华西雨屏区不同林龄杉木人工林土壤酶活性的动态变化1)
2021-07-30涂程伟彭彩云柳苹玉李琪张亚李天玲肖玖金
涂程伟 彭彩云 柳苹玉 李琪 张亚 李天玲 肖玖金
(四川农业大学,成都,611130)
人工林是我国工业用材的主要来源,兼具生态效益,杉木(Cunninghamialanceolata)具有生长快,用途广等优点[1-2]。杉木林多年连栽,面临着杉木林土壤肥力和生物多样性降低的现象[3]。在所有的土壤营养循环过程中,都有土壤酶的直接或间接参与[4-5]。土壤酶是土壤中的一类特殊的具有催化作用的蛋白质,其活性与微生物养分需求和土壤养分供给息息相关,并影响着林木生长的整个过程[6-7]。森林土壤酶活性的影响因素是一个复杂的综合体系,许多学者开展了不同植被下土壤酶活性研究[8-12],认为不同植被类型下土壤酶活性特征不同。如酸雨区杉木林氧化还原类酶在不同林龄的土壤中差异显著,且与pH值显著相关[13];随杉木人工林林龄的增加,土壤许多理化性质的退化会导致酸性磷酸酶、过氧化氢酶、脲酶、蛋白酶活性均有所降低[14];杉木林根际土壤酶活性计量比与取样季节显著相关[15];降水量的减少使参与碳循环的酶活性提高,并且干湿季土壤酶活性变化的主要驱动因子不一致[16]。总的来说,不同林龄与季节拥有不同的温度、水分、微生物量、真菌细菌比、植被生长情况等条件,共同驱动土壤酶活性发生变化[17-21]。
华西雨屏区是川西气候独特的多雨地带[22],而对该地区不同林龄杉木林土壤酶活性季节动态未见报道,因此,研究该区不同林龄杉木林土壤酶活性特征,以及土壤酶活性的响应机制,以期为人工杉木林的经营调整与改造提供技术支持。
1 研究区概况
研究区位于眉山市洪雅县洪雅林场八面山工区,地处四川盆地西南边缘,海拔1 316 m,属中亚热带湿润气候,气候温和、雨量充沛,年降水量1 435.5 mm,土壤类型为山地黄壤,地带性植被为亚热带常绿阔叶林[23]。
2 研究方法
2.1 样地设置
在2008年、2001年、1993年营造的杉木纯林中,于2017年选择类似的地形地貌、海拔、母岩、土壤类型、坡度、坡位等分别为设置3个20 m×20 m重复固定样地,分别为样地1(幼龄林,林龄10 a)、样地2(中龄林,林龄17 a)、样地3(近熟林,林龄25 a)。样地的概况和主要理化性质见表1、2。

表1 样地概况

表2 土壤主要理化性质
2.2 土壤酶活性的测定
2018年1月(冬季)、4月(春季)、7月(夏季)、10月(秋季)在不同林龄各样地内随机设置“品”字形取样点,去除地表覆盖物,使用环刀取0~10 cm土样,每层重复3次,带回实验室阴干,土样磨细后过20目筛,放入试剂瓶封好备用。参照文献[25],分别测定纤维素水解酶(CBH)、β-葡萄糖苷酶(βG)、酸性磷酸酶(AP)、β-N-乙酰氨基葡萄糖苷酶(NAG)、多酚氧化酶(PPO)、过氧化物酶(POD)等土壤酶活性。
2.3 数据处理
用EXCEL对数据进行收集与整理,SPSS19.0进行数据分析。对不同林龄及季节的土壤理化性质、土壤酶活性进行单因素、双因素方差分析和最小显著差异法(LSD)检验,显著性水平设定为α=0.05。
3 结果与分析
3.1 林龄、季节以及林龄×季节交互作用对酶活性的影响
由表3可知,林龄、季节以及林龄×季节交互作用对纤维素水解酶(CBH)及β-N-乙酰氨基葡萄糖苷酶(NAG)活性影响均极显著(P<0.01);林龄对β-葡萄糖苷酶与酸性磷酸酶活性的影响达极显著(P<0.01),季节对β-葡萄糖苷酶(βG)和酸性磷酸酶(AP)活性的影响小于林龄的影响,也达显著水平(P<0.05);林龄对多酚氧化酶(PPO)的影响不显著,但季节以及林龄×季节交互作用对多酚氧化酶(PPO)活性的影响达极显著(P<0.01);林龄及季节均对过氧化物酶(POD)活性有显著影响(P<0.05),但林龄×季节交互作用对过氧化物酶(POD)活性的影响不显著。纤维素水解酶(CBH)、β-葡萄糖苷酶(βG)、β-N-乙酰氨基葡萄糖苷酶(NAG)、多酚氧化酶(PPO)等受双因素交互作用影响极显著(P<0.01),即说明不同林龄的酶活性随季节变化趋势有所不同。
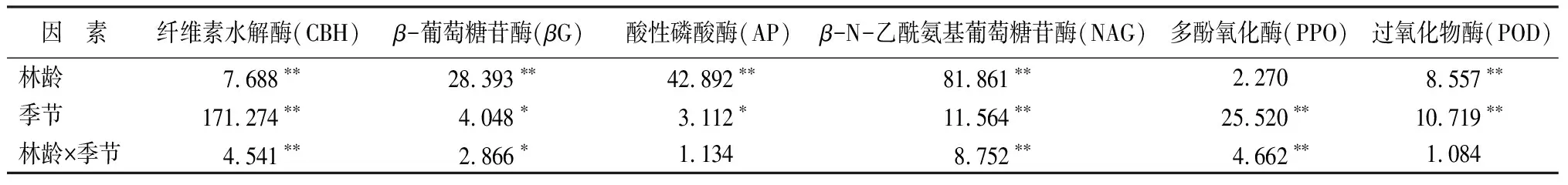
表3 酶活性的双因素方差分析(F值)
3.2 不同林龄不同季节的杉木林土壤酶活性
由表4可知,林龄10 a的杉木林土壤的纤维素水解酶(CBH)活性显著高于林龄17 a和25 a杉木林土壤(P<0.05),林龄25 a的杉木林土壤的纤维素水解酶(CBH)活性大于林龄17 a的杉木林土壤,但差异不显著(P>0.05),表明幼龄林土壤CBH酶活性较高;春季CBH酶活性均极显著高于其它季节(P<0.01);25 a杉木林土壤CBH酶活性在春季高于17 a杉木林土壤,但夏季却显著低于17 a杉木林土壤(P<0.05)。

表4 各林龄不同月份土壤CBH酶活性
由表5可知,10 a杉木林土壤的β-葡萄糖苷酶(βG)活性显著高于25 a杉木林土壤(P<0.05),25 a杉木林土壤的β-葡萄糖苷酶(βG)活性显著高于17 a杉木林土壤(P<0.05),βG酶活性随林龄增长呈现出先降低后升高的趋势;17 a杉木林土壤下,春季βG酶活性显著低于其它季节(P<0.05),其它林龄中βG酶活性的季节差异均不显著(P>0.05)。17 a杉木林土壤βG酶活性在春季均较低,且3种林龄下土壤酶活性由夏至秋的变化趋势均不同。

表5 各林龄不同月份土壤βG酶活性
由表6可知,10 a杉木林土壤的酸性磷酸酶(AP)活性极显著高于17 a杉木林土壤(P<0.01),17 a杉木林土壤的酸性磷酸酶(AP)活性显著高于25 a杉木林土壤(P<0.05),AP酶活性随林龄增长呈现出持续降低的趋势。10 a林龄中夏季土壤AP酶活性显著高于其它季节(P<0.05),17 a、25 a杉木林土壤AP酶活性除秋季较低外,季节性变化不大。

表6 各林龄不同月份土壤AP酶活性
由表7可知,10 a杉木林土壤的β-N-乙酰氨基葡萄糖苷酶(NAG)活性显著高于17 a和25 a杉木林土壤(P<0.05),17 a与25 a杉木林之间的土壤β-N-乙酰氨基葡萄糖苷酶(NAG)活性差异不显著(P>0.05),说明幼龄林下土壤NAG酶活性较高。10 a杉木林下夏季土壤NAG酶活性显著高于其它季(P<0.05),17 a杉木林土壤中夏季NAG酶活性亦显著高于春秋季(P<0.05),25 a杉木林土壤NAG酶活性在夏秋季显著低于冬春季(P<0.05),但17 a、25 a林龄下土壤NAG酶活性随季节变化幅度明显较10 a林龄下的变化幅度小。

表7 各林龄不同月份土壤NAG酶活性
由表8可知,各杉木林龄间的土壤多酚氧化酶(PPO)活性差异不显著(P>0.05)。各样地中冬季土壤多酚氧化酶(PPO)活性均显著高于春季(P<0.05),10 a、17 a杉木林夏秋季的土壤多酚氧化酶(PPO)活性差异不显著(P>0.05),25 a杉木林下土壤多酚氧化酶(PPO)活性在秋季显著高于春夏季(P<0.05),PPO酶活性由春至冬总体呈现上升趋势。

表8 各林龄不同月份土壤PPO酶活性
由表9可知,10 a与17 a杉木林土壤中过氧化物酶(POD)活性未有显著差异(P>0.05),但显著低于25 a杉木林土壤(P<0.05),说明杉木近熟林下POD酶活性较高。各样地夏季土壤POD酶活性均显著低于其它季节(P<0.05),POD酶活性由春至冬呈现先降低后升高的趋势。

表9 各林龄不同月份土壤POD酶活性
4 讨论
本试验中各土壤养分随林龄增长均有不同程度下降,与许多学者研究结果一致[24,26-29],说明研究地杉木在成熟前的迅速生长阶段,土壤养分消耗大于养分归还。林龄对土壤理化性质,水热状况和生物区系有着重要影响,同时影响土壤酶活性状况[21]。对比不同林龄发现,幼龄林土壤的4种水解酶类活性较高,表明其有机质分解迅速。耿玉清等[30]研究发现,随着林龄的增长,土壤酸性磷酸酶、转化酶和脲酶等多种酶活性皆下降。由于初期人工造林措施对土壤的改造,改善了土壤健康状况,促进了养分循环。但随着林木生长,土壤养分消耗,土壤紧实度增加,林内光照及土壤水热等条件变差,进一步导致微生物数量及酶活性显著降低[31-32]。而多酚氧化酶(PPO)、过氧化物酶(POD)的活性表现与水解酶有所不同,水解酶分解土壤中的活性有机质,而氧化酶分解土壤顽固的有机质[33-34],因此,不同酶的功能存在较大差异,这也部分支持了本研究的结果。多酚氧化酶(PPO)活性在林龄25 a的杉木林土壤显著高于林龄10 a的的杉木林土壤,且过氧化物酶(POD)活性在林龄25 a的杉木林土壤显著高于林龄10 a和17 a的杉木林土壤。多酚氧化酶(PPO)和过氧化物酶(POD)主要作用于木质素等难降解碳组分,其活性的变化说明林龄25 a杉木林土壤惰性碳库的分解较活跃,由于土壤惰性碳变化需要较长时间,这也是林龄10 a和17 a的林地之间未有显著差异的原因。在森林演替到后期才便于惰性有机碳的积累[35-36],印证了本研究的结果。
从不同季节看,纤维素水解酶(CBH)活性以春季显著高于其它季,说明其春季纤维素分解旺盛,由春至秋持续走低,这与陈光升[37]及宋学贵等[38]在华西雨屏区的研究结果一致;降水减少后参与碳循环的水解酶活性有所提高,这意味着降水量过大引起酶活性的降低,华西雨屏区夏秋季大量集中降水导致土壤持水量过高,水气热不协调,影响对纤维素的分解,加之试验地较为缺氮,集中降水对土壤表层氮磷的淋失[39],对纤维素水解酶(CBH)活性也有较大影响。而纤维素水解酶(CBH)活性在4月份(春季)最强,由于研究地气候非常湿润,尽管在4月份,水分仍未达限制水平,而此时土壤理化性质较适宜,植物根系和微生物活跃等因素,对纤维素水解酶(CBH)活性有较大的促进作用。β-葡萄糖苷酶(βG)主要降解富含纤维的细胞壁,释放其胞内营养,本试验中,β-葡萄糖苷酶(βG)多数情况下随季节变化并不显著,与鲍勇等[6]研究结果不完全一致,原因是研究区立地、林分及气候的差异。但βG酶在林龄17 a杉木土壤中,春季显著低于其它季,原因是土壤微环境不同,目前对βG酶相关研究较少,还很难完全解释βG酶在土壤中的变化规律。酸性磷酸酶(AP)和β-N-乙酰氨基葡萄糖苷酶(NAG)在林龄10 a杉木林土壤中,随季节变化趋势较林龄17 a和25 a的林地明显,林龄10 a的杉木林土壤内,酸性磷酸酶(AP)和β-N-乙酰氨基葡萄糖苷酶(NAG)酶活性均表现为夏季显著高于其它季,说明夏季土壤中有机氮磷分解较为活跃。研究区夏秋降水集中,土壤湿度增加,酸性磷酸(AP)酶以及N-乙酰氨基葡糖苷(NAG)酶活性有显著提高,同时夏季土壤动物及微生物量迅速增长,提高AP及NAG酶活性,而秋季酶活性降低则是土壤湿度过大,透气性差[40-42];而林龄17 a和25 a的杉木林土壤中,AP与NAG酶未发现明显规律性变化,原因是随林龄增长,森林生态系统自我反馈调节机制的成熟,林内环境趋于稳定。多酚氧化酶(PPO)、过氧化物酶(POD)活性总体表现为秋冬季大于春夏季,与相关研究结果一致[37,43],同时在与本试验区气候相近地区对杉木凋落物量的研究表明,杉木林凋落物量总体亦为秋冬季大于春夏季[44-45],因而PPO与POD酶活性的差异主要是植物残体增加,微生物、土壤动物活动加剧并向土壤中释放酶类,这对于促进秋季大量凋落物的木质素降解和腐殖化具有积极作用。
林龄及季节双因素交互作用对酶活性亦有显著影响,但不同林龄的土壤酶活性随季节变化的趋势有所区别。林龄25 a的杉木林土壤中,CBH酶活性由春季至夏季下降速度明显比林龄17 a杉木林迅速;AP酶与NAG酶在林龄10 a的杉木林土壤中,随季节变化而显著变化,在林龄17 a和25 a的杉木林土壤中变化幅度明显较小。进一步说明土壤酶受多种复杂因素的共同作用,很难完全解释其变化规律,还需进一步研究。
5 结论
研究区杉木人工林在成熟之前地力有不同程度的衰退,杉木林土壤中氮素较低。幼龄林土壤中,4种水解酶类活性均显著高于其它龄级,土壤条件总体较为优越,养分循环较为迅速;而两种氧化酶则相反,在近熟林才表现出显著提高。土壤酶活性季节动态方面,CBH酶活性春季显著高于其它季,βG酶多数情况下随季节变化并不显著,但总体春季有所降低;AP酶与NAG酶在杉木幼龄林土壤中变化显著,酶活性均为夏季显著高于其它季;PPO酶总体呈现出冬季显著高于其它季,POD酶活性亦为冬季最高,夏季显著低于其它季。不同林龄和季节对土壤酶活性存在明显交互作用的影响,说明不同林龄下立地条件的差异都有可能改变酶活性随季节变化的趋势。
