应用CRISPR/Cas9技术敲除SERPINF2基因对牛病毒性腹泻病毒复制的影响
2021-06-23付强陈俊贞郭妍婷姚刚冉多良史慧君
付强 陈俊贞 郭妍婷 姚刚 冉多良 史慧君
(新疆农业大学动物医学学院,乌鲁木齐 830052)
牛病毒性腹泻病毒(Bovine viral diarrhea virus,BVDV)是引起牛病毒性腹泻/黏膜病(Bovine viral diarrhea/mucosal disease,BVD/MD)的主要病原[1],主要感染牛、羊和驼科动物,感染后常呈现体温升高、腹泻、黏膜充血及蹄叶炎等临床症状[2]。此外,BVDV还会引起母牛流产、死胎及牛群持续性感染,给牛群健康管理造成危害[3]。BVDV可通过血液、排泄物等途径水平传播,感染BVDV的牛发病率高、病死率低;还可以通过怀孕母牛垂直传播,感染孕牛产下的犊牛终身带毒,持续排毒,是重要传染源,同时也是生物制品、冻精、血清等主要污染源,BVDV是反刍动物最严重的病原体之一,会对养牛业造成严重的经济损失,危害极大[4]。探明该病毒胞内复制的关键因子途径将为BVD的诊断和防控提供理论依据[5]。
α2抗纤溶酶(serpin peptidase inhibitor clade F member 2,SERPINF2)是编码丝氨酸蛋白抑制剂SERPINF2家族的一员[6]。这个分子在1969年由Aoki等[7]发现,其主要的功能是能够有效抑制纤溶过程,1976年Moroi和Aoki[8]从人血浆中成功分离并鉴定了这个纤溶抑制剂。因为它能够迅速抑制纤溶酶活性并且免疫电泳迁移速度与α2球蛋白相当,因此命名为 α2-plasmin inhibitor[9]。
本课题组前期使用BVDV TC株感染Cajal间质细胞(interstitial cells of Cajal,ICC)建立转录组差异表达谱,发现SERPINF2基因表达量显著性上调[10],该基因是否是BVDV胞内复制的关键因子,对BVDV的胞内复制有何影响尚未见研究。本研究应用CRISPR/Cas9基因编辑技术敲除SERPINF2基因,研究SERPINF2基因对BVDV复制的影响。
1 材料与方法
1.1 材料
1.1.1 病毒、质粒及细胞 BVDV新疆分离株TC由新疆农业大学传染病实验室冉多良教授惠赠[11];pSPAX2(12260)、pMD2.G(12259)、lentiCRISPR v2(53961)质粒购自美国Addgene;MDBK细胞和HEK-293T细胞购自中国科学院典型培养物保藏委员会细胞库;大肠杆菌Stbl3感受态细胞购自北京博迈德基因技术有限公司。
1.1.2 试剂和仪器 聚乙烯亚胺(polyethylenimine,PEI)购 自 美 国 Polysciences公 司;TRIzol购 自Ambion公司;BsmB I、和T4 DNA Ligase购自New England Biolabs公 司;聚 凝 胺(TR-1003)购 自Sigma公司 ;Endofree Plasmid Miniprep Kit(PD1212)购自BIOMIGA公司;高糖DMEM、胎牛血清FBS、山羊血清和0.25% Trypsin-EDTA购自BI公司;PrimeScriptTMRT Master Mix(Perfect Real Time)试剂盒(RR036A)购自TaKaRa公司;嘌呤霉素购自上海碧云天生物技术有限公司;Marker Ⅰ(MD101)、PCR MIX、D2000 DNA Marker(MD114)、SuperReal PreMix Plus(SYBR Green)、细胞基因组DNA提取试剂盒、普通琼脂糖凝胶DNA回收试剂盒(DP210)购自天根生化科技(北京)有限公司;常规的化学试剂购自上海生工生物工程股份有限公司;ABI 7500 Fast Real-Time PCR System购自美国ABI公司;倒置荧光显微镜(Nikon TE2000U)购自Nikon公司。
1.2 方法
1.2.1 sgRNA序列设计、合成 根据GenBank数据库中SERPINF2(登录号:282522)基因序列用Benchling(https://benchling.com)设计,由苏州金唯智生物科技有限公司合成,序列见表1。分别取10 μL sgRNA正、反单链(100 μmol/L),混匀后置于37℃孵育30 min,95℃孵育5 min,以5℃/min速度降温至25℃,置于-20℃保存。

表1 sgRNA序列Table 1 SgRNA sequences
1.2.2 lentiCRISPR v2-SERPINF2-sgRNA质 粒 构建 用BsmB I限制性内切酶酶切lentiCRISPR v2质粒后,用1% DNA凝胶电泳检测结果。该质粒酶切后会得到两条不同大小的DNA片段,将12 988 bp大小的目的条带用TIANGEN普通琼脂糖凝胶DNA回收试剂盒回收。将回收产物和sgRNA双链用T4 DNA连接酶4℃过夜连接,连接产物转化至Stbl3感受态细胞中,涂板后培养过夜,次日挑取单克隆菌落,PCR和测序鉴定。
1.2.3 SERPINF2 KO细胞建立 六孔细胞培养板每孔用500 μL的0.1%明胶包被30 min后弃明胶,接种5×105个HEK-293T细胞,置于细胞培养箱中培养。待细胞密度达到80%时,分别取3 μg lentiCRISPR v2-SERPINF2-sgRNA 质粒、3 μg pSPAX2质粒和 1.5 μg PMD2.G 质粒与 38.5 μL PEI(1 mg/mL)混匀,转移至细胞培养液中。孵育3 h后更换正常细胞培养液,置于细胞培养箱中继续培养48 h,收集培养液1 500 r/min离心5 min,收集上清液悬浮感染MDBK细胞,同时加入聚凝胺(8 μg/mL)增加感染效率。感染24 h后更换新鲜细胞培养液,继续培养48 h后加入嘌呤霉素(4 μg/mL)筛选4代获得阳性细胞。
1.2.4 SERPINF2敲除效率检测 收集SERPINF2 KO细胞,使用基因组DNA提取试剂盒提取细胞基因组DNA,通过PCR扩增目的片段,将目的片段回收后连接到pMD18-simple T载体上,转化至大肠杆菌Stbl3感受态细胞中,单克隆菌落经PCR后,随机挑选30个单克隆菌落送至苏州金唯智生物科技有限公司进行测序。PCR扩增所用引物序列为F :5′-AGTACAGGGGCTTTGAAGGAAATCT-3′,R :5′-TCCAGTCCAAAAGAGACACTGGAAC-3′。
1.2.5 qRT-PCR检测SERPINF2基因和BVDV RNA水平 将MDBK细胞按照5×105个细胞/孔接种至6孔细胞培养板中,待细胞汇合度达到70%时,加入1 000 TCID50BVDV感染细胞,分别于感染后36 h、48 h时,收集细胞和培养液并加入TRIzol裂解液裂解,提取总RNA,反转录1 μg总RNA成cDNA;以cDNA为模板,使用qRT-PCR检测SERPINF2 mRNA水平[10]。
分别接种等量的SERPINF2 KO细胞和对照组Scramble细胞并感染1 000 TCID50的BVDV,分别于感染后0、12、24、36、48 h时,收集细胞和培养液并提取总RNA,反转录1 μg总RNA成cDNA,同上采用 qRT-PCR 检测 BVDV 5′UTR RNA 水平[12]。
1.2.6 免疫荧光染色检测BVDV双链RNA含量 将SERPINF2 KO和Scramble细胞按照5×104个/孔的密度接种至铺有爬片的24孔细胞培养板中,待细胞密度达到80%时,用1 000 TCID50BVDV分别感染12、24、36、48 h后弃掉培养液,加200 μL 4%多聚甲醛室温下固定15 min,加入200 μL封闭液(1%山羊血清+3% BSA+1% Triton X-100,溶于PBS)室温下孵育1 h,加入J2 anti-dsRNA IgG2a单克隆抗体(1∶1 200稀释)4℃孵育过夜。加入Cy3标记山羊抗鼠IgG(H+L)二抗(1∶200稀释)避光室温孵育1 h,封片在激光共聚焦显微镜下观察荧光情况。
1.2.7 BVDV CPE情况和病毒滴度检测 将SERPINF2 KO细胞和对照Scramble细胞按照1×106个细胞/孔的密度接种至6孔细胞培养板中培养;待细胞贴壁后,加入1 000 TCID50BVDV感染细胞,分别于感染后12、24、36和48 h时,使用倒置显微镜观察CPE情况;并将细胞和培养液反复冻融3次,收集病毒悬液至1.5 mL离心管,1 000 r/min离心10 min收集上清液,使用Reed-Muench方法测定各上清液病毒滴度[12]。
1.2.8 统计分析 使用SPSS 17.0 for Windows(SPSS Inc.Chicago,IL,USA)进行统计学分析。*表示通过t检验确定的统计学显著性差异(*P <0.05;**P<0.01)。
2 结果
2.1 lentiCRISPR v2质粒酶切菌液PCR鉴定结果
使用限制性内切酶BsmB I酶切lentiCRISPR v2载体,用1%琼脂糖凝胶电泳检测到两条清晰的条带,其中小片段大小约2 000 bp,与预期结果一致(图1)。
2.2 SERPINF2基因敲除效率检测
应用CRISPR/Cas9技术敲除SERPINF2基因,收集部分细胞,通过PCR扩增目的片段,克隆至pMD18-simple T载体中,随机送样测序检测SERPINF2基因敲除效率。结果显示,测序成功的27株阳性克隆中有19株基因序列发生缺失(-)和插入(+)等移码突变,共有70.37%(19/27)样本的基因发生改变(图2),表明MDBK细胞的SERPINF2基因敲除率为70.37%。

图1 lentiCRISPR v2质粒酶切鉴定结果Fig.1 Identification of lentiCRISPR V2 plasmid by enzyme digestion

WT:野生型;-:碱基缺失;+:碱基插入;数字:碱基个数;(数字):缺失或插入等突变的克隆数WT:wild type, -:base deletion, +:base insertion, Number:base number, (Number):Clone number of mutation such as deletion or insertion
2.3 qRT-PCR检测SERPINF2 mRNA转录水平
为检测BVDV感染SERPINF2表达的影响,对BVDV感染MDBK细胞36和48 h后,使用qRTPCR检测SERPINF2 mRNA转录水平。结果发现,与未感染BVDV相比,BVDV感染36 h和48 h时SERPINF2 mRNA转录水平分别提高约3.19和12.46倍(图3),表明BVDV感染MDBK细胞显著性提高SERPINF2 mRNA转录水平。
2.4 qRT-PCR检测BVDV感染SERPINF2 KO细胞后BVDV RNA水平
BVDV感染SERPINF2 KO细胞和对照组Scramble细胞12、24、36、48 h后,提取总RNA,qRT-PCR检测BVDV 5′UTR RNA水平。结果显示与对照组Scramble细胞相比,BVDV感染SERPINF2 KO细胞12 h后BVDV RNA水平显著降低(图4)。
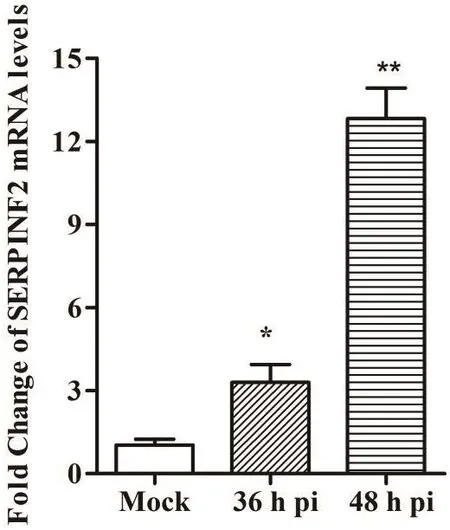
图3 qRT-PCR检测BVDV感染MDBK细胞后SERPINF2 mRNA转录水平Fig.3 Transcription levels of SERPINF2 mRNA in the MDBK cells infected by BVDV via qRT-PCR
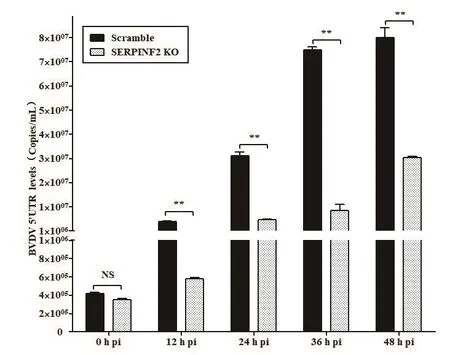
图4 qRT-PCR检测BVDV感染SERPINF2 KO细胞后BVDV RNA水平Fig.4 qRT-PCR detection of BVDV RNA level in the SERPINF2 KO cells infected with BVDV
2.5 免疫荧光染色检测BVDV感染SERPINF2 KO细胞后BVDV dsRNA含量
BVDV感染SERPINF2 KO细胞和对照组Scramble细胞12、24、36、48 h,用免疫荧光染色后观察细胞内BVDV sgRNA的分布情况,激光共聚焦显微镜下BVDV dsRNA呈红色荧光,结果可见SERPINF2 KO细胞中红色荧光显著少于对照组Scramble细胞(图5),表明BVDV dsRNA含量降低,BVDV的复制被阻止。

图5 免疫荧光染色检测BVDV感染SERPINF2 KO细胞后BVDV双链RNA含量结果Fig.5 Detection of BVDV dsRNA accumulation in the SERPINF2 KO cells infected with BVDV by immunofluorescence staining
2.6 BVDV感染SERPINF2 KO细胞后病毒滴度检测
BVDV感染SERPINF2 KO细胞和对照组Scramble细胞12、24、36、48 h,反复冻融后收集病毒悬液,使用Reed-Muench法测定病毒滴度变化。结果显示BVDV感染SERPINF2 KO细胞12 h后的病毒滴度与对照组Scramble细胞相比显著性降低(图6)。结果表明敲除SERPINF2基因能够显著性阻止BVDV复制。

图6 BVDV感染SERPINF2 KO细胞后病毒滴度检测结果Fig.6 Detection results of virus titer in the SERPINF2 KO cells infected with BVDV
2.7 BVDV感染SERPINF2 KO细胞后CPE检测结果
BVDV感染SERPINF2 KO细胞和对照Scramble细胞12、24、36、48 h后,在倒置显微镜下观察细胞CPE情况。结果显示BVDV感染SERPINF2 KO细胞24 h后CPE明显少于对照组,BVDV感染36 h后对照组Scramble细胞大量病变脱落,SERPINF2 KO细胞虽出现较多病变,但细胞脱落情况较少(图7)。表明敲除SERPINF2基因能够抑制BVDV感染造成的细胞CPE。

图7 BVDV感染SERPINF2 KO细胞后致细胞病变效应检测结果Fig.7 Detection results of cytopathic effect in the SERPINF2 KO cells infected with BVDV
3 讨论
利用CRISPR/Cas9基因编辑技术敲除靶基因,选择靶基因的识别序列(sgRNA识别序列)是关键。目标基因的识别序列一般是在前间区序列邻近基序(protospacer adjacent motif,PAM)三联核昔酸结构(NGG或者NAG,N代表任何碱基)前的20个左右的DNA碱基序列[13]。为了实现对SERPINF2基因的敲除,原则上可以将目标序列设计在SERPINF2基因启动子或外显子区域,进行敲除对SERPINF2基因目标序列进行切割并经HDR修复时引入突变。理论上可行的sgRNA识别片段不一定能最终实现基因的敲除,所以本实验尝试性设计了3个符合条件的SERPINF2基因的sgRNA识别序列,进而实现对SERPINF2基因的有效敲除。最终成功敲除MDBK细胞的SERPINF2基因,敲除效率为70.37%。为研究SERPINF2基因对BVDV的胞内复制的影响及作用机制奠定基础。
SERPINF2是SERPIN超家族的重要成员,其主要作用是通过抑制plasmin活性参与抗溶血,阻碍细胞因子生成、血管发生等生物学过程[14]。有病历研究发现,SERPINF2突变会造成胞内转运异常和功能障碍[15]。BVDV TC株感染ICC细胞转录组差异性分析发现,SERPINF2基因表达量显著性上调[10],感染MDBK细胞也得到同样的结果,考虑SERPINF2敲除会引起胞内转运异常和功能障碍,从而影响BVDV的胞内转录复制。
本研究发现BVDV感染MDBK细胞后SERPINF2基因表达量上调,与前期建立的BVDV感染宿主细胞建立的转录组差异表达谱相一致。利用CRISPR/Cas9基因编辑技术建立SERPINF2敲除的MDBK细胞SERPINF2 KO,并验证敲除SERPINF2基因能够显著性降低BVDV RNA水平、dsRNA积累,降低CPE情况和病毒滴度,表明SERPINF2敲除影响BVDV在宿主细胞中复制,而该基因通过何种途径影响BVDV的复制将在后续研究中深入探索。
4 结论
BVDV感染MDBK细胞SERPINF2基因表达量上调;利用CRISPR/Cas9基因编辑技术成功建立SERPINF2 KO细胞,敲除率为70.37%;SERPINF2基因敲除能够抑制BVDV在宿主细胞中的复制。