陆地棉与野生斯特提棉远缘杂种性状鉴定及遗传解析
2021-06-23赵祝跃申状状王一帆杨亚杰荣二花吴玉香
赵祝跃 申状状 王一帆 杨亚杰 荣二花 吴玉香
(山西农业大学农学院,太谷 030801)
棉花属双子叶植物锦葵科(Malvaceae)、棉属(Gossypium),是世界上最重要的经济作物之一,也是种植面积最广的纤维作物。最新提出的棉属分类系统为4个亚属,目前已经发现的有52个种(变种),9个组,其中5个为异源四倍体棉种(2n=4X=52)[1],分别是陆地棉(G.hirsutum,[AD]1)、海岛棉(G.barbadense,[AD]2)、达尔文氏棉(G.darwinii,[AD]5)、夏威夷棉(G.tomentosum,[AD]3)和黄褐棉(G.mustelinum,[AD]4),其基因组为AD组,其余46个为二倍体棉种(2n=2X=26),其基因组分别为A、B、C、D、E、F、G和K[2]。棉属广泛分布于澳洲、美洲、非洲和阿拉伯半岛等地区,经过了不同地域长期的自然选择和人工选择后,产生了许多的栽培棉种和众多的野生棉种[3]。陆地棉(G.hirsutum)为AD染色体组异源四倍体栽培棉,是目前中国乃至世界栽培范围最广的棉种。在部分其他地区,海岛棉、草棉和亚洲棉等也有种植。棉属栽培种和野生种是自然界丰富的种质资源,但栽培棉种缺乏部分野生种的优良特性。因此,将野生棉的优良性状导入栽培棉,从而创造出多抗优异新种质,对于棉花遗传育种有着重要意义。
远缘杂交就是不同种属间甚至亲缘关系更远的物种之间的杂交。棉花远缘杂交育种在一定程度上打破了物种间的界限,综合各棉种的优良特性到一个杂种中,从而创造出新的优异种质资源。目前,国内外的很多学者已经成功地将野生棉的部分优良基因导入到栽培棉中,选育出一批优良的陆地棉品种[4]。美国南卡罗莱那州Pee Dee农业试验站将亚洲棉和瑟伯氏棉(G.thurberi)远缘杂交,进行基因重组,培育成纤维品质优异的PD系统品种[5]。中国农业科学院棉花研究所通过用亚洲棉(G.arboreum)、斯特提棉(G.sturtianum)和陆地棉(G.hirsutum)杂交育成了中G2、中G3等兼抗枯、抗黄萎病新品系[6]。肖松华等[7](2011-2012)通过对陆地棉与异常棉(G.anomalum)、辣根棉(G.armourianum)、旱地棉(G.aridum)、雷蒙德氏棉(G.raimondii)、黄褐棉(G.mustelinum)等野生棉的远缘杂交和回交后育成的65份陆地棉为背景远缘种质系然后进行筛选,获得了抗黄萎病的棉花新种质,为我国抗病新品种的选育提供了材料。近年来,我国棉花种质资源的数量位居世界第四,但棉花资源的遗传多样性并不高。因此,棉花远缘杂交是创造异源新种质最有效的办法,也是棉花新品种选育的主要途径。此外,还要积极和棉花种质资源的富集国开展更多的交流[8]。
陆地棉属于AD染色体组异源四倍体栽培棉,原产于中美洲墨西哥的高地及加勒比海地区,亦称高原棉,最早因在美洲大陆种植而闻名,具有产量高、适应性广、结铃性强、皮棉产量高等优良品质[9]。野生斯特提棉(G.sturtianum,C组,2n=2X=26)原产于澳大利亚的中部和南部,具有许多优良性状如抗寒、抗萎病、生长势强、纤维优质、早熟等,尤其是植株有腺体种子无腺体的特性是重要的农艺性状,是现有栽培棉种所不具备的。斯特提棉的棉籽仁中没有色素腺体,待幼苗时色素腺体出现在植株的茎叶、花蕾等器官上[10]。通过远缘杂交以期将斯特提棉有益特性转入栽培种陆地棉中,创造综合父母本优良性状的新种质。
本研究以申状状等[11]前期合成的陆地棉与野生斯特提棉远缘杂种为材料,与其亲本陆地棉和斯特提棉进行初步形态性状比较,然后对杂种进行细胞遗传学研究,旨在观察不育杂种F1减数分裂异常行为,分析出现异常现象的原因,并对杂种进行SSR分子标记鉴定,以期为杂种提供更加直观的分子遗传学证据,进一步为棉花遗传育种和种质创新提供理论依据。
1 材料与方法
1.1 材料
母本栽培种陆地棉品种中棉所16,父本野生种斯特提棉(表1),以及本实验室前期合成的陆地棉和斯特提棉的远缘杂种F1。将材料同时种植于温室的花盆中进行后期观察和鉴定。
表1 供试材料及其地理分布Table 1 Materials and their geographical distribution
1.2 方法
1.2.1 形态学鉴定 以当年种成的植株为材料,取完全成熟的同龄器官和组织,分别记录植株、叶片和花等形态性状,并和父母本做比较,从形态学上分析鉴定杂种的真实性。从父本、母本和杂种植株的相同位置取完全展开的叶片进行观察并测量分析,包括形状、大小、短柔毛和腺体颜色等性状。在开花期,将杂种植株的完全展开花的形态特征与其亲本进行比较分析,包括小苞片、花萼、花冠、柱头、雄蕊、花粉和雌蕊等,记录并进行相关性状描述。
1.2.2 细胞遗传学鉴定
1.2.2.1 取材与固定 从杂种植株选取不同发育阶段的花蕾,并立即固定在新制备的Carnoy液(乙醇∶乙酸= 3∶1,V / V)中,24 h后将固定的花蕾转移到70%乙醇中并在4℃下储存,用于细胞遗传学观察。
1.2.2.2 制片 首先,取成熟植株上的花蕾,在干净滤纸上用解剖针和镊子轻轻的剥开,取花蕾中大约5-7颗花药置于蒸馏水清洗过的载玻片上,然后滴加少量改良卡宝品红[12](原液A:3 g碱性品红+100 mL 70%乙醇,原液B:10 mL原液A+90 mL 5%苯酚水溶液,充分混匀,置37℃温箱中2-4 h,原液C:55 mL原液B+6 mL冰乙酸+6 mL甲醛,充分混匀,染色液:10-20 mL原液C+80 mL 45%乙酸+1 g山梨醇)溶液,用镊子充分挤压花药将花粉母细胞挤出,去除可见杂质,盖上盖玻片,最后将滤纸放在盖破片上,用大拇指在其正上方垂直按压玻片吸去多余的染液。
1.2.2.3 镜检 使用具有自动相机的Olympus BX60显微镜从新制备的载玻片中观察并照相,选取至少500个花粉母细胞进行减数分裂行为观察,记录并做统计分析。
1.2.3 SSR分子标记鉴定 使用CTAB法提取来自远缘杂种植株的基因组DNA及其亲本种质DNA,DNA 扩增的总体积为 20 μL,含有 2 μL10× 缓冲液,1.6 μL MgCl2(25 mmol/L),0.2 μL dNTPs(10 mmol/L),6 μL 模板 DNA(50 ng /μL),2 μL 正向和反向引物(2.5 μmol/L),0.2 μL Taq 聚合酶(5 U/μL)和8 μL ddH2O。筛选出5对多态性简单序列重复(SSR)引物进行扩增,94℃预变性3 min,94℃变性50 s,58℃复性50 s,72℃延伸2 min,35个循环,72℃延伸10 min。通过PAGE分离扩增的片段(20 μL)并通过银染分离照相,对扩增清晰的条带进行分析,参考申状状等[13]进行银染检测。SSR引物序列来自http://www.cottonmarker.org,由北京赛百盛基因技术有限公司合成。
1.2.4 数据统计与分析 在胶片观察灯下读带,按照条带清晰,重复性好的标准进行了筛选,每次实验均重复2次,只有2次重复均相同的扩增带才被用于统计分析。记录杂种和亲本的相同和差异条带。
2 结果
2.1 远缘杂种F1的形态性状分析
母本陆地棉植株形态特征为:主茎干直立粗壮,为深绿色,下部枝叶较少或没有,嫩枝或嫩叶被有稀或密的茸毛,也有无茸毛的,花瓣为乳白色。父本斯特提棉植株枝条纤细,为青绿色,叶片比较小,呈梨形,叶脉弧形。杂种植株具有较强的生长势,株高高于双亲,主茎干直立且硬,翠绿色,枝叶繁茂,植株色素腺体多,茎毛短(图1)。

图1 亲本与杂种的植株比较Fig.1 Plant comparison between parents and hybrid
对杂种及其亲本的叶片进行分析比较发现(图2),母本陆地棉的叶片为掌状、三裂片、裂片呈宽三角形、锐尖、基部不收缩、颜色为深绿;父本斯特提棉的叶片比较小、呈梨形、叶脉弧形、颜色为翠绿色;杂种F1的叶片大于双亲,叶色与父本接近为翠绿色,叶面积比父本大比母本小,掌状,三裂,叶裂刻较深,叶片整体形态趋于母本陆地棉,叶脉深浅介于双亲之间,叶片茸毛较短。

图2 亲本与杂种的叶片比较(标尺:20 mm)Fig.2 Leaf comparison between parents and hybrid(Ruler: 20 mm)
对杂种及其亲本的花进行比较分析发现(图3),母本陆地棉花瓣为乳白色,基部无红斑;父本斯特提棉花冠较小,花瓣颜色为淡紫色,花瓣基部有深紫色花斑,花丝深紫色;杂种F1的花瓣颜色为粉红色,基部花斑与父本相近,颜色为深红色且基斑较大,花药为乳白色,雌蕊柱头乳白色像母本,柱头长度比母本长但比父本短。

图3 亲本与杂种的花朵比较Fig.3 Flower comparison between parents and hybrid
2.2 远缘杂种细胞遗传学鉴定
为了研究杂种的细胞遗传特性,本实验对杂种的花粉母细胞(PMC)减数分裂行为进行观察。结果表明:杂种的减数分裂出现许多异常行为,观察分别发现PMC的异常二分体(图4-A)、PMC的三分体(图4-B)、PMC的异常四分体(图4-C)、PMC的部分正常四分体(图4-D)和PMC的多分体(图4-E-H)等,以至于最后形成许多异常的花粉粒(图5),分析其原因是由于杂种减数分裂后期I的非同步和不均等染色体分离造成的。通过统计观察,在500个多分体中,分别有120个二分体(24.00%)、52个三分体(10.40%)、203个四分体(40.60%)和125个其他多分体(25.00%)(表2)。其中在观察到的203个四分体中,148个是正常四分体(占72.91%),55个是异常四分体(占27.09%)。异常的三分体、异常四分体、异常多分体都不能形成正常的花粉粒,因此本研究也同时观察到各种异常花粉粒,包括发育不良的花粉粒、畸形的花粉粒和花粉粒破裂等(图5)。在500个选定观察的花粉粒中,有380个花粉粒(占76.00%)是正常的,120个(占24.00%)为异常花粉粒,观察发现大部分异常的花粉粒都属于未发育完全的类型。
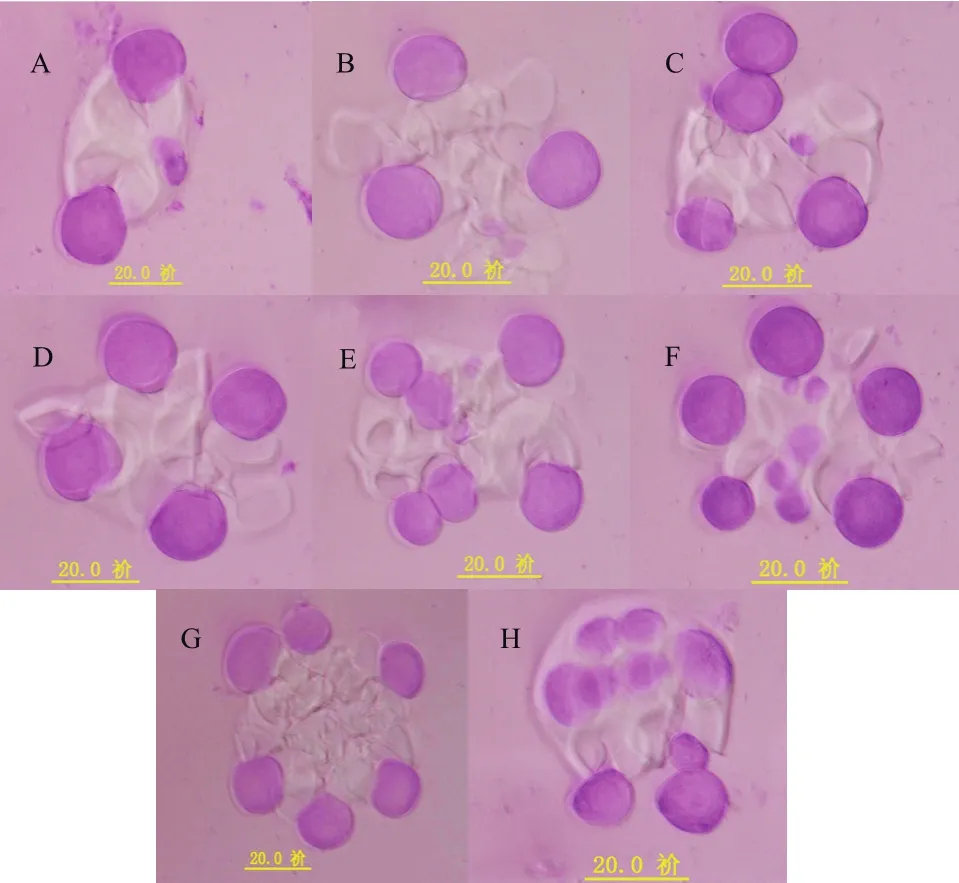
图4 杂种的正常和异常减数分裂行为(标尺:20 μm)Fig.4 Normal and abnormal meiosis behavior of G.hirsutum×G.sturtianum F1 hybrid (Ruler: 20 μm)
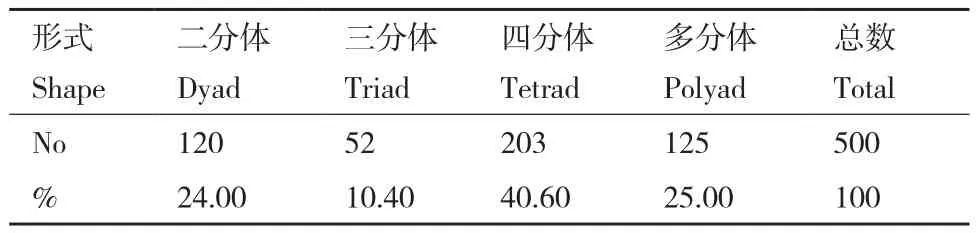
表2 杂种减数第二次分裂末期的多分体数量及比例Table 2 Number of multispores in telophase II of ADC hybrid meiosis
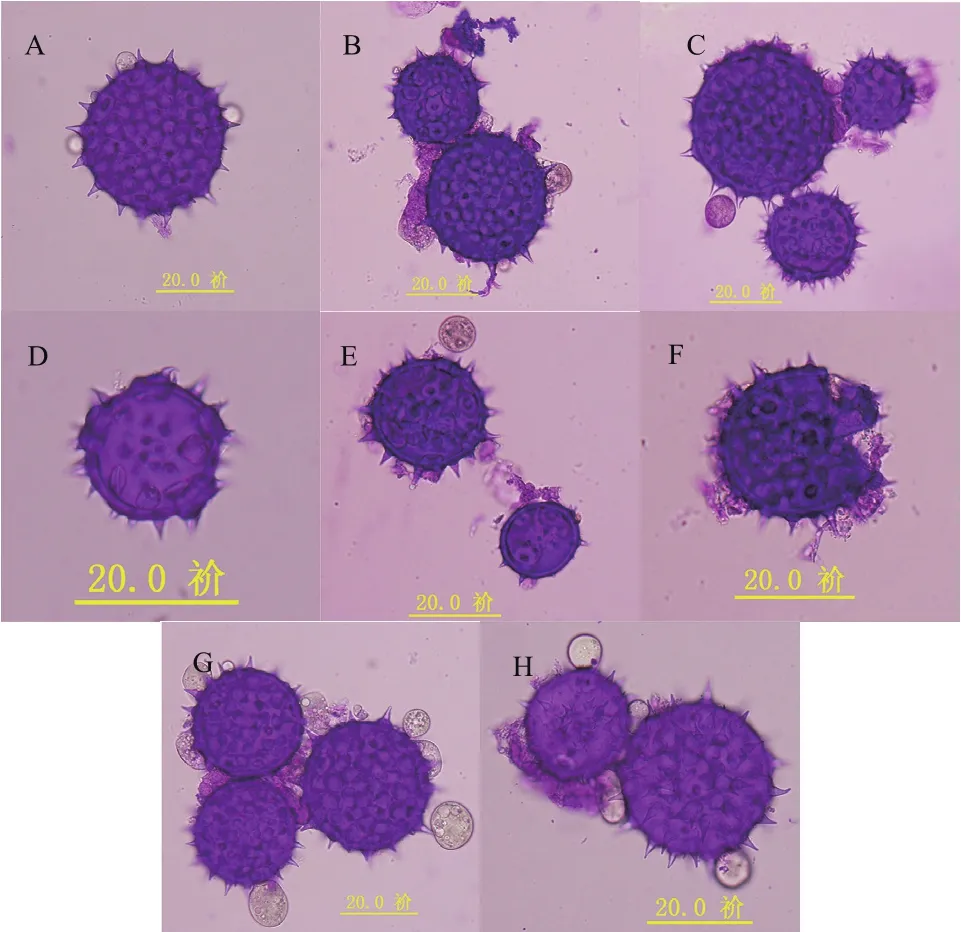
图5 杂种减数分裂后的正常和异常花粉粒(标尺:20 μm)Fig.5 Various normal and abnormal pollen grains after meiosis of F1 hybrid (Ruler: 20 μm)
2.3 远缘杂种SSR分子标记鉴定
通过对陆地棉、斯特提棉和其杂种在Marker2000下的SSR引物筛选和多态性分析,最后从中挑选出5对在三者间存在多态性的引物(图6)。为了更加深入的了解杂种和亲本之间在分子水平上的差异,使用5对多态性SSR引物进行了杂种和两个亲本的SSR分析。结果显示,在陆地棉和斯特提棉的杂种中存在26条清晰的多态性带。其中,在引物NAU1085中有6条清晰的条带,2条与母本陆地棉相同,另外4条则为父母本都没有的杂种新扩增出的特异性条带;在引物NAU1209中,观察到扩增出3条清晰的条带,其中一条与母本相同,一条与父本相同,也就是双亲的互补条带,剩余1条是杂种的特异性条带;在引物NAU3405中有5条清晰的条带,其中1条只与母本陆地棉相同,2条双亲的互补条带,另外2条是杂种扩增出的特异性条带;在引物TMB1125中,共找到7条清晰的条带,其中有3条只与母本陆地棉相同,2条同时与父母本相同,1条与父本斯特提棉相同,1条是杂种的特异性条带;在最后一对引物NAU2026中,有5条清晰的条带,其中有2条只与母本陆地棉相同,2条双亲的互补条带,还有一条是杂种的特异性条带。因此,在引物NAU1085、引物NAU1209、引物NAU3405、引物TMB1125和引物NAU2026中,共找到9条杂种新扩增出的特异性条带。除此之外,在这5对引物中有3对引物条带的分子量范围在100 bp-250 bp之间,有2对引物条带的分子量范围在250 bp-500 bp之间。
所有引物都扩增出了多态性产物。在这些扩增产物中,杂种通过引物NAU1209、引物NAU3405和引物NAU2026中扩增的条带结果表明其中有来自两个亲本的互补条带,在这5对引物中,都扩增出亲本所没有杂种的特异性条带。在这26条清晰的条带中,其中9条与母本陆地棉相同,2条与父本斯特提棉相同,6条是双亲的互补带,还有9条是两个亲本所没有的杂种新扩增出的特异性条带,所占的遗传成分比例分别是 34.62%、7.69%、23.07%和34.62%(表3)。结果初步表明了在母本陆地棉和父本斯特提棉的杂交过程中可能引起了染色体重组和片段交换,后续验证还在进行中。结果也表明陆地棉和斯特提棉的杂种中存在丰富的遗传变异,SSR分子标记鉴定结果分析证明该杂种是陆地棉和斯特提棉的真杂种。
3 讨论
3.1 野生斯特提棉的育种利用价值
棉属野生种具有很多栽培种所没有的优良品质,如丰产基因源,提高结铃性的有比克氏棉、瑟伯氏棉等;优质纤维基因源,其纤维品质主要的指标是长度、细度和强度,可以提高强度的有瑟伯氏棉、斯特提棉和毛棉等;抗病基因源,含有抗黄萎病基因的棉花主要有斯特提棉、澳洲棉、比克氏棉以及哈克尼西棉等。除此以外,棉属野生种还具有抗虫抗逆基因,细胞质雄性不育基因,无腺体基因,早苞落叶基因和致死基因[14]。野生斯特提棉具有抗根腐病,耐0-5℃低温及改进纤维强力的潜在特性,是棉花品种改良的优质材料,同时也具有抗虫、抗寒、抗霜、对光周期不敏感、抗黄萎病、枯萎病、生长势强、纤维优质和早熟等优良性状[15]。除此以外,Fxyxell[10]发现斯特提棉还具有许多陆地棉所没有的优良性状,如种子腺体延缓发育,种子在休眠状态时无腺体,在萌发后腺体开始形成,植株有腺体。
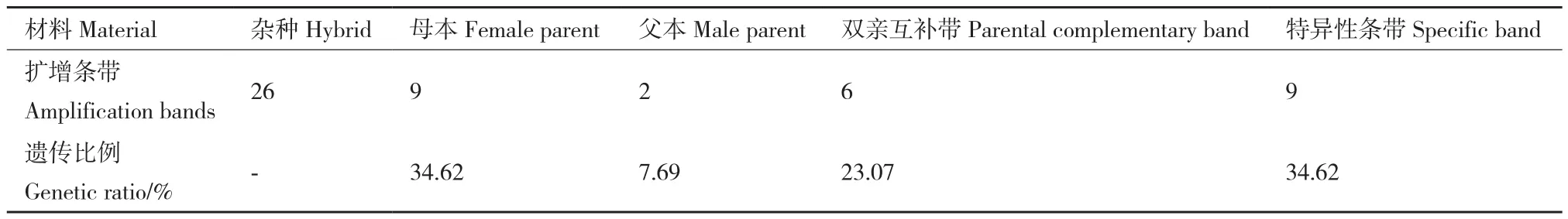
表3 杂种SSR的多态性来源分布Table 3 Source distribution of SSR polymorphisms in hybrid
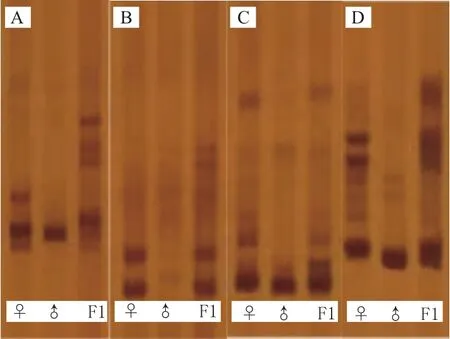
图6 SSR引物在杂种及父母本中的扩增结果Fig.6 Amplification results of SSR primers in hybrid and parents
因此,将斯特提棉的优良性状导入陆地棉中,创造出有育种价值的新材料。研究报道梁理民等[16]通过棉属种间杂交获得陆地棉×斯特提棉可育杂种,经过两次回交后,选育出优质、抗病性强、抗病大铃、抗病高衣分、抗病抗旱和抗黄萎病等多种优良新种质,又从陆地棉×斯特提棉杂种后代中育成抗病、优质、高产、抗旱、耐盐碱的棉花新品种秦远4号和抗病新种质6类24份,对丰富棉花遗传资源起到了重要的作用。崔淑芳等[17]利用海岛棉、陆地棉及野生棉进行棉属种间远缘杂交培育而成遗传基础丰富、高产、优质、抗病品种冀棉25,它的育成克服了远缘杂交障碍,拓宽了品种遗传基础。目前,从我国棉花远缘杂交所取得的成果看来,棉花的种质资源得到了创新,有大批已经或者即将进入棉花的原始资料库,许多野生棉的有利基因也已经进入基因库中,为棉花的种质创新提供了新材料[18]。
3.2 远缘杂种不育机理及育性恢复方法探讨
植物远缘杂交产生的杂种不育指的是杂种的第一代或者第二代植株虽然可以正常生长,但是因为远缘杂交不亲和,双亲的亲缘关系比较远,遗传差异大造成的染色体不平衡,在生理上也不协调,这些都会影响到雌雄配子不能形成合子,而导致不能生殖的现象[19]。远缘杂交不亲和的原因主要是亲缘关系较远的双亲在结构上、生理上还有遗传上的差异,染色体组间不协调等从而不能完成正常的受精,双亲的基因组成也对亲和性有影响[20]。李灵娇[21]通过对草雷杂种进行细胞遗传学鉴定发现草雷杂种高度不育的原因主要是杂种花粉母细胞减数分裂行为异常,同源染色体不能正常配对和不均等分离,形成许多异常孢子,导致大部分花粉粒败育。
长期以来国内外的棉花学家都将重点放在棉属种间杂交和棉花远缘杂种育性恢复探讨上,克服远缘杂交不亲和性。杂种育性恢复的主要方法有染色体加倍。在棉花育种中,也有很多远缘杂种由于双亲染色体组或染色体数目的不同而缺少了同源性,导致F1在减数分裂时染色体不能进行联会,从而不能形成足够数量有生活力的配子而导致不育。其主要解决方法是通过染色体诱导加倍和大量的回交和自交恢复其育性。如陆地棉×司笃克氏棉得到杂种三倍体F1不育,将其诱变加倍成异源六倍体恢复其育性[22];亚洲棉×司笃克氏棉得到杂种二倍体,将其诱变加倍成异源四倍体恢复其育性[23];以及本课题组前期合成陆地棉×斯特提棉杂种三倍体F1,申状状等[11]用0.2%的秋水仙碱琼脂凝胶涂抹生长点将其诱导加倍成异源六倍体,通过对M0、M1的育性统计结果分析发现,随着倍性的增加,育性正在逐渐恢复。郑赟等[24]通过陆地棉和C组野生澳洲棉远缘杂交获得杂种F1,对其进行染色体加倍,将加倍的异源六倍体再与B组野生绿顶棉杂交,产生陆地棉、澳洲棉、绿顶棉的异源四倍体杂种,进一步探讨有效的育性恢复方法。张伯静等[25]也采用染色体加倍和大量回交的方法获得了亚洲棉、比克氏棉和陆地棉的稳定可育的异源四倍体种质。雷蒙德氏棉、斯特提棉和陆地棉通过远缘杂交获得其三元杂种,并通过人工授粉与亲本进行的大量的回交和自交得到稳定的遗传后代[26]。吴玉香等[27]报道多年活体保存也是解决棉花种间杂种不育的途径之一,由于异源双亲染色体组的不同,造成了4个栽培种种间四元杂种高度不育,通过嫁接繁殖的方法延长杂种的时间,经过15年的生理协调,其雌雄配子的育性都得到了部分恢复,并对保存了15年的四元杂种从细胞学方面进行了鉴定,观察发现染色体的构型逐渐协调,染色体组趋于平衡。这一结果说明延长种间杂种活体的保存时间对于杂种的育性恢复是有利的。
3.3 远缘杂种SSR分子标记鉴定分析探讨
SSR(simple sequence repeats)是简单重复序列长度多态性标记,以PCR为基础快速简便,品种间多态性丰富,对杂交种及其亲本的鉴定更具优势。近年来,SSR分子标记因其具有高丰度性、成本低、耗时少、多态性高和重复性好,已广泛应用于棉花种质鉴定中[28]。石建斌等[29]采用前期筛选出的26对SSR核心引物,对收集保存的58份棉花种质资源进行纯度检测和遗传多样性分析。赵程杰等[30]利用SSR标记对53个棉花品种进行纯度检测和真实性鉴定。Larbouga等[31]利用SSR分子标记多态性对121份棉花种质资源的遗传多样性进行了分析。申状状等[13]对亚瑟远缘杂种进行了SSR分子鉴定,进一步印证了杂种的真实性。王一帆等[32]对杂交育成的亚拟杂种进行SSR分子标记鉴定,进一步从分子水平证明了杂种是组合亚洲棉和拟似棉遗传成分的真杂种。本实验对陆地棉和斯特提棉的远缘杂种也进行了SSR分子标记鉴定,结果表明:杂种F1不仅扩增出双亲的互补带,还扩增出双亲没有的特异性条带,遗传成分比例分别为:父本占7.69%,母本占34.62%,双亲的互补带占23.07%,特异性新带占34.62%,既表明在杂交过程中可能引起了染色体重组和片段交换,也从分子水平证实了该杂种是陆地棉和斯特提棉的真杂种,同时为棉花遗传育种和种质创新提供了有价值的材料。
4 结论
本研究对陆地棉与野生斯特提棉远缘杂种进行了形态学、细胞遗传学以及SSR分子标记分析和鉴定,形态性状表明杂种介于双亲之间,并表现出显著的杂种优势;细胞遗传学进一步揭示了杂种不育的主要原因;SSR分子标记从分子水平证实了远缘杂种的真实性,同时表明在杂交过程中发生了基因重组。
