不同抗性棉花品种PR基因响应大丽轮枝菌的表达分析
2021-05-13何芳孙琦李彪刘政黄家风
何芳,孙琦,李彪,刘政,黄家风
(石河子大学农学院/新疆绿洲农业病虫害治理与植保资源利用重点实验室,新疆 石河子832003)
大丽轮枝菌(Verticilliumdahliae)引起的棉花黄萎病是当前威胁我国棉花生产最主要的病害,每年在发病严重的地区造成棉花减产高达30%以上[1]。大丽轮枝菌沿植物根部维管组织向上侵染,其形成的微菌核在土壤中长期存活[2],常规农业防治和药剂防治难以奏效,因此针对该病害的防治主要集中在选育和利用抗病品种上。目前推广应用的棉花品种抗黄萎病性能不断提高,但是黄萎病依然发生严重,究其原因除了陆地缺乏棉抗黄萎病种质资源外[3],病原种群结构发生变化也是一个重要因素[4]。一般认为棉花黄萎病菌落叶型致病力比非落叶型致病力强[4-5],落叶型菌系呈现逐年增加趋势,已逐步取代非落叶型菌系成为棉田的主要致病类型[4,6]。然而不同的棉花品种响应大丽轮枝菌的侵染是否存在差异尚不清楚。
植物与病原互作中,植物为抵御病原物侵染,通过细胞膜上的模式识别受体(pattern recognition receptors,PRR)识别病原物相关分子模式(pathogen-associated molecular patterns,PAMP)触发基础免疫反应[7-8],如气孔关闭、活性氧爆发、胼胝质沉积以及病程相关(Pathogenesis related,PR)基因的表达等,以达到抑制病原物生长,实现植物对病原物的防御[9-10]。植物的PR基因既能响应生物胁迫也能响应非生物胁迫[11-12],其编码的PR蛋白在植物体外表现出潜在的抗菌活性,在植物体内的积累与植物的抗性反应密切相关[11]。根据来源植物、电泳迁移率、氨基酸序列的同源性和生化功能等特性,PR蛋白基因分为17个家族[12-13]。PR1是第1个被发现的多基因家族,表达PR1基因的转基因烟草和番茄可明显增强对卵菌和真菌的抵抗能力[14-15],并且PR1基因已被广泛用于过敏性反应(HR)介导的防御途径和水杨酸介导的抗性反应被激活的指示基因[16]。PR2基因家族编码β-1,3葡聚糖酶,能催化降解病原真菌细胞壁中的β-1,3葡聚糖成分[17]。PR3、PR4、PR8、PR11基因家族编码几丁质酶,能够降解病原真菌细胞壁中的几丁质成分[17-20]。PR5基因家族编码的蛋白包括 permatins、zeamatins、osmotins和类甜蛋白(thaumamatin like proteins,TLP),具抗真菌活性[21],分别过表达烟草和茶树TLP基因的转基因马铃薯均表现明显的抗真菌特性[22-23]。PR6基因家族编码蛋白酶抑制剂(PI),其靶标是昆虫和微生物的蛋白酶,体内过表达PR6基因的拟南芥和烟草能增强对灰霉菌的抗性[11,24]。PR7基因编码的蛋白最早从番茄中提纯获得,是由柑橘裂皮类病毒(Citrusexocortisviroid)诱导产生的一种胞内蛋白酶[25]。PR9基因家族编码过氧化物酶是细胞壁构建过程中的关键酶,当植物遭受病原物侵染时,PR9蛋白通过促进木质素沉积,增加细胞壁机械性能以抵御病原物入侵[26]。PR10基因家族编码的蛋白具有核酸酶活性,如甜椒CaPR10基因受烟草花叶病毒(TMV)诱导表达产生的核酸酶,通过降解病毒核酸使甜椒具有更强的抗病毒能力[27],此外,PR10蛋白还具有配体结合特性和翻译后修饰等功能[21]。PR12基因家族编码植物防御素,通过抑制真菌生长使植物具有抗真菌特性,如过表达萝卜防御素基因的转基因烟草对长柄链格孢产生抗性[11,28]。PR13基因家族编码硫素蛋白,异源表达拟南芥硫素基因的转基因番茄和异源表达大麦硫素基因的转基因烟草分别对镰刀菌和丁香假单胞菌产生抗性,表明硫素蛋白对植物病原真菌和细菌都具有抗菌活性[29-30]。PR14基因家族编码脂质转运蛋白(LTPs),功能包括角质素合成、生物氧化、植物系统获得抗性的信号转导、以及抗真菌和细菌活性;异源表达大麦LTP基因的转基因拟南芥对丁香假单胞菌和灰霉菌都能产生抗性[11,31]。PR15和PR16基因编码的蛋白都具有草酸氧化酶活性,在大麦受白粉菌侵染时诱导表达,并通过产生过氧化氢参与植物防御反应[32-33]。PR17基因受病原物诱导表达并能与病原物效应子直接互作,如烟草NtPRp27基因和小麦WCI-5基因分别受TMV和小麦白粉病菌诱导表达并积累[34-35],大麦HvPR-17c通过与布氏白粉病菌(Blumeriagraminis)的效应子互作阻止病原菌侵染[36]。
杜旋等[37]研究发现大丽轮枝菌 PAMP诱导处理后GhPR基因普遍上调表达,但是对于不同抗性陆地棉品种受大丽轮枝菌侵染以及同一个抗性品种受不同致病型大丽轮枝菌诱导后,14个GhPR基因表达是否存在差异,目前尚未明确。本文研究抗、感病棉花品种中GhPR基因响应寄主和非寄主来源的落叶型和非落叶型大丽轮枝菌的表达差异,为进一步揭示棉花-大丽轮枝菌互作机制奠定基础。
1 材料与方法
1.1 供试菌株
供试的大丽轮枝菌菌株共有4种,其中棉花黄萎病菌落叶型菌株V592和非落叶型菌株I6均分离自新疆棉花(棉区栽培品种),由本实验室分离保存;非落叶型菌株V31分离自宁波茄子,由宁波动植物出入境检验检疫局段维军惠赠;落叶型菌株S1分离自内蒙古向日葵,由内蒙古农业大学景岚教授惠赠。以上供试菌株的致病力和致病型均得到验证[38]。
1.2 供试棉花品种
高感黄萎病品种为新陆早1号和军棉1号,抗黄萎病棉花品种为中植棉2号。
1.3 无菌棉苗的培育
挑选脱绒后籽粒饱满的棉花种子,剥去种皮,用0.1%升汞消毒20 min,期间多次震荡混匀,无菌水冲洗5或6遍,放入MS培养基中,28 ℃暗培养,3 d后长出的无菌棉苗扶正,放入盛有无菌水小烧杯,再转入大烧杯中封口,28 ℃光照培养备用。
1.4 大丽轮枝菌对棉苗根部的处理
将PDA平板上培养7 d的菌株用10 mm打孔器取10块菌饼,放入Czapek-Dox培养基中26 ℃,200 r/min摇培 3 d,4层纱布过滤收集真菌的分生孢子,用无菌水调整孢子浓度至107cfu/mL。在含200 mL无菌水培养的棉苗中加入1 mL孢子悬浮液,然后分别在接种3、6、12 h剪下棉花根系,无菌水冲洗后液氮速冻保存或直接进行RNA提取。以无菌水处理的棉花作为阴性对照。每一处理3次重复。
1.5 棉花根组织总RNA提取和实时定量PCR(RT-qPCR)分析
利用EASYspin Plus植物RNA快速提取试剂盒(北京艾德莱生物科技有限公司)提取棉花根组织总RNA,于-80 ℃保存备用,每个材料取3株样品;利用Prime Script RT reagent Kit with gDNA Eraser(北京宝日医生物技术有限公司)系统对RNA进行反转录,然后利用PowerUp SYBR Green Master Mix(Life Technologies)试剂盒和7500 Real Time PCR System(Life Technologies)系统对目标基因GhPR的转录表达通过qPCR进行测定。所用引物见表1,以Ghactin基因(DQ266153)作为内参基因[37]对目标基因的转录水平进行标准化;将无菌水处理对照棉花的GhPR基因表达水平设为1,将标准化的GhPR基因表达水平与对照GhPR基因表达水平进行比较,获得各个GhPR基因转录水平的相对表达量;每个样品设3个重复。qPCR反应程序为95 ℃,10 min;95 ℃,15 s;60 ℃,60 s;40 cycles。

表1 检测陆地棉GhPRs基因转录表达所用引物[37]
根据2-ΔΔCt方法对数据进行处理,试验数据采用SPSS 17.0软件进行统计分析。
2 结果与分析
2.1 感病棉花品种GhPR基因响应棉花黄萎病菌的表达分析
研究感病棉花品种的GhPR基因响应不同致病型大丽轮枝菌的表达差异中,数据采用Dunnett法进行显著性差异检验(**P<0.01),误差线表示3个重复的标准误差。首先用棉花黄萎病菌的落叶型菌株V592和非落叶型菌株I6对感病品种军棉1号进行诱导处理,结果见图1。
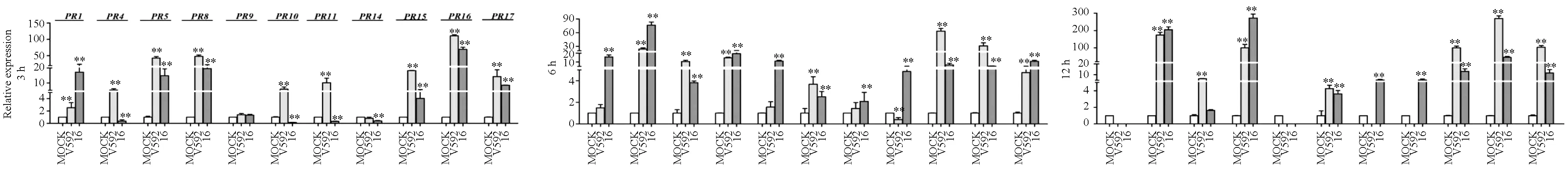
图1 V592和I6菌株诱导处理感病品种军棉1号后GhPR基因转录水平的表达情况
图1显示:V592菌株诱导9个GhPR基因上调表达,I6菌株诱导11个GhPR基因上调表达,多数GhPR基因在3 h即出现上调表达,并持续至6 h或12 h。比较2个菌株诱导的每个GhPR基因的最高相对表达量可知:落叶型菌株V592在3 h诱导GhPR5、GhPR10、GhPR11基因和12 h诱导GhPR15、GhPR16和GhPR17基因的相对表达量达到最大,分别是阴性对照的42.9倍、5.7倍、10.1倍、100.9倍、269.8倍和103.5倍;而I6菌株在3 h诱导GhPR5基因、12 h诱导GhPR10、GhPR11、GhPR15、GhPR16和GhPR17基因的最大相对表达量分别是阴性对照的14.8倍、3.6倍、5.3倍、13.1倍、69.4倍和11.8倍,明显低于V592诱导处理后的表达量,表明落叶型菌株V592诱导这 6个基因上调表达的程度比非落叶型菌株I6强。然而经I6菌株诱导处理,GhPR1、GhPR4、GhPR8、GhPR9和GhPR145个基因分别在3 h、12 h、12 h、6 h和12 h时相对表达量达到最高,分别是阴性对照的17.2倍、204.3倍、272.4倍、11.1倍和5.1倍,明显高于经V592菌株诱导处理后的表达量,V592菌株诱导GhPR1、GhPR4和GhPR8基因的最高相对表达量分别是阴性对照的2.5倍(3 h)、173.6倍(12 h)和99.5倍(12 h),而GhPR9和GhPR14基因受V592菌株诱导并不表现上调表达,表明非落叶型菌株I6诱导GhPR1、GhPR4、GhPR8、GhPR9和GhPR145个基因上调表达的程度比落叶型菌株V592强。上述结果表明,在棉花感病品种军棉1号上,棉花黄萎病菌落叶型菌株和非落叶型菌株均能诱导不同的GhPR基因上调表达,但2种致病型黄萎病菌诱导上调表达的GhPR基因个数及表达量存在差异。
再用V592菌株和I6菌株对感病品种新陆早1号进行诱导处理,结果见图2。
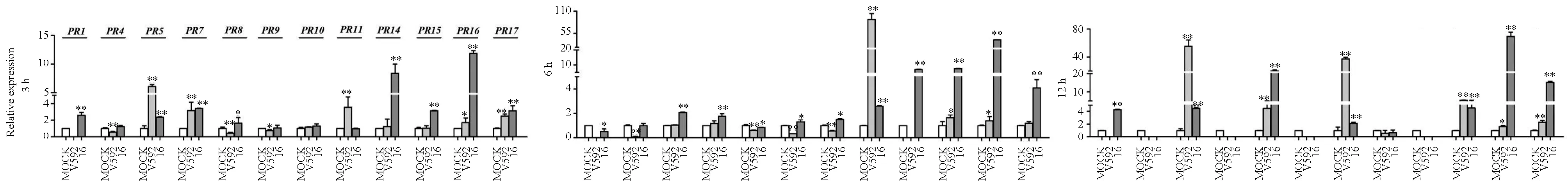
图2 V592和I6菌株诱导处理感病品种新陆早1号后GhPR基因转录水平的表达情况
图2显示:落叶型菌株V592诱导7个GhPR基因上调表达,非落叶型菌株I6诱导10个GhPR基因上调表达。对2个菌株诱导的每个GhPR基因的最高相对表达量进行比较可知:落叶型菌株V592在12 h诱导GhPR5和GhPR10基因、6 h诱导GhPR11基因的最大相对表达量分别是阴性对照的55.3倍、37.4倍和89.1倍;而I6菌株在12 h诱导GhPR5和GhPR10基因、6 h诱导GhPR11基因的最高相对表达量分别是对照的4.5倍、2.1倍和2.6倍,明显低于V592诱导处理后的表达量,表明在感病品种新陆早1号上,落叶型菌株V592诱导这3个基因上调表达的程度比非落叶型菌株I6强。然而经I6菌株诱导处理,GhPR8、GhPR15和GhPR17基因分别在12、6、12 h时相对表达量达到最高,分别是对照的20.1倍、7.6倍和15.2倍,明显高于V592菌株对这3个基因的诱导表达,V592菌株均在12 h诱导GhPR8、GhPR15和GhPR17基因达到最大相对表达量,分别是对照的4.5倍、5.3倍和2.5倍。I6在12 h诱导GhPR1基因、3 h诱导GhPR14基因、12 h诱导GhPR16基因的最大相对表达量分别是对照的4.3倍、8.4倍和69.4倍,而这3个基因均不受V592菌株诱导表达。因此在感病品种新陆早1号上,非落叶型菌株I6诱导GhPR1、GhPR8、GhPR14、GhPR15、GhPR16和GhPR176个基因上调表达的程度比落叶型菌株V592强。
上述结果表明:在棉花感病品种新陆早1号上,棉花黄萎病菌落叶型菌株V592诱导上调表达的GhPR基因总数及相对表达量达到最大的GhPR基因数都没有I6菌株诱导的GhPR基因多。
2.2 抗病棉花品种GhPR基因响应棉花黄萎病菌的表达分析
为了研究抗病棉花品种的GhPR基因响应不同致病型大丽轮枝菌的表达差异,用落叶型菌株V592和非落叶型菌株I6对抗病品种中植棉2号进行诱导处理,结果见图3。

图3 V592和I6菌株诱导处理抗病品种中植棉2号后GhPR基因转录水平的表达情况
图3显示:V592菌株诱导9个GhPR基因上调表达,不诱导GhPR7、GhPR9和GhPR11基因表达;I6菌株诱导11个GhPR基因上调表达,不诱导GhPR9基因表达。对2个菌株诱导的每个GhPR基因的最高相对表达量进行比较可知:虽然I6菌株可诱导11个GhPR基因上调表达,但每个基因上调表达的幅度不大,其中GhPR16基因上调幅度最高,12 h时其最高相对表达量是对照的22.6倍,GhPR5基因上调幅度最低,12 h时其最高相对表达量是对照的2.5倍。然而V592菌株诱导上调表达的9个基因,其中8个基因均在诱导6 h时相对表达量达到最高,与阴性对照相比,每个GhPR基因的上调表达幅度显著增高,如GhPR5和GhPR16基因的最高相对表达量分别是对照的1928.5倍和1407.3倍,远远高于I6菌株诱导GhPR5和GhPR16基因最高相对表达量分别是对照的2.5倍和22.6倍;V592菌株诱导GhPR8、GhPR14和GhPR15基因的最高相对表达量分别是对照的359.4倍、489.8倍和394.7倍;GhPR4、GhPR10和GhPR17基因的最高相对表达量分别是对照的178.1倍、192.6倍和118.7倍;GhPR1基因上调表达的幅度最小,3 h时其最高相对表达量也是对照的75.9倍。
综上可知:在棉花抗病品种中植棉2号上,落叶型菌株V592诱导9个GhPR基因上调表达的程度明显比非落叶型菌株I6强,且GhPR基因上调表达的幅度显著增强。
2.3 感病棉花品种GhPR基因响应非寄主来源大丽轮枝菌的表达分析
为了明确感病棉花品种的GhPR基因响应非寄主作物来源大丽轮枝菌的表达差异,分别用分离自向日葵的落叶型菌株S1和茄子上的非落叶型菌株V31诱导处理感病棉花品种军棉1号,结果见图4。
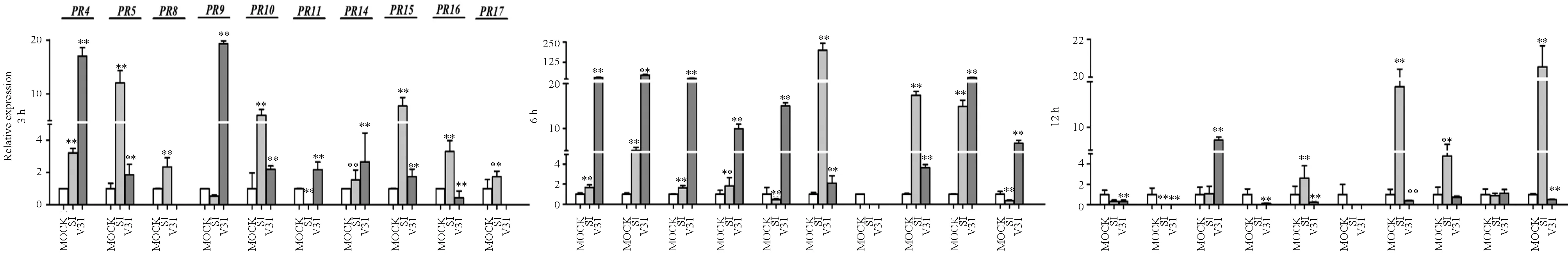
图4 其他寄主来源的大丽轮枝菌诱导处理感病品种军棉1号后GhPR基因转录水平的表达情况
图4显示:S1和V31菌株均诱导10个GhPR基因上调表达。比较2个菌株诱导的每个GhPR基因的最高相对表达量可知:落叶型菌株S1在6 h诱导GhPR11和GhPR15基因、12 h诱导GhPR14和GhPR17基因上调表达幅度最大,其最高相对表达量分别是对照的198.6倍、17.4倍、18.9倍和20.5倍;而V31诱导这4个基因的最大相对表达量分别是对照的2.2倍、3.7倍、2.7倍和6.7倍,因此S1诱导GhPR11、GhPR15、GhPR14和GhPR174个基因达到的最高相对表达量明显高于V31诱导的表达量。非落叶型菌株V31在3h诱导GhPR9基因、6h诱导GhPR4、GhPR5、GhPR8、GhPR10和GhPR16基因上调表达的最大相对表达量分别是对照的19.3倍、28.5倍、46.2倍、23.2倍、15.0倍和28.7倍,明显高于S1诱导这6个基因在不同时段的最高相对表达量,S1诱导GhPR9、GhPR4、GhPR5、GhPR8、GhPR10和GhPR166个基因的最大相对表达量分别是对照的1.8倍、3.2倍、12.0倍、2.4倍、6.0倍和14.8倍。
再用非寄主作物作物来源的2个菌株诱导处理感病品种新陆早1号,结果(图5)显示:S1菌株诱导11个GhPR基因上调表达,不诱导GhPR5基因上调表达;V31菌株诱导12个GhPR基因上调表达。比较各菌株诱导的每个GhPR基因的最高相对表达量可知:S1菌株诱导5个GhPR基因的上调表达量明显高于V31菌株,即S1菌株在12h诱导GhPR4基因、6h诱导GhPR1、GhPR9、GhPR10和GhPR15基因上调表达幅度最大,其最高相对表达量分别是对照的20.7倍、38.0倍、38.0倍、38.0倍和19.6倍;而V31诱导这5个基因达到的最高相对表达量分别是对照的17.4倍、6.2倍、31.8倍、3.4倍和11.1倍。V31菌株诱导7个GhPR基因的上调表达量明显高于S1菌株,即V31菌株在6h诱导GhPR5、GhPR7和GhPR14基因、12h诱导GhPR8、GhPR11和GhPR16基因、3h诱导GhPR17基因上调表达幅度最大,其最高相对表达量分别是对照的4.5倍、7.4倍、15.0倍、137.6倍、14.4倍、152.2倍和9.1倍;而S1诱导这7个基因达到的最高相对表达量分别是对照的1.6倍、4.1倍、3.5倍、10.2倍、9.9倍、54.8倍和7.7倍。
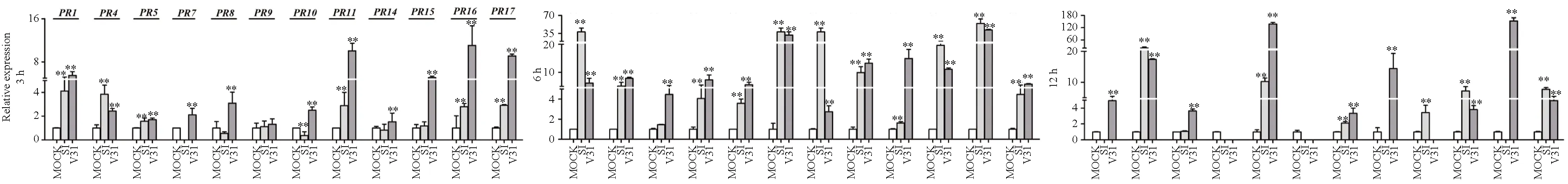
图5 其他寄主来源的大丽轮枝菌诱导处理感病品种新陆早1号后GhPR基因转录水平的表达情况
上述结果表明:大丽轮枝菌非寄主作物来源的落叶型菌株S1和非落叶型菌株V31均可诱导感病品种军棉1号和新陆早1号不同的GhPR基因上调表达,非落叶型菌株V31诱导GhPR基因达到最大相对表达量的基因个数比落叶型菌株S1诱导的多。
2.4 抗病棉花品种GhPR基因响应非寄主来源大丽轮枝菌的表达分析
用非寄主作物来源的2个菌株诱导处理棉花抗病品种中植棉2号,结果(图6)显示:S1菌株和V31菌株均诱导12个GhPR基因上调表达。对2个菌株诱导的每个GhPR基因的最高相对表达量进行比较可知:S1菌株诱导9个GhPR基因的上调表达量明显高于V31菌株,即S1菌株在3 h诱导GhPR4、GhPR14、GhPR15和GhPR174个基因上调表达的最高相对表达量分别是对照的186.0倍、36.9倍、31.5倍和14.4倍;12 h诱导GhPR1、GhPR8、GhPR9、GhPR10和GhPR115个基因上调表达的最高相对表达量分别是对照的40.6倍、7.6倍、8.5倍、22.2倍和174.8倍;V31诱导这9个基因达到的最高相对表达量分别是对照的8.5倍、21.2倍、3.1倍、1.9倍、13.1倍、3.7倍、1.5倍、5.4倍和14.4倍。而V31菌株仅诱导2个GhPR基因的最大相对表达量明显高于S1菌株,即V31菌株在3 h诱导GhPR5和GhPR16基因上调表达的最高相对表达量分别是对照的6.7倍和32倍;S1诱导这2个基因达到的最高相对表达量分别是对照的5.8倍、6.2倍。
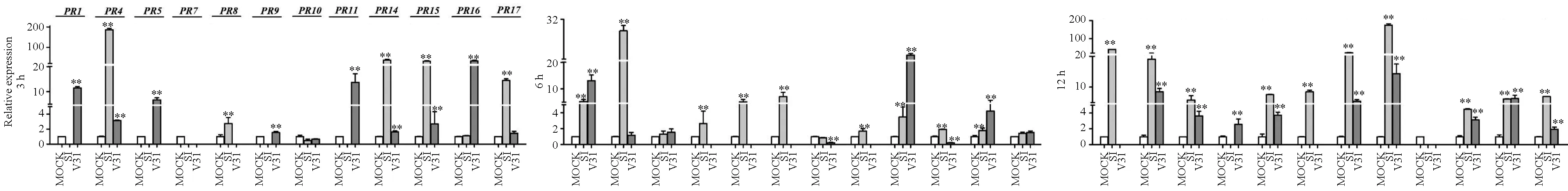
图6 其他寄主来源的大丽轮枝菌诱导处理抗病品种中植棉2号后GhPR基因转录水平的表达情况
上述结果表明,大丽轮枝菌非寄主作物来源的落叶型菌株S1和非落叶型菌株V31均可诱导抗病品种中植棉2号不同的GhPR基因上调表达,落叶型菌株S1诱导GhPR基因达到最大相对表达量的基因个数明显多于V31菌株。
3 结论与讨论
(1)在2个感病棉花品种上,寄主来源的落叶型和非落叶型黄萎病菌均能诱导GhPR基因普遍上调表达,而在感病品种军棉1号上,2个菌株诱导GhPR基因上调表达的能力相当;在感病品种新陆早1号上,非落叶型菌株I6诱导GhPR基因上调表达的能力比落叶型菌株V592强,表明棉花感病品种的GhPR基因对落叶型和非落叶型黄萎病菌侵染均有一定的响应能力。然而感病品种对2种致病类型黄萎病菌的侵染却表现明显不同的抗性反应:对于非落叶型菌株的侵染表现一定的抗病性,如2000年以前,新疆棉区黄萎病菌以非落叶型菌系为主[39],没有落叶型黄萎病菌的发生,虽然当时种植的棉花品种多为感病品种,但棉花黄萎病发生并不严重,推测可能与非落叶型菌系能够诱导感病品种不同的GhPR基因上调表达,诱导了植物免疫有关;尽管感病品种的GhPR基因对落叶型黄萎病菌也有一定的响应能力,但是感病品种对于落叶型菌株的侵染却表现高度敏感[38],其原因是否与落叶型黄萎病菌比非落叶型黄萎病菌产生更多或更强的效应子蛋白,能够克服感病品种的免疫应答有关,有待进一步研究。
(2)在棉花抗病品种中植棉2号上,落叶型菌株V592诱导GhPR基因上调表达的强度明显比非落叶型菌株I6强,如V592诱导中植棉2号GhPR5和GhPR16基因的最高相对表达量分别是对照的1928.5倍和1407.3倍,表明棉花抗病品种的GhPR基因对棉花黄萎病菌落叶型菌系具有更强的响应能力。然而尽管目前推广应用的棉花品种抗病性不断增强,但是近年棉花主产区黄萎病却逐年加重,落叶型黄萎病菌已逐步取代非落叶型菌系成为棉田只要的致病类型[6],说明随着品种抗性的不断增强,病原菌致病性与之协同进化也在不断增强。有研究表明,大麦的HvPR17c(PR17蛋白)可以与布氏白粉病菌(Blumeriagraminis)的效应子直接互作阻止病原菌侵染[36];也有研究表明,棉花黄萎病菌产生的效应子PevD1可以抑制GhPR5蛋白的抗真菌活性从而克服寄主的防御反应[40]。虽然目前在棉花-黄萎病菌中没有发现类似于番茄大丽轮枝菌1号生理小种AVe1基因诱导含Ve1基因的番茄产生抗病性的互作关系[41],但是棉花抗病品种的GhPR基因响应落叶型黄萎病菌表现强烈表达可能与落叶型黄萎病菌产生了更多有效的效应子激活了寄主的抗性反应有关,也可能与抗病品种中含有更可以多识别效应子的抗病蛋白有关,但是GhPR蛋白-效应子-抗病蛋白之间存在怎样的互作关系有待深入研究。
用非寄主作物的2个大丽轮枝菌菌株分别处理2个感病棉花品种和1个抗病棉花品种,结果表明:非落叶型菌株V31诱导感病品种GhPR基因上调表达的能力比落叶型菌株S1强;落叶型菌株S1诱导抗病品种GhPR基因上调表达的能力明显比V31菌株强。这与寄主来源的V592和I6菌株分别在抗病和感病品种上的诱导结果基本一致。从基因上调表达的幅度来看,在抗病品种上,非落叶型菌株I6和V31诱导GhPR基因表达的上调幅度都比较小;但是对于落叶型菌株,非寄主来源的S1菌株诱导GhPR基因上调表达幅度明显低于寄主来源的V592。表明抗病品种GhPR基因响应寄主来源落叶型菌株的能力比响应非寄主来源落叶型菌株的能力强。
(3)杜旋等根据大丽轮枝菌坏死和乙烯诱导蛋白(necrosis and ethylene-inducing peptide 1-like protein,NLP)设计了3个多肽片段nlp20Vd2、nlp20Vd2和 nlp20Vd2作为PAMP分子诱导处理棉花感病品种新陆早6号,结果显示,3个多肽片段都能诱导GhPR基因上调表达,除了nlp20Vd2诱导的2个GhPR基因上调幅度与对照相比分别达到13和16倍外,其他GhPRs基因的上调幅度与对照相比都在2~6倍之间[37]。而本研究直接用大丽轮枝菌处理棉花根系,非落叶型菌株诱导感病品种GhPR基因的上调幅度和落叶型菌株诱导抗病品种GhPR基因的上调幅度均普遍高于单个肽段诱导GhPR基因的上调幅度,尤其是落叶型菌株诱导抗病品种GhPR基因的上调幅度远远超过单个肽段诱导GhPR基因的上调幅度,这可能与大丽轮枝菌具更多的PAMP分子有关,因为除了NLP蛋白具有PAMP活性,植物病原真菌的几丁质寡糖、木聚糖酶、多聚半乳糖醛羧酶(PGs)都可作为PAMP分子触发PTI[42],激活包括PR基因表达在内的植物免疫反应。
(4)比较3个棉花品种响应棉花黄萎病菌侵染,GhPR5、GhPR8、GhPR10、GhPR15、GhPR17这5个基因均表现明显上调表达,且大多数在诱导处理 6 h时表达量达到最大,说明这5个基因可作为棉花响应棉花黄萎病菌免疫反应的指示基因,诱导处理6 h时为最佳测定时间;GhPR4、GhPR5、GhPR8、GhPR10、GhPR14、GhPR15、GhPR16和GhPR17均在诱导处理6 h时表达量最高,可作为抗病品种响应落叶型黄萎病菌免疫反应的指示基因。另外,本研究发现,3个品种的GhPR2和GhPR3基因响应大丽轮枝菌侵染均不表现明显上调表达,有的甚至是下调表达,其原因有待进一步研究。
