大兴安岭南段山杨死亡对其更新的影响1)
2021-07-30赵鹏武刘兵兵管立娟周梅舒洋
赵鹏武 刘兵兵 管立娟 周梅 舒洋
(内蒙古赛罕乌拉森林生态系统国家定位观测研究站(内蒙古农业大学),呼和浩特,010019)
次生林是森林生态系统重要的植被类型之一[1],也是半干旱区主要的森林类型,拥有丰富的物种组成,复杂的林分结构以及脆弱的环境特征。次生林通常呈现丰富的生物多样性,具有重要的生态功能且维持着区域陆地生态系统的平衡。近一个世纪以来,全球性增温导致气候剧烈变化,对陆地生态系统的恶劣影响不断显现。在气候变化背景下,半干旱区次生林发生大面积非正常死亡[2-5],以森林死亡和更新为标志的植被动态研究成为当前植物地理学研究的热点[6-7]。
内蒙古大兴安岭南段地处森林-草原交错区,也是“罕山次生林”的典型地段,边缘效应明显,生态系统的结构、功能及过程复杂。森林-草原交错区具有丰富的生物多样性和较高的生产力,但是抗干扰能力较差,较易受到气候变化的影响[1]。由于全球气候变化所引起的干旱、高温、生境破碎等,大兴安岭南段森林-草原交错区发生了具有较强斑块性的大面积山杨死亡现象。曾楠[1]对大兴安岭南段阔叶林乔木死亡动态研究表明,2015年乔木数量约为2012年调查乔木的1.66倍,2015年山杨株数较2012年增长了10%;干旱是导致大兴安岭南段树木死亡的主要因素之一,由于乔木死亡导致该地区出现大量林窗,林窗面积主要集中在50~170 m2,平均面积为134 m2[8-9]。然而,对于该地区山杨死亡后,森林能否更新,以及更新情况研究较少。因此,山杨林大面积死亡后能否快速天然更新,以及影响区域森林更新的因素值得研究。
本研究以大兴安岭南段山杨次生林为研究对象,研究不同山杨死亡梯度下林下乔木更新特征,建立不同山杨死亡梯度下林分因子与森林更新的相关关系,揭示山杨死亡后森林更新的影响机制,对区域森林生物多样性的维持、群落的稳定性保护以及次生林演替规律了解具有一定的参考价值。同时,为次生林恢复制定森林资源经营方案和培育保护措施,加强森林生态系统管理提供决策依据。
1 研究区概况
研究区位于内蒙古赤峰市巴林右旗赛罕乌拉国家级自然保护区内,地理坐标为43°59′~44°27′N,118°18′~118°55′E[10]。该区属于大兴安岭南部山地阿尔山支脉,地貌类型为中山山地,总体地势为东北高西南低,从东北向西南逐渐倾斜,平均海拔高度1 000 m,最高海拔1 997 m;研究区属温带半湿润温寒气候区,最高气温29 ℃,最低气温-32 ℃,年平均气温2 ℃,年均降水量400 mm,年蒸发量2 050 mm,湿润度为0.5~0.8[11]。土壤类型主要有山地黑土、灰色森林土、棕壤土以及暗粟钙土。主要森林类型有杨桦混交林、杨桦蒙古栎混交林、蒙古栎纯林等阔叶次生林,主要树种有山杨(Populusdavidiana)、白桦(Betulaplatyphylla)、蒙古栎(Quercusmongolica)、黑桦(Betuladahurica)、大果榆(Ulmusmacrocarpa),林下以虎榛子(Ostryopsisdavidiana)、兴安杜鹃(Rhododendrondahuricum)、照山白(Rhododendronmicranthum)和草类为主[12]。
2 研究方法
2.1 样地设置与调查
2019年8—10月,在研究区内以空间代时间的方法,选择山杨成年树基本存活的林分作为对照样地(L),设置3个重复(L1、L2、L3);选择成年山杨开始出现死亡林分(多以枯梢、将死或已死枯立山杨)作为山杨中度死亡样地(K),设置3个重复(K1、K2、K3);选择林中大部分山杨基本死亡,形成大量倒木作为山杨重度死亡样地(D),设置3个重复(D1、D2、D3)。3类样地共计9个标准样地,样地规格为20 m×20 m。
对样地内胸径≥1 cm的乔木进行每木检尺。在样地四个角及中心位置分别设置5 m×5 m的幼树幼苗调查样方5个,共计45(5×9)个样方,将胸径小于1 cm或树高小于1.3 m的植株定义为幼苗,将1 cm≤胸径≤5 cm的植株定义为幼树,记录幼苗及幼树枯梢和死亡情况。更新方式采用“刨土法”调查,分为实生苗和萌生苗(调查中幼苗与幼树全部以萌生为主)。同时,在幼苗样方中调查灌木物种数、盖度、多度、高度等,共计45(5×9)个样方。在幼苗、灌木样方内设置2 m×2 m的草本调查样方,调查物种数、盖度、多度、高度等,共计45(5×9)个样方。在各幼苗样地内随机选择3~4个点,进行枯落物厚度测量。林分郁闭度测量采用人工抬头望法[13]及FV2200冠层分析仪相结合进行测量。
幼苗幼树枯梢率=幼苗幼树枯梢株树/幼苗幼树总株数×100%;幼苗幼树死亡率=幼苗幼树死亡株树/幼苗幼树总株数×100%。
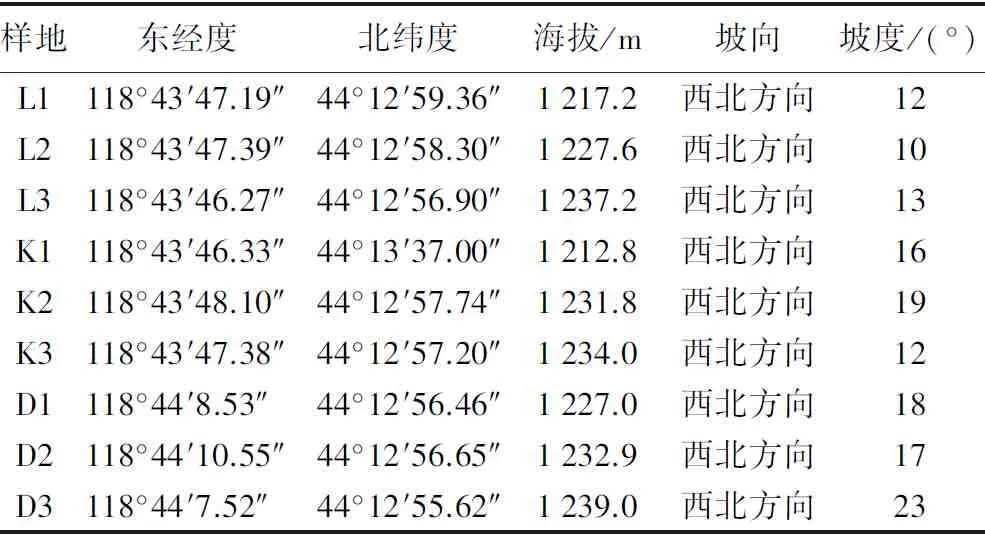
表1 试验样地基本信息
2.2 数据处理
运用Excel 2010进行数据初步整理,采用IBM SPSS 23进行单因素方差分析,运用R软件进行pearson相关分析。
3 结果与分析
3.1 山杨死亡对更新密度的影响
由表2可知,山杨不同死亡梯度下更新密度差异较大,山杨重度死亡样地的更新密度为7 308.33株/hm2,山杨中度死亡样地更新密度为2 866.67株/hm2,对照样地的更新密度425.00株/hm2,山杨重度死亡样地的更新密度分别是山杨中度死亡样地和对照样地的2.55倍和17.20倍。山杨重度死亡样地和山杨中度死亡样地的更新密度与对照样地差异性显著(P<0.05),山杨中度死亡样地的更新密度与山杨重度死亡样地差异性不显著,但更新密度是对照样地的6.75倍。不同死亡梯度下,山杨更新均以幼树为主。

表2 不同山杨死亡梯度下幼苗幼树密度特征
3.2 山杨死亡对更新幼苗(树)生长的影响
由表3可知,山杨不同死亡梯度下幼苗(树)平均树高和平均胸径(基径)具有不同的特征,山杨重度死亡样地的平均树高最大,平均树高为3.71 m,比山杨中度死亡样地和对照样地平均树高分别高0.86 m和2 m。山杨重度死亡样地与山杨中度死亡样地平均树高无显著差异,但均与对照样地差异显著(P<0.05)。山杨重度死亡样地中平均胸径最大,平均胸径为3.05 cm,比山杨中度死亡样地和对照样地平均胸径分别大0.34 cm和1.29 cm。山杨重度死亡样地与山杨中度死亡样地平均胸径无显著差异,但均与对照样地差异显著(P<0.05)。

表3 不同山杨死亡梯度下幼苗幼树生长特征
由表4可知,对不同山杨死亡梯度下幼苗(树)生长情况分布特征不同,随着胸径(或基径)的不断增大,山杨重度死亡样地、中度死亡样地和对照样地更新密度均呈现出先增后减的趋势,且在胸径为1~2 cm时增幅最大。在任何胸径范围内,更新密度均呈现出山杨重度死亡样地最大,对照样地最小。山杨重度死亡样地中,胸径在3~4 cm范围内更新密度最大,为1 866.67株/hm2。山杨中度死亡样地与对照样地均为胸径在1~2 cm范围内更新密度最大,分别为1 075.00株/hm2和166.00株/hm2。
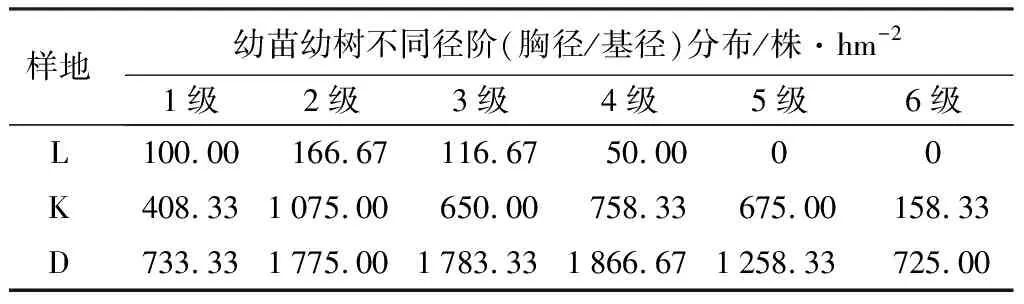
表4 不同山杨死亡梯度下幼苗幼树不同径阶的分布
由表5可知,不同死亡梯度下树高分布差异较大,山杨重度死亡样地中,随着树高的不断增高,更新密度逐渐增大,在树高级为8~9级时密度最大,更新密度为1 300株/hm2。山杨中度死亡样地树高分布特征呈现出先增大后减少的趋势,在树高级为2~4级时增幅最大,在3~4级时更新密度最大,更新密度为1 300株/hm2。树高在5级以上时,山杨重度死亡样地更新密度均高于山杨中度死亡样地。对照样地中在树高级为0~1级时,更新密度略高于山杨重度死亡样地和山杨中度死亡样地,在3~4级时更新密度最大,更新密度为233株/hm2。
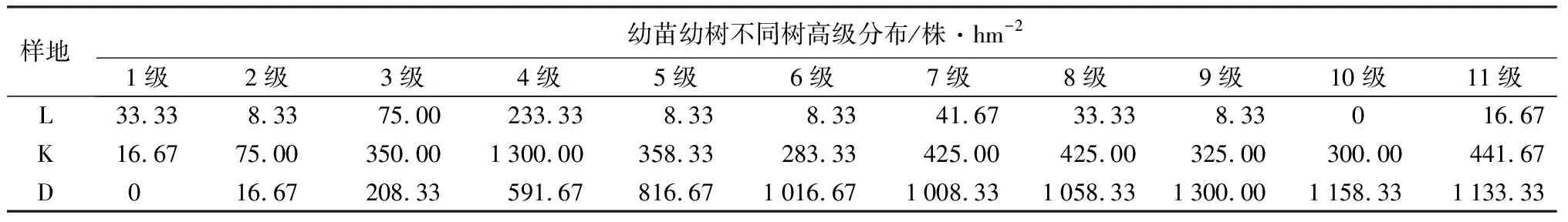
表5 不同山杨死亡梯度下幼苗幼树不同树高级高的分布
3.3 山杨死亡对幼苗幼树死亡率及枯梢率的影响
由表6可知,山杨不同死亡梯度下幼苗幼树枯梢率和死亡率不同,对照样地幼苗枯梢率较大,枯梢率为26.1%,未发现幼树枯梢现象;山杨重度死亡样地中有少量幼苗幼树枯梢现象;山杨中度死亡样地未发现幼苗幼树枯梢现象。山杨重度死亡样地幼苗死亡率较高,幼苗死亡率为63.9%,幼树死亡率略有升高;山杨中度死亡样地中幼苗死亡率为22.8%;对照样地未发现幼苗幼树死亡现象。

表6 不同死亡梯度下幼苗幼树死亡率及枯梢率 %
3.4 更新特征及其影响因素分析
由表7可知,幼苗幼树密度的更新密度、平均树高、平均胸径分别与郁闭度、林分密度呈显著负相关(P<0.05)。幼苗枯梢率分别与郁闭度、林分密度成显著正相关(P<0.05)。幼苗死亡率、幼树枯梢、死亡率分别与更新密度、郁闭度成显著负相关(P<0.05)。

表7 更新特征及其影响因素相关性
4 结论与讨论
大兴安岭南段山杨死亡后具有很强的更新能力,最大更新密度为7 967.33株/hm2,表明该地区山杨死亡后具有很强的繁殖下一代的能力,山杨更新密度与郁闭度和林分密度呈显著负相关(P<0.05)。在调查过程中发现,研究区所有山杨更新苗均为萌蘖更新。一方面,因为林下环境因素不适合种子的萌发,比如灌木和草本盖度、枯落物厚度等,由于受到枯落物、灌木和草本的阻隔影响,更新种子无法与土壤表面接触,因而缺乏种子发芽所需的水分和养分,极大的降低了种子的萌发率,使种子丧失更多成功定居的机会,从而影响实生更新质量[14];在立地环境因子中,林冠高度、林分郁闭度、凋落物厚度和林分密度是影响种子分布和幼苗数量的主要环境因子[15];草本盖度、凋落物层厚度和凋落叶中化感物质的抑制作用可能是导致红椎在针叶树人工林下更新不良的主要原因[16]。另一方面,因为种子不具有萌发能力,而在面临干旱胁迫下,山杨采取萌蘖更新(R策略)的高效繁殖策略延续下一代[17]。
山杨重度死亡样地更新密度最大,且拥有更多的幼树和幼苗,样地调查中也未发现实生幼苗和幼树,那么新增加的大量幼苗和幼树从何而来?原因是山杨林受干旱胁迫使树木生长衰退或地上部分的枯死[18],应对外界环境的变化,改变自己的营养生长为繁殖生长,通过地下根系形成大量无性系延续下一代[17]。
山杨死亡后更新幼苗具有形成幼树的能力,且出现生长速度加快的现象。在杨树和针叶树的混交林研究中指出,针叶树死亡后杨树表现出生长释放的现象[19];对死亡率与森林更新的研究发现,重度采伐(死亡)有利于森林更新,尤其是无性繁殖[20]。本研究中幼树密度显著高于幼苗密度,山杨中度死亡样地中,在树高级2~4级时,更新密度增幅最大,在3~4级时更新密度最大,更新密度为1 300株/hm2,同范围内的对照样地变化较小。这是因为在山杨开始死亡后,林分内可利用光、温等环境因子得到释放,促使更新幼苗开始快速生长。当幼苗达到一定密度后,由于幼苗与幼树之间又存在密度制约效应,其生长速度开始下降[21]。山杨重度死亡样地中,更新密度随着树高的不断增大而增大,在8~9级时更新密度达到最大后出现下降,这是因为山杨全部死亡后,更新的幼苗幼树生长再次得到释放,进而越来越多幼苗长成幼树,但是过多的幼树最终还是超过了环境容纳量,开始出现增长缓慢的现象,刘兵兵等[22]对大兴安岭南段杨桦次生林林下更新特征的研究发现,一定开阔度的林窗能够显著地促进林下物种更新与生长。本文相关分析也指出,幼苗(树)平均树高和平均胸径与郁闭度和林分密度呈显著负相关(P<0.05)。山杨幼苗(树)胸径(或基径)呈现山杨重度死亡样地最大,对照样地最小,充分说明了环境释放可促进幼苗生长释放,促进生长速度加快。
山杨重度死亡样地和山杨中度死亡样地中幼苗死亡率较高,分别为63.89%和22.76%,幼苗死亡率与更新密度(幼树)呈显著正相关(P<0.05)。这是因为后更新的幼苗竞争能力较弱,然而相对较稳定且密度高的幼树具有较强的竞争力。幼树死亡率整体较低,这也是幼树能够逐渐长成林的关键指标。但是,从山杨中度死亡样地到山杨重度死亡样地的过渡阶段内,幼树死亡率有上升的趋势。这是因为树木的不断生长,光、热等环境资源需求量不断增大,当达到一定限度时,便开始出现种内自疏现象[23]。因此,可以推断随着幼树的不断生长,幼树的死亡率逐渐升高,当达到合理的环境空间后,死亡率便逐渐降低,直至达到该种群生态位稳定的林分密度阈值。
随着山杨林不同梯度的生长衰退和生态过程对森林更新的影响,大兴安岭南段半干旱区山杨林受到干旱胁迫后,将立刻停止地上部分的营养生长,刺激地下根系开始繁殖生长,形成大量无性系繁殖,这也是大兴安岭南段山杨在干旱背景拥有快速更新能力的主要因素。因此,探讨和揭示半干旱区森林在气候变化中的生长、死亡和更新动态,对森林更新机制的了解具有重要的意义。
