of penetrating germ tube. The two strains showed similarities in terms of high germination rate(>90.00%) and appressoria rate(>80.00%). There were shor-ter germ tubes grew before penetrating on gentle surface topography than on strumae surface topography. Strain IFCF01 exhibited longer penetrating germ tubes compared to GZSL-1 on all surface topographies. Besides, strain GZSL-1 exhibited significantly shorter penetrating germ tubes compared to strain IFCF01 on gentle surface topography(P<0.05). Strains GZSL-1 and IFCF01 began to germinate within 16 and 8 h, and penetrations were commonly observed within 24 and 32 h. After 72 and 96 h post inoculation, primary and secondary conidiophore emerged and the mycelial covered the entire cuticular surface. 【Conclusion】SEM observation reveals the infection process of the highly pathogenic B. bassiana strain GZSL-1 and C. fumosorosea strain IFCF01 against S. frugiperda, the results of this study will further provide the theoretical basis and reference for strain improvement and field application.
Key words: Beauveria bassiana; Cordyceps fumosorosea; Spodoptera frugiperda; cuticle infection; scanning electron microscopy
Foundation item: National Key Research and Development Program of China(2019YFD0300104); Science and Technology Planning Project of Guangdong(2020A1414010040); International Science and Technology Organization Cooperation Project of “One Belt And One Road” of Guangzhou Association for Science & Technology(G20210101005); Scientific Research Foundation of Shenzhen Institute of Technology(2111016)
0 引言
【研究意义】生物防治是害虫综合治理(IPM)的基本策略,在各种常用的生物防治技术中,基于昆虫病原真菌的微生物防治有助于减轻化学农药对人、畜和天敌昆虫的毒害,促进生态平衡的良性循环,有利于绿色农产品的生产及以出口创汇为目标的农业产业化(何恒果等,2004;杨普云等,2018;张元臣等,2019;张志春等,2020)。病原真菌防治害虫的过程,是一种生命替代另一种生命的过程。不像其他病原微生物(如细菌、病毒)需要被寄主摄取,真菌通过主动侵染控制虫体而不依赖寄主的生活习性。真菌接触至寄主昆虫体表后,需要在一定的温、湿度下经一定时间侵入,因此,昆虫病原真菌侵染成功与否,与分生孢子在寄主体表的侵染过程密切相关(王音等,2005)。分生孢子对寄主表皮的非特异性附着是侵染过程的起始事件(Rangel et al.,2008;Chouvenc et al.,2009),随后孢子萌发产生芽管或附着胞,生长为菌丝(Amóra et al.,2010),依靠这些侵染构造穿透寄主体壁进入体腔,克服寄主细胞和体液免疫防御,吸取寄主体内的养分而生长、增殖,在适宜的环境条件下,菌体从寄主体内穿出产生新的分生孢子并引起水平传播(Khun et al.,2021)。昆虫表皮是抵御病原真菌侵染的第一道屏障(Butt et al.,1995),由于不同昆虫间表皮的物理结构和化学组成存在差异,导致一种病原真菌对不同种类昆虫或同种昆虫不同虫态、龄期的防效不同,表现为菌株对不同靶标昆虫的致病力差异和寄主专化现象(蒲顺昌等,2013)。草地贪夜蛾(Spodoptera frugiperda)是热带和亚热带许多国家的重大迁飞性害虫,其具有寄主范围广泛、取食能力强和抗药性严重等特點,防控难度大(Casmuze et al.,2010;Early et al.,2018;Montezano et al.,2018;姜玉英等,2019;张磊等,2019)。自2019年我国首次发现草地贪夜蛾入侵,各省(区、市)、地级市的科研院所及有关部门加紧了对其应急预案及防控工作长效机制的构建,利用昆虫病原真菌对该虫进行微生物防治具有良好应用前景,是当前防控中的一个研究热点。【前人研究进展】球孢白僵菌(Beauveria bassiana)和玫烟色虫草(Cordyceps fumosorosea)是研究和应用较为广泛的昆虫病原真菌,全球先后有171个真菌杀虫剂产品问世,其中以球孢白僵菌和玫烟色虫草为有效成分的制剂分别占33.9%和5.8%(de Faria and Wraight,2007;Hussain et al.,2014),在多种农林害虫微生物防治中发挥了重要作用(蒲蛰龙和李增智,1996;Zimmermann,2008;Arthurs and Dara,2019)。关于球孢白僵菌和玫烟色虫草与草地贪夜蛾的互作,国外已有大量且系统的研究报道(Bouamama et al.,2010;Akutse et al.,2020;Corrêa et al.,2020;González et al.,2020;Russo et al.,2020),国内相关研究才刚起步,特别是本地病原真菌与草地贪夜蛾互作体系中菌体侵染模式与寄主致病性的相关性尚无报道。在昆虫病原真菌常见侵染结构中,金龟子绿僵菌(Metarhizium ani-sopliae)分生孢子通过形成附着胞结构对西花蓟马(Frankiniella occidentalis)表皮进行穿透,类似的还有淡紫拟青霉(Paecilomyces lilacinu)对爪哇根结线虫(Meloidogyne javanica)(Holland et al.,2002)、两型蜡蚧菌(Lecanicillium dimorphum)对棕榈红蚧(Phoenicococcus marlatti)(Asensio et al.,2005)和蜡蚧轮枝菌(L. lecanii)对拟褐圆蚧(Coccus hesperidum)(Liu et al.,2011)等。其他的诸如舞毒蛾噬虫霉(Entomophaga maimaiga)和球孢白僵菌,直接以菌丝或芽管通过表皮上的气孔进行穿透,而不需要形成附着胞(Hajek and Eastburn,2003;Asensio et al.,2005;Mauchline et al.,2011)。不同种类病原真菌或同种病原真菌不同菌株在侵染速度上也存在差异。金龟子绿僵菌侵染小菜蛾(Plutella xylostella)约在接种后22 h以穿透钉侵入表皮(王音等,2005),侵染微小扇头蜱(Rhipicephalus microplus)在接种后48 h观察到以芽管从排泄孔直接穿透(Bernardo et al.,2018);玫烟色虫草侵染菜青虫(Pieris rapae)(张奂等,2007)和小菜蛾(Lei et al.,2021)均在接种后24 h以芽管穿透体壁进入寄主血腔;球孢白僵菌接种玉米螟(Ostrinia nubilalis)后24 h可观察到大量孢子萌发并以芽管或菌丝穿透体壁(刘忱和郭志红,2019),而接种光肩星天牛(Anoplophora glabripennis)则需48 h才出现孢子萌发和穿透现象(邓彩萍等,2012)。【本研究切入点】本课题组通过前期对草地贪夜蛾昆虫病原真菌种类及防治潜能的评估(雷妍圆等,2020a,2020b),鉴定出对草地贪夜蛾具有高致病性的病原真菌球孢白僵菌菌株GZSL-1和玫烟色虫草菌株IFCF01,其在1.0×107孢子/mL的孢子浓度下对草地贪夜蛾2龄幼虫的致死中时(LT50)分别为2.77和2.22 d,致死中浓度(LC50)分别为1.17×105和3.42×103孢子/mL,表现出极大的生防潜力,但关于这2种病原真菌在草地贪夜蛾体表侵染模式尚未开展相关研究,不同菌株间孢子萌发行为、侵入方式的差异仍不清楚。【拟解决的关键问题】以球孢白僵菌菌株GZSL-1和玫烟色虫草菌株IFCF01为材料,利用扫描电镜对球孢白僵菌和玫烟色虫草侵染草地贪夜蛾幼虫过程中侵染结构的形成和表皮穿透过程进行超微结构研究,以明确高效菌株对草地贪夜蛾的侵染模式,探讨寄主与病原真菌间的互作关系,为阐明昆虫病原真菌的生态适应性和毒力进化,提高昆虫病原真菌对草地贪夜蛾的防效提供理论依据。
1 材料与方法
1. 1 供试虫源和菌株培养
草地贪夜蛾幼虫采集自广东省广州市白云区钟落潭镇广东省农业科学院白云基地种植的粤甜28甜玉米(Zea mays L.)植株上,在实验室用玉米叶和果穗饲养至化蛹,待其羽化产卵后建立实验室种群作为供试虫源。室内饲养条件为(26±1)℃,相对湿度60%~90% ,光周期L∶D=14 h∶10 h。
供试球孢白僵菌菌株GZSL-1和玫烟色虫草菌株IFCF01保存于广东省农业科学院植物保护研究所,于-80 ℃下保藏。菌株接种于萨氏(SDAY)培养基(葡萄糖40 g/L,蛋白胨10 g/L,酵母膏10 g/L,琼脂20 g/L),于温度(28±1)℃,相对湿度(70±5)%,全黑暗的恒温培养箱中培养20 d后供试。
1. 2 接种及取样方法
用灭菌软毛笔将SDAY培养基上的供试菌株分生孢子粉刷入装有灭菌0.05%吐温-80溶液的烧杯(50 mL)中,经涡流振荡,点样血球计数板,配制浓度为1.0×107孢子/mL的孢子悬浮液,作为接种的菌液。每种菌株为1个处理,以灭菌0.05%吐温-80溶液处理为对照。采用浸虫法处理草地贪夜蛾幼虫,选取个体大小一致的草地贪夜蛾2龄幼虫,放入不同菌株孢子悬浮液浸渍10 s后挑出,置于滤纸上吸去多余水分,移至皿底垫有湿润滤纸片的培养皿(d=7.5 cm)中集体饲养(5头/皿)(王道通等,2020),皿内放入新鲜玉米叶供其取食。每个处理25头幼虫,3次重复,处理后的幼虫置于人工气候箱中饲养[温度(26±1)℃,相对湿度(80±5)%,光周期L∶D=14 h∶10 h],每天观察幼虫存活情况及感染病虫的外部特征,将死亡幼虫保湿培养,根据前期研究结果(雷妍圆等,2020a,2020b),分别于接种后第8、16、24、32、48、72和96 h依次取样,将样品置于2.5%戊二醛进行前固定。
1. 3 扫描电镜样品制备及观察
取1.2中收集和固定好的感菌草地贪夜蛾幼虫,用0.1 mol/L磷酸缓冲液漂洗3次;1%锇酸后固定,再次用0.1 mol/L磷酸缓冲液漂洗3次;然后以50%、70%、80%、90%、95%和100%的乙醇梯度脱水,以醋酸异戊酯过渡,最后将样品置于临界点干燥仪干燥,再将干燥好的样品粘台,以离子溅射仪喷金镀膜,在扫描电镜(S-3400N,Hitachi)下观察拍照。
1. 4 分生孢子生长参数测量及数据处理
当分生孢子的芽管长度与分生孢子宽度相等时,视为分生孢子已萌发(Safavi et al.,2007)。当牙管末端膨大达芽管宽度的1.5倍以上时,即视为形成附着胞。穿透芽管的长度为从分生孢子萌发点到寄主表皮穿透点间的长度。各菌株接种后16 h,分别在草地贪夜蛾幼虫体表不同结构区统计萌发孢子数、附着胞形成数和孢子总数,计算孢子萌发率和附着胞率,每个结构区选取25个分生孢子,每株菌株处理重复3头幼虫。接种后32 h,测量不同菌株处理的幼虫体表不同结构区内穿透芽管长度,在电镜视野下于虫体每种结构区内随机测量5个分生孢子,同一种结构区共计15个,每种菌株处理重复3头幼虫。
试验数据经Excel 2013整理后,使用SPSS 20.0进行处理分析,采用单因素方差分析(One-way ANOVA)进行差异显著性分析及Tukey HSD事后检验(α=0.05)。
2 结果与分析
2. 1 草地贪夜蛾幼虫体表构造特点
在电镜视野下,草地贪夜蛾幼虫不同部位体表外形构造存在较大差异,根据外形结构特征,将其分为瘤状突起结构区和平缓结构区2类(图1)。瘤状突起结构区包括体节背、腹面及靠近刚毛窝、气门的区域,其特点是体表由许多排列较规则的瘤状或刺瘤状突起构成,突起基部明显分离,突起间存在短距离的平缓区域,平缓区域大小和瘤的长度因具体部位而异(图1-A)。平缓结构区包括头壳、背板、胸足、腹足、臀足、足节间膜、足末端、趾钩、气门、刚毛窝、刚毛表面和臀节等区域,其特点是表面平滑或仅有微小起伏(图1-B)。
2. 2 球孢白僵菌和玫烟色虫草在草地贪夜蛾幼虫体表附着和萌发
电镜视野下,2种病原真菌的分生孢子均能附着在草地贪夜蛾幼虫体表不同区域,包括刚毛(图2-A)、刚毛窝(图2-A)、气门内(图2-B)及胸足(图2-C)等部位。2株菌株表现出较为相似的孢子萌发行为。接种菌株GZSL-1和IFCF01后,孢子分别在16和8 h萌发(图2-A)。孢子萌发过程中产生2种类型的芽管。第1种,分生孢子的芽管顶端膨大产生附着胞结构(图2-D和图2-G),紧紧固着在幼虫表皮上;芽管具有向幼虫体表定向生长的能力,并进一步发育形成单芽管(图2-E和图2-H)。第2种,孢子萌发后形成双芽管(图2-F和图2-I),芽管沿着表皮生长较长,且在菌株IFCF01的芽管上还观察到进一步出现分枝(图2-I)。2种病原真菌的分生孢子和附着胞表面皆可见黏液层(图2-D和图2-G)。
2. 3 球孢白僵菌和玫烟色虫草在草地贪夜蛾幼虫体表的菌丝生长及穿透过程
菌株GZSL-1和IFCF01均观察到在草地贪夜蛾幼虫体表各结构区上的穿透行为。如图3所示,2种病原真菌分别在平缓结构区(图3-A和图3-D)和瘤状突起结构区(图3-B和图3-E)以芽管直接穿透幼虫表皮,或芽管以原有直径延伸较长距离,形成菌丝并超过孢子纵轴长度,经几次弯折沿幼虫体壁方向定向生长,当到达适合入侵位置时,以菌丝直接穿透(图3-C和图3-F)。至接种后24 h,穿透行为普遍增多,在幼蟲体表各区域包括刚毛和刚毛窝附近也观察到菌丝缠绕和穿透现象,芽管端部与幼虫体壁接触处可见黏液层(图3-G)。随着菌丝的生长,至接种后32 h,菌丝形成次级分枝进行穿透,穿透点可见穿透孔(图3-H和图3-I)。
2. 4 球孢白僵菌和玫烟色虫草在草地贪夜蛾幼虫体表的次级侵染
昆虫病原真菌穿透草地贪夜蛾幼虫表皮进入体内,虫体内部提供的营养被大量消耗,菌体从幼虫体内穿出,在体外继续生长。至接种后48 h,2种病原真菌的菌丝体几乎包围了整个虫体(图4-A和图4-D);接种后72 h,可见初级分生孢子形成(图4-B和图4-E)。菌株GZSL-1分生孢子梗着生于营养菌丝上,产孢细胞在菌丝上簇生,基部为球形、近球形,产孢轴较长,轴上具小齿突,呈“之”字形弯曲(图4-B)。菌株IFCF01的菌丝分隔,光滑;分生孢子梗直立,产生于菌丝上;瓶梗规则着生在分生孢子梗上,呈梭形,基部椭圆形膨大,向上逐渐变细形成一个细的颈部(图4-E)。接种后96 h,新产生的分生孢子聚集成簇,此时2株菌株均出现二级至多级产孢。菌株GZSL-1分生孢子透明、光滑,球形或近球形,大小为1.94 ?m×1.62 ?m(圖4-C),菌株IFCF01分生孢子表面光滑,长椭圆形,大小为3.69 μm×1.20 μm(图4-F)。
2. 5 球孢白僵菌和玫烟色虫草在草地贪夜蛾幼虫体表的生长指标
由图5可知,2种昆虫病原真菌在草地贪夜蛾幼虫表皮各结构区均有较高的分生孢子萌发率(>90.00%),且菌株间无显著差异(P>0.05,下同)(图5-A)。与菌株GZSL-1相比,菌株IFCF01的附着胞率稍高,菌株GZSL-1和IFCF01在瘤状突起结构区及平缓结构区的附着胞率分别为(85.33±2.31)%、(82.67±4.62)%和(92.00±6.93)%、(88.00±4.00)%,但2株菌株及各结构区间无显著差异,均高于80.00% (图5-B)。对不同结构区分生孢子萌发后形成的芽管穿透幼虫体壁时的长度进行测量,结果表明2株菌株的穿透长度均以瘤状突起结构区最长,平缓结构区的芽管最短。同时,菌株GZSL-1在各结构区形成的穿透芽管长度均短于菌株IFCF01相应的芽管长度,特别是在平缓结构区的芽管长度差异最大,显著短于菌株IFCF01(P<0.05)(图5-C)。
3 讨论
昆虫病原真菌侵染与寄主防御在恒定的相互选择压力下进行,两者的相互作用有可能引起菌株变异、寄主范围变化,以及(低毒)菌株经由寄主“继代”时出现毒力增强(即复壮)的现象。寄主表皮作为虫菌互作的第一个接触点和屏障,对整个侵染过程起决定性作用,即最终决定菌体成功感染或寄主成功抵御真菌(Ortiz-Urquiza and Keyhani,2013)。前人研究表明,寄主表皮结构对昆虫病原真菌的附着萌发和致病性有重要影响(Butt et al.,1995)。王音等(2005)报道金龟子绿僵菌入侵小菜蛾有利部位为表皮平缓结构区的足部与头壳部位,其分生孢子在头壳萌发后芽管能立即膨大形成附着胞直接刺入,而在表皮起伏变化大的嵴状突起结构区,芽管无法迅速发现合适的附着和穿透区域,需经较长时间定向生长才会发生穿透或依表皮结构形成附着胞后穿透,因而从该类区域入侵时间较平缓区略为滞后。另有不同观点认为,昆虫的幼虫头壳强烈骨骼化,该区域表皮结构光滑且坚硬,不利于分生孢子附着。幼虫蜕皮从头部开始,孢子如未能及时附着,即首先被脱去,因而头壳并非入侵最有利的薄弱部位。而在表皮有突起的结构区,这些突起恰好成为卡住孢子的良好构造,可在一定程度上防止孢子掉落,反而有利于分生孢子附着和侵染(Lei et al.,2021)。蜕皮作为昆虫的一种自我防御,可最大限度降低分生孢子在其表皮上定殖。作为回应,真菌必须快速萌发穿透,以逃避寄主的防御行为反应。从本研究结果来看,2种昆虫病原真菌的分生孢子均观察到高密度分布在草地贪夜蛾幼虫体表瘤状突起结构区和平缓结构区,说明菌株对草地贪夜蛾幼虫体壁具有无差别性的迅速侵染能力,可攻击的位点较多,很可能是2株菌株对草地贪夜蛾致病力较强且致病速度较快的原因之一(雷妍圆等,2020a,2020b)。
根据侵染过程的时间轴,分生孢子在寄主体表的附着和萌发,是昆虫病原真菌成功侵染的先决条件。分生孢子萌发主要受3个因素影响:(1)较高的湿度(Liu and Stansly,2009);(2)遇到适合的寄主体壁,因为只有在特异性寄主的体壁上才有刺激孢子萌发的物质(Gillespie et al.,2002);(3)必须克服昆虫体壁上某些物质如抗菌肽、脂肪酸等的抑菌作用(Latgé et al.,1987)。附着胞的产生是大部分昆虫病原真菌在寄主上定殖的前提,对于建立病原与寄主间的关系极其重要。有研究表明分生孢子在寄主表皮上的附着能力越强,其致病力也越强(Altre et al.,1999)。也有研究认为孢子对寄主的致病力与孢子的萌发率、附着数量和附着胞形成之间无相关性,而是取决于侵染速度(Ibrahim et al.,2002;Ment et al.,2010)。据报道,在烟粉虱(Bemisia tabaci)若虫不同表皮结构区域观察到球孢白僵菌分生孢子不同的萌发行为,在某些区域,孢子多为单向萌发,而另一些区域则出现单、双向萌发,不同结构区域表皮理化特性与分生孢子间的作用,导致这种不同的萌发反应(Liu et al.,2010)。从本研究来看,2株菌株的分生孢子均能产生单芽管或双芽管,均可形成附着胞结构,且单、双芽管在各结构区皆有分布,并无特定的选择偏好性,亦表明附着胞的形成为非专化性,这是否可以认为是菌株的一种高效侵染策略仍有待进一步研究。还有研究表明,单向萌发的分生孢子能产生强壮的芽管,且大多发育为附着胞结构,而双向萌发的分生孢子则更倾向于生长为无穿透行为的菌丝,这种不同的萌发极性可作为致病力的一个指标(Talaei-Hassanloui et al.,2007)。本研究观察到的单附着胞现象与上述萌发方式较为吻合,也与Ghaffari等(2017)的报道一致,2株菌株均未出现双附着胞,且单附着胞均由单向萌发的芽管形成,双向萌发的芽管未出现穿透行为,而是通过芽管进一步延长,在草地贪夜蛾幼虫表皮上形成菌丝网络结构。此外,电镜视野下,2株菌株在草地贪夜蛾幼虫表皮附着部位的附着胞分泌黏液物质,提示寄主表皮的成分刺激了分生孢子活化,帮助其在体壁上附着,提供有利于胞外酶活动的环境,在穿透寄主表皮过程中发挥活性(Safavi,2010;Khan et al.,2016)。
分生孢子在寄主体表穿透的速度及数量,是昆虫病原真菌在寄主体表成功定殖和致病寄主昆虫的关键环节(Lei et al.,2021)。绿僵菌分生孢子通常以附着胞结构对寄主表皮进行穿透(St. Leger et al.,1991;樊美珍和李增智,1994)。而本研究中2株菌株均可以附着胞、芽管或菌丝等形式直接穿透草地贪夜蛾幼虫表皮,其入侵形式较多,很可能是影响其致病效率的因素之一。草地贪夜蛾幼虫体表不同结构区对附着胞的产生和穿透菌丝的长度有一定影响。2株菌株在草地贪夜蛾幼虫体表各结构区萌发后芽管末端很快膨大形成附着胞,或以芽管直接入侵,其萌发率和附着胞率均较高。该结果进一步证实了Butt等(1995)报道的菌株致病力不仅取决于孢子的萌发,还有分生孢子在体表上的附着力。但并不是所有的昆虫病原真菌都必须产生附着胞后才能侵染寄主昆虫。如金龟子绿僵菌在辣根猿叶甲(Phaedon cochleariae)和油菜金头跳甲(Psylliodes chrysocephaIa)体表产生的附着胞比在桃蚜(Myzus persicae)和萝卜蚜(Lipaphis erysimi)上的多,但在蚜虫上即便没有附着胞产生,萌发的芽管同样也能侵入蚜虫表皮(Butt et al.,1995);球孢白僵菌侵染马铃薯甲虫(Leptinotarsa decemlineata)时通过附着胞和芽管穿透(Duan et al.,2017),但在侵染棉铃虫(Heliothis zea)时并没有形成附着胞结构(Pekrul and Grula,1979);玫烟色虫草可以附着胞、芽管或菌丝等多种形式侵染小菜蛾(Lei et al.,2021)。此外,本研究还观察到菌株IFCF01分生孢子可经由幼虫体壁上的气门进入。类似现象在球孢白僵菌对棉铃虫的侵染模式中也有报道(Pekrul and Grula,1979)。与之相反的是,关于球孢白僵菌对桑天牛(Apriona germari)的研究结果显示气门并非理想的突破口,因为气门中的过滤结构成为阻止孢子进入的天然屏障(王晓红等,2009)。
本研究的菌株GZSL-1和IFCF01在草地贪夜蛾2龄幼虫体表萌发起始时间稍有差别,菌株IFCF01最早观察到萌发是在接种后8 h,菌株GZSL-1是在接种后16 h ,但菌株GZSL-1在接种后24 h已有较多穿透体壁的行为发生,活跃穿透期在24~32 h,且在平缓结构区穿透表皮芽管长度显著短于菌株IFCF01。菌株IFCF01在接种后32 h 才出现较多穿透行为,活跃穿透期为32~48 h,在各结构区穿透芽管长度较前者长。说明菌株GZSL-1比菌株IFCF01对草地贪夜蛾2龄幼虫具有更快的侵染速度。此前关于2株菌株的生测结果(雷妍圆等,2020a,2020b)显示,菌株GZSL-1对草地贪夜蛾2 龄幼虫的LT50比菌株IFCF01快0.55 d,与本研究的观察结果相吻合。此外,对穿透寄主表皮的芽管长度测量结果表明,2株菌株均以平缓结构区的芽管最短,明显短于瘤状突起结构区,说明在该部分区域入侵所用的时间最短,速度最快。菌株能快速侵入幼虫表皮平缓结构区的头壳,意味着可迅速损伤幼虫脑组织,破坏其神经系统,这有可能是2株菌株快速侵染寄主的策略之一
真菌對寄主的侵染主要有两种机制,一种是穿透寄主表皮的机械压力,如孢子萌发时芽管定向生长产生的压力(Fang et al.,2009);另一种是酶类物质对表皮的降解作用,如次级代谢物、胞外蛋白酶、几丁质酶和酯酶等(Zhang et al.,2008;Safavi,2010;Staats et al.,2013;Khan et al.,2016)。前者由昆虫病原真菌结构发挥作用,如本研究扫描电镜观察到的附着胞结构,后者仍有待进一步探究,以评估虫体诱导下菌株产生的次生代谢物和酶类物质在侵染过程中所发挥的作用。另外,真菌对昆虫控制能力强弱不仅体现在致病力上,其自然环境温度、紫外线耐受性等因素均会影响田间防治效果。可以预期的是,昆虫病原真菌的生活环境相似,自然条件下能混杂发生,如果几种病原真菌能够对同一生境内的害虫同时起控制作用,势必在生物防治中起事半功倍的作用。
4 结论
本研究结果在室内可控条件下获得,2种病原真菌球孢白僵菌和玫烟色虫草在草地贪夜蛾体表侵染模式观察证实了菌株的高致病性,基本阐明了设置条件下菌株分生孢子侵染草地贪夜蛾幼虫的表观过程,为广东本地的草地贪夜蛾生防真菌球孢白僵菌菌株GZSL-1和玫烟色虫草菌株IFCF01的改良和田间应用打下理论基础。
参考文献:
邓彩萍,闫喜中,刘红霞,郝赤,骆有庆. 2012. 球孢白僵菌侵染光肩星天牛幼虫的扫描电镜及组织病理观察[J]. 林业科学,48(3):105-109. doi:10.11707/j.1001-7488.20120317. [Deng C P,Yan X Z,Liu H X,Hao C,Luo Y Q. 2012. Pathological observation of Anoplophora glabripennis larva infected by Beauveria bassiana by using SEM and light microscope[J]. Scientia Silvae Sinicae,48(3): 105-109.]
樊美珍,李增智. 1994. 营养物和培养条件对虫生真菌附着胞形成的影响[J]. 安徽农业大学学报,21(2):123-130. [Fan M Z,Li Z Z. 1994. Impact of nutrients and culture conditions on appresorium formation of entomogenous fungi[J]. Journal of Anhui Agricultural University,21(2):123-130.]
何恒果,李正跃,陈斌,文良柱. 2004. 虫生真菌对害虫防治的研究与应用[J]. 云南农业大学学报,19(2): 167-173. doi:10.16211/j.issn.1004-390x(n).2004.02.011. [He H G,Li Z Y,Chen B,Wen L Z. 2004. Current status in basic and applied research on entomopathogenic fungi for pest mana-gement[J]. Journal of Yunnan Agricultural University,19(2): 167-173.]
姜玉英,刘杰,朱晓明. 2019. 草地贪夜蛾侵入我国的发生动态和未来趋势分析[J]. 中国植保导刊,39(2): 33-35. doi:10.3969/j.issn.1672-6820.2019.02.006. [Jiang Y Y,Liu J,Zhu X M. 2019. Analysis on the occurrence dynamics of invasion and future trend of fall armyworm Spodoptera frugiperda in China[J]. China Plant Protection,39(2): 33-35.]
雷妍圆,吕利华,王裕华,王德森. 2020a. 一株玫烟色虫草对草地贪夜蛾的致病性研究[J]. 环境昆虫学报,42(1): 68-75. doi:10.3969/j.issn.1674-0858.2020.01.9. [Lei Y Y,Lü L H,Wang Y H,Wang D S. 2020a. Pathogenicity study of a strain of Cordyceps fumosorosea to Spodoptera frugiperda(Lepidoptera: Noctuidae)[J]. Journal of Environmental Entomology,42(1): 68-75.]
雷妍圆,章玉苹,薛志洪,王裕华,黄少华,吕利华. 2020b. 一株球孢白僵菌的分离鉴定及其对草地贪夜蛾的致病性[J]. 环境昆虫学报,42(3): 593-601. doi:10.3969/j.issn. 1674-0858.2020.03.10. [Lei Y Y,Zhang Y P,Xue Z H,Wang Y H,Huang S H,Lü L H. 2020b. Isolation and identification of a Beauveria bassiana isolate and its pathogenicity to Spodoptera frugiperda (Lepidoptera: Noctuidae)[J]. Journal of Environmental Entomology,42(3): 593-601.]
劉忱,郭志红. 2019. 球孢白僵菌侵染玉米螟的电镜观察[J]. 电子显微学报,38(2): 144-149. doi:10.3969/j.issn.1000-6281.2019.02.009. [Liu C,Guo Z H. 2019. Electron microscopic observations on Beauveria bassiana infecting Ostrinia nubilalis[J]. Journal of Chinese Electron Microscopy Society,38(2): 144-149.]
蒲顺昌,秦丽,陈名君,蔡悦,黄勃. 2013. 马尾松林中球孢白僵菌寄主转移和专化性的SSR标记分析[J]. 菌物学报,32(4): 698-709. doi:10.13346/j.mycosystema.2013.04.004. [Pu S C,Qin L,Chen M J,Cai Y,Huang B. 2013. Host shift and host specificity analysis of Beauveria bassiana in Massons pine plantation based on SSR molecular marker[J]. Mycosystema,32(4): 698-709.]
蒲蛰龙,李增智. 1996. 昆虫真菌学[M]. 合肥: 安徽科学技术出版社. [Pu Z L,Li Z Z. 1996. Insect mycology[M]. Hefei: Anhui Science and Technology Press.]
王道通,张蕾,程云霞,江幸福. 2020. 草地贪夜蛾幼虫龄期对自相残杀行为的影响[J]. 植物保护,46(3): 94-98. doi:10. 16688/j.zwbh.2019589. [Wang D T,Zhang L,Cheng Y X,Jiang X F. 2020. Larval stage related cannibalism in the fall armyworm,Spodoptera frugiperda[J]. Plant Protection,46(3): 94-98.]
王晓红,黄大庄,杨忠岐,李会平,郑建伟. 2009. 白僵菌感染桑天牛幼虫致病过程的显微观察[J]. 蚕业科学,35(2): 374-378. doi:10.3969/j.issn.0257-4799.2009.02.025. [Wang X H,Huang D Z,Yang Z Q,Li H P,Zheng J W. 2009. Microscopic observations of infection process of Beauveria bassiana on the cuticle of Apriona germari larvae[J]. Science of Sericulture,35(2): 374-378.]
王音,雷仲仁,张青文,问锦曾. 2005. 绿僵菌侵染小菜蛾体表过程的显微观察[J]. 昆虫学报,48(2):188-193. doi:10. 16380/j.kcxb.2005.02.007. [Wang Y,Lei Z R,Zhang Q W,Wen J Z. 2005. Microscopic observations of infection process of Metarhizium anisopliae on the cuticle of the diamondback moth,Plutella xylostella[J]. Acta Entomologica Sinica,48(2):188-193.]
杨普云,王凯,厉建萌,李文星,尹俊梅. 2018. 以农药减量控害助力农业绿色发展[J]. 植物保护,44(5): 95-100. doi:10.16688/j.zwbh.2018276. [Yang P Y,Wang K,Li J M,Li W X,Yin J M. 2018. Promoting green agricultural development through eliminating pesticide overuses in crop pest management[J]. Plant Protection,44(5): 95-100.]
張奂,张仙红,张未仲,张亚丽. 2007. 玫烟色拟青霉对菜青虫的侵染及致病作用[J]. 植物保护,33(2):64-67. doi:10. 3969/j.issn.0529-1542.2007.02.017. [Zhang H,Zhang X H,Zhang W Z,Zhang Y L. 2007. Histopathological chan-ges of the larvae of Pieris rapae infected by Paecilomyces fumosoroseus[J]. Plant Protection,33(2):64-67.]
张磊,靳明辉,张丹丹,姜玉英,刘杰,吴孔明,萧玉涛. 2019. 入侵云南草地贪夜蛾的分子鉴定[J]. 植物保护,45(2): 19-24. doi:10.16688/j.zwbh.2019121. [Zhang L,Jin M H,Zhang D D,Jiang Y Y,Liu J,Wu K M,Xiao Y T. 2019. Molecular identification of invasive fall armyworm Spodoptera frugiperda in Yunnan Province[J]. Plant Protection,45(2): 19-24.]
张元臣,蔡新,张国强,董丽丽,王景顺. 2019. 黏虫溶菌酶基因Mswly的克隆与真核表达[J]. 河南农业科学,48(5):70-77. doi:10.15933/j.cnki.1004-3268.2019.05.011. [Zhang Y C,Cai X,Zhang G Q,Dong L L,Wang J S. 2019. Cloning and eukaryotic expression of Mswly in oriental army-worm, Mythimna separata[J]. Journal of Henan Agricultural Sciences,48(5):70-77.]
张志春,张怡,沈迎春,郭慧芳. 2020. 杀虫真菌爪哇棒束孢对非洲菊烟粉虱作用特点和控害效果[J]. 江苏农业学报,36(6):1398-1402. doi:10.3969/j.issn.1000-4440.2020.06. 007. [Zhang Z C,Zhang Y,Shen Y C,Guo H F. 2020. Characteristics and control effects of insecticidal fungus Isaria javanica against Bemisia tabaci of Gerbera jameso-nii[J]. Jiangsu Journal of Agricultural Sciences,36(6):1398-1402.]
Akutse K S,Khamis F M,Ambele F C,Kimemia J W,Ekesi S,Subramanian S. 2020. Combining insect pathogenic fungi and a pheromone trap for sustainable management of the fall armyworm,Spodoptera frugiperda(Lepidoptera:Noctuidae)[J]. Journal of Invertebrate Pathology,177: 107477. doi:10.1016/j.jip.2020.107477.
Altre J A,Vandenberg J D,Cantone F A. 1999. Pathogenicity of Paecilomyces fumosoroseus isolates to diamondback moth,Plutella xylostella: Correlation with spore size,germination speed,and attachment to cuticle[J]. Journal of Invertebrate Pathology,73(3): 332-338. doi:10.1006/jipa. 1999.4844.
Amóra S S A,Bevilaqua C M L,Carneiro-Feijo F M,de Macedo Assuncao Pereira R H,Dutra Alves N,de Moráis Freiré F A,Kamimura M T,de Oliveira D M,Luna-Alves Lima E ?,Gadelha Rocha M F. 2010. The effects of the fungus Metarhizium anisopliae var. acridum on different stages of Lutzomya longipalpis(Diptera: Psychodidae)[J]. Acta Tropica,113(3):214-220. doi:10.1016/j.actatropica.2009.10.018.
Arthurs S,Dara S K. 2019. Microbial biopesticides for invertebrate pests and their markets in the United States[J]. Journal of Invertebrate Pathology,165: 13-21. doi:10.1016/j.jip. 2018.01.008.
Asensio L,Lopez-Llorca L V,Lopez-Jimenez J A. 2005. Use of light,scanning electron microscopy and bioassays to evaluate parasitism by entomopathogenic fungi of the red scale insect of palms (Phoenicococcus marlatti Ckll.,1899)[J]. Micron,36(2): 169-175. doi:10.1016/j.micron.2004. 09.004.
Bernardo C C,Barreto L P,e Silva C D S,Luz C,Arruda W,Fernandes ? K. 2018. Conidia and blastospores of Metarhizium spp. and Beauveria bassiana s.l.:Their development during the infection process and virulence against the tick Rhipicephalus microplus[J]. Ticks and Tick-borne Diseases,9(5): 1334-1342. doi:10.1016/j.ttbdis.2018.06. 001.
Bouamama N,Vidal C,Fargues J. 2010. Effects of fluctuating moisture and temperature regimes on the persistence of quiescent conidia of Isaria fumosorosea[J]. Journal of Invertebrate Pathology,105(2): 139-144. doi:10.1016/j.jip. 2010.05.014.
Butt T M,Ibrathim L,Clark S J,Beckett A. 1995. The germination behaviour of Metarhizium anisopliae on the surface of aphid and flea beetle cuticles[J]. Mycological Research,99(8):945-950. doi:10.1016/s0953-7562(09)80754-5.
Casmuze A,Juárez M L,Socías M G,Murúa M G,Prieto S,Medina S,Willink E,Gastaminza G. 2010. Revisión de los hospederos del gusano cogollero del maíz,Spodoptera frugiperda(Lepidoptera:Noctuidae)[J]. Revista de la Sociedad Entomológica Argentina,69(3-4): 209-231.
Chouvenc T,Su N Y,Robert A. 2009. Cellular encapsulation in the Eastern subterranean termite,Reticulitermes flavipes (Isoptera),against infection by the entomopathogenic fungus Metarhizium anisopliae[J]. Journal of Invertebrate Pathology,101(3): 234-241. doi:10.1016/j.jip.2009. 05.008.
Corrêa B,da Silveira Duarte V,Silva D M,Mascarin G M,Júnior I D. 2020. Comparative analysis of blastospore production and virulence of Beauveria bassiana and Cordyceps fumosorosea against soybean pests[J]. BioControl,65(3): 323-337. doi:10.1007/s10526-020-09999-6.
de Faria M R,Wraight S P. 2007. Mycoinsecticides and mycoa-caricides: A comprehensive list with worldwide coverage and international classification of formulation types[J]. Biological Control,43(3): 237-256. doi:10.1016/j.bio-control.2007.08.001.
Duan Y L,Wu H,Ma Z Y,Yang L,Ma D Y. 2017. Scanning electron microscopy and histopathological observations of Beauveria bassiana infection of Colorado potato beetle larvae[J]. Microbial Pathogenesis,111: 435-439. doi:10.1016/j.micpath.2017.09.025.
Early R,Gonzalez-Moreno P,Murphy S T,Day R. 2018. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda,the fall armyworm[J]. NeoBiota,40: 25-50. doi:10.3897/neobiota.40.28165.
Fang W G,Pava-Ripoll M,Wang S B,St. Leger R. 2009. Protein kinase A regulates production of virulence determinants by the entomopathogenic fungus,Metarhizium ani-sopliae[J]. Fungal Genetics and Biology,46(3): 277-285. doi: 10.1016/j.fgb.2008.12.001.
Ghaffari S,Karimi J,Kamali S,Moghadam E M. 2017. Biocontrol of Planococcus citri (Hemiptera: Pseudococcidae) by Lecanicillium longisporum and Lecanicillium lecanii under laboratory and greenhouse conditions[J]. Journal of Asia-Pacific Entomology,20(2): 605-612. doi:10.1016/j.aspen.2017.03.019.
Gillespie J P,Bailey A M,Cobb B,Vilcinskas A. 2002. Fungi as elicitors of insect immune responses[J]. Archives of Insect Biochemistry and Physiology,44(2):49-68. doi:10. 1002/1520-6327(200006)44:2<49::aid-arch1>3.0.co;2-f.
González Y R,Taibo A D,Jiménez J A,Portal O. 2020. Endophytic establishment of Beauveria bassiana and Metarhizium anisopliae in maize plants and its effect against Spodoptera frugiperda(J. E. Smith)(Lepidoptera: Noctui-dae) larvae[J]. Egyptian Journal of Biological Pest Control,30(1): 1-6. doi:10.1186/s41938-020-00223-2.
Hajek A E,Eastburn C C. 2003. Attachment and germination of Entomophaga maimaiga conidia on host and non-host larval cuticle[J]. Journal of Invertebrate Pathology,82(1): 12-22. doi:10.1016/s0022-2011(02)00198-2.
Holland R J,Gunasekera T S,Williams K L,Nevalainen K M H. 2002. Ultrastructure and properties of Paecilomyces lilacinus spores[J]. Canadian Journal of Microbiology,48(10): 879-885. doi:10.1139/w02-083.
Hussain A,Rizwan-ul-Haq M,Al-Ayedh H,Al-Jabr A M. 2014. Mycoinsecticides: Potential and future perspective[J]. Recent Patents on Food,Nutrition & Agriculture,6(1): 45-53. doi:10.2174/2212798406666140613113905.
Ibrahim L,Butt T M,Jenkinson P. 2002. Effect of artificial culture media on germination,growth,virulence and surface properties of the entomopathogenic hyphomycete Metarhizium anisopliae[J]. Mycological Research,106(6): 705-715. doi:10.1017/s0953756202006044.
Khan S,Nadir S,Lihua G,Xu J C,Holmes K A,Dewen Q. 2016. Identification and characterization of an insect toxin protein,Bb70p,from the entomopathogenic fungus,Beauveria bassiana,using Galleria mellonella as a mo-del system[J]. Journal of Invertebrate Pathology,133: 87-94. doi:10.1016/j.jip.2015.11.010.
Khun K K,Ash G J,Stevens M M,Huwer R K,Wilson B A L. 2021. Transmission of Metarhizium anisopliae and Beauveria bassiana to adults of Kuschelorhynchus macadamiae(Coleoptera:Curculionidae) from infected adults and conidiated cadavers[J]. Scientific Reports,11(1): 2188. doi:10.1038/s41598-021-81647-0.
Latgé J P,Sampedro L,Brey P,Diaquin M. 1987. Aggressiveness of Conidiobolus obscurus against the pea aphid: Influence of cuticular extracts on ballistospore germination of aggressive and non-aggressive strains[J]. Microbiology,133(7): 1987-1997. doi:10.1099/00221287-133-7-1987.
Lei Y Y,Hussain A,Guan Z Y,Wang D S,Jaleel W,Lü L H,He Y R. 2021. Unraveling the mode of action of Cordyceps fumosorosea: Potential biocontrol agent against Plu-tella xylostella(Lepidoptera: Plutellidae)[J]. Insects,12(2): 179. doi: 10.3390/insects12020179.
Liu T X,Stansly P A. 2009. Effects of relative humidity on efficacy of BotaniGuardTM(Beauveria bassiana) on nym-phs of sweetpotato whitefly,Bemisia tabaci(Hemiptera: Aleyrodidae) on hibiscus in greenhouses[J]. Southwest Entomologist,34(2): 189-191. doi:10.3958/059.034.0209.
Liu W M,Xie Y P,Xue J L,Zhang Y F,Zhang X M. 2011. Ultrastructural and cytochemical characterization of brown soft scale Coccus hesperidum(Hemiptera:Coccidae) infected by the Lecanicillium lecanii (Ascomycota: Hypocreales)[J]. Micron,42(1):71-79. doi:10.1016/j.micron. 2010.07.011.
Liu Z,Lei Z R,Hua B Z,Wang H H,Liu T X. 2010. Germination behavior of Beauveria bassiana(Deuteromycotina: Hyphomycetes) on Bemisia tabaci(Hemiptera: Aleyrodidae) nymphs[J]. Journal of Entomological Science,45(4): 322-334. doi:10.18474/0749-8004-45.4.322.
Mauchline N,Hallet I,Hill G,Casonato S. 2011. Process of infection of armored scale insects (Diaspididae) by an entomopathogenic Cosmospora sp.[J]. Journal of Invertebrate Pathology,108(1): 46-51. doi:10.1016/j.jip.2011. 07.002.
Ment D,Gindin G,Rot A,Soroker V,Glazer I,Barel S,Samish M. 2010. Novel technique for quantifying adhesion of Metarhizium anisopliae conidia to the tick cuticle[J]. Applied and Environmental Microbiology,76(11): 3521-3528. doi:10.1128/AEM.02596-09.
Montezano D G,Specht A,Sosa-Gómez D R,Roque-Specht V F,Sousa-Silva J C. 2018. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas[J]. African Entomology,26(2):286-300. doi:10.4001/003. 026.0286.
Ortiz-Urquiza A,Keyhani N O. 2013. Action on the surface: Entomopathogenic fungi versus the insect cuticle[J]. Insects,4(3): 357-374. doi:10.3390/insects4030357.
Pekrul S,Grula E A. 1979. Mode of infection of the corn earworm(Heliothis zea) by Beauveria bassiana as revealed by scanning electron microscopy[J]. Journal of Invertebrate Pathology,34(3): 238-247. doi:10.1016/0022-2011(79)90069-7.
Rangel D E N,Alston D G,Roberts D W. 2008. Effects of physical and nutritional stress conditions during mycelial growth on conidial germination speed,adhesion to host cuticle,and virulence of Metarhizium anisopliae,an entomopathogenic fungus[J]. Mycological Research,112(11): 1355-1361. doi:10.1016/j.mycres.2008.04.011.
Russo M L,Jaber L R,Scorsetti A C,Vianna F,Cabello M N,Pelizza S A. 2020. Effect of entomopathogenic fungi introduced as corn endophytes on the development,reproduction,and food preference of the invasive fall armyworm Spodoptera frugiperda[J]. Journal of Pest Science. doi:10.1007/s10340-020-01302-x.
Safavi S. 2010. Isolation,identification and pathogenicity assessment of a new isolate of entomopathogenic fungus,Beauveria bassiana in Iran[J]. Journal of Plant Protection Research,50(2): 158-163. doi:10.2478/v10045-010-0027-z.
Safavi S A,Shah F A,Pakdel A K,Reza Rasoulian G,Bandani A R,Butt T M. 2007. Effect of nutrition on growth and virulence of the entomopathogenic fungus Beauveria bassiana[J]. FEMS Microbiology Letters,270(1): 116-123. doi:10.1111/j.1574-6968.2007.00666.x.
St. Leger R J,Goettel M,Roberts D W,Staples R C. 1991. Prepenetration events during infection of host cuticle by Metarhizium anisopliae[J]. Journal of Invertebrate Patho-logy,58(2):168-179. doi:10.1016/0022-2011(91)90061-t.
Staats,C C,Kmetzsch L,Lubeck I,Junges A,Vainstein M H,Schrank A. 2013. Metarhizium anisopliae chitinase CHIT30 is involved in heat-shock stress and contributes to virulence against Dysdercus peruvianus[J]. Fungal Biology,117(2): 137-144. doi:10.1016/j.funbio.2012.12.006.
Talaei-Hassanloui R,Kharazi-pakdel A,Goettel M S,Little S,Mozaffari J. 2007. Germination polarity of Beauveria bassiana conidia and its possible correlation with virulence[J]. Journal of Invertebrate Pathology,94(2): 102-107. doi:10.1016/j.jip.2006.09.009.
Zhang Y J,Liu X Z,Wang M. 2008. Cloning,expression,and characterization of two novel cuticle-degrading serine proteases from the entomopathogenic fungus Cordyceps sinensis[J]. Research in Microbiology,159(6): 462-469. doi:10.1016/j.resmic.2008.04.004.
Zimmermann G. 2008. The entomopathogenic fungi Isaria farinosa(formerly Paecilomyces farinosus) and the Isaria fumosorosea species complex(formerly Paecilomyces fumosoroseus): Biology,ecology and use in biological control[J]. Biocontrol Science and Technology,18(9): 865-901. doi:10.1080/09583150802471812.
(責任编辑 麻小燕)
