原料乳离心除菌对UHT乳品质的改善作用
2021-05-12孙玥吴华星段雪梅李琳赵广生马莺李启明
孙玥,吴华星,段雪梅,李琳,赵广生,马莺,李启明
(1.哈尔滨工业大学化工与化学学院,哈尔滨 150090;2.新希望乳业股份有限公司,成都 610063;3.乳品营养与功能四川省重点实验室,成都 610000;4.杭州新希望双峰乳业有限公司,杭州 310000)
0 引 言
热处理是保证乳制品安全最重要的手段,超高温瞬时杀菌(Ultra-high temperature,UHT)是牛乳在135~150℃下加热3~5 s,然后无菌包装于微量透气的容器中,生产出来的产品称UHT乳[1]。UHT乳可以在环境温度下保存一年,通常认为可接受的最佳时间是6个月。UHT热处理可杀死细菌、体细胞以及酶,但不能杀死全部的耐热芽孢菌,一些耐热性内源酶、耐热芽孢产生的酶、以及微生物及体细胞自溶所释放的胞内酶在UHT乳贮藏期间保持活性,严重地影响了UHT乳的质量[2]。离心除菌是基于颗粒(微生物、脂肪球、不溶性蛋白质等)和分散相之间密度的差异除去原料乳体细胞和细菌。离心除菌机是专门设计在55~65℃下以9 000g的转速高速离心牛乳(离心时间<1 s),可以将细菌总数减少86.0%~92.0%,芽孢去除率为90.0%~98.0%,厌氧芽孢杆菌的去除率为97.4%~98.7%,需氧芽孢杆菌的去除率为94.1%~97.7%[3]。嗜冷菌是导致牛乳变质的主要微生物,大多数嗜冷菌产生的脂肪酶和蛋白酶显著降低了巴氏杀菌乳和UHT乳及其他乳制品的质量和保质期[4]。离心除菌要求原料在55~60℃预热,预热足以消除生乳中99.99%的嗜冷菌,离心除菌可以在预热的基础上进一步去除89.66%的嗜冷菌[5]。生乳中含有的体细胞在热处理过程中对单核细胞增生李斯特菌有保护作用,体细胞中含有大量的酶(过氧化氢酶、蛋白酶和脂肪酶),牛乳贮藏过程中失活的体细胞分泌的酶也对乳质量产生影响[6]。离心除菌可以将95%的体细胞去除[3]。离心除菌技术可以显著地降低原料乳微生物和体细胞数量,也避免了微生物菌体和失活体细胞分泌的酶对在贮藏过程乳制品品质的影响。
在同一乳品厂,按照相同的工艺生产两批常规UHT(N-UHT)乳和离心除菌UHT乳(C-UHT),每一批的原料乳相同。研究了离心除菌对UHT乳货架期质量品质的改善作用。
1 材料与方法
1.1 材料与设备
牛乳来源于新希望自有牧场,原乳细菌总数小于2×105cfu/mL,体细胞小于5×105个。WSC-S色差计,上海仪电物理光学仪器有限公司;Zetasizer Nano ZS纳米粒度及Zeta电位分析仪,英国Malvern公司;Kinexus Pro+流变仪,英国Malvern公司。
1.2 实验方法
在同一乳品厂,按照相同的工艺分别生产了两批常规UHT(N-UHT)乳和离心除菌UHT乳(C-UHT),每一批的原料乳相同。N-UHT乳杀菌参数为135℃/4 s;C-UHT工艺参数为:55~60℃原料乳在7 200g下离心,回流速率为300~400 kg/h,离心后的原料乳经过135℃/4 s杀菌。UHT乳在常温下贮藏6个月,每间隔1个月取样。测定乳样品的白度、粒度、zeta电位、流变性、酸度、过氧化值、纤溶蛋白酶、总蛋白、乳清蛋白、酪蛋白、非蛋白氮的变化。
2 结果与讨论
2.1 贮藏期间UHT乳的物理性质变化
牛乳是由脂肪分散相和水胶体连续相组成的胶体溶液。牛乳的物理性质与水相似,但由于连续相中存在各种溶质(蛋白质、乳糖和盐)以及乳化和胶体组分的分散程度而改变,超高温杀菌强度会导致牛乳的化学成分和物理属性发生变化[7]。
2.1.1 色泽
牛乳是白色不透明的乳状液,牛乳加工和贮藏过程中颜色变化主要与乳糖和乳蛋白之间的美拉德反应有关[8]。白度是表示物质表面白色的程度,以白色含量的百分率表示,0为黑,100为白。常规UHT乳(N-UHT)和离心除菌UHT乳(C-UHT)在6个月贮藏期内白度值(%)变化如图1所示。
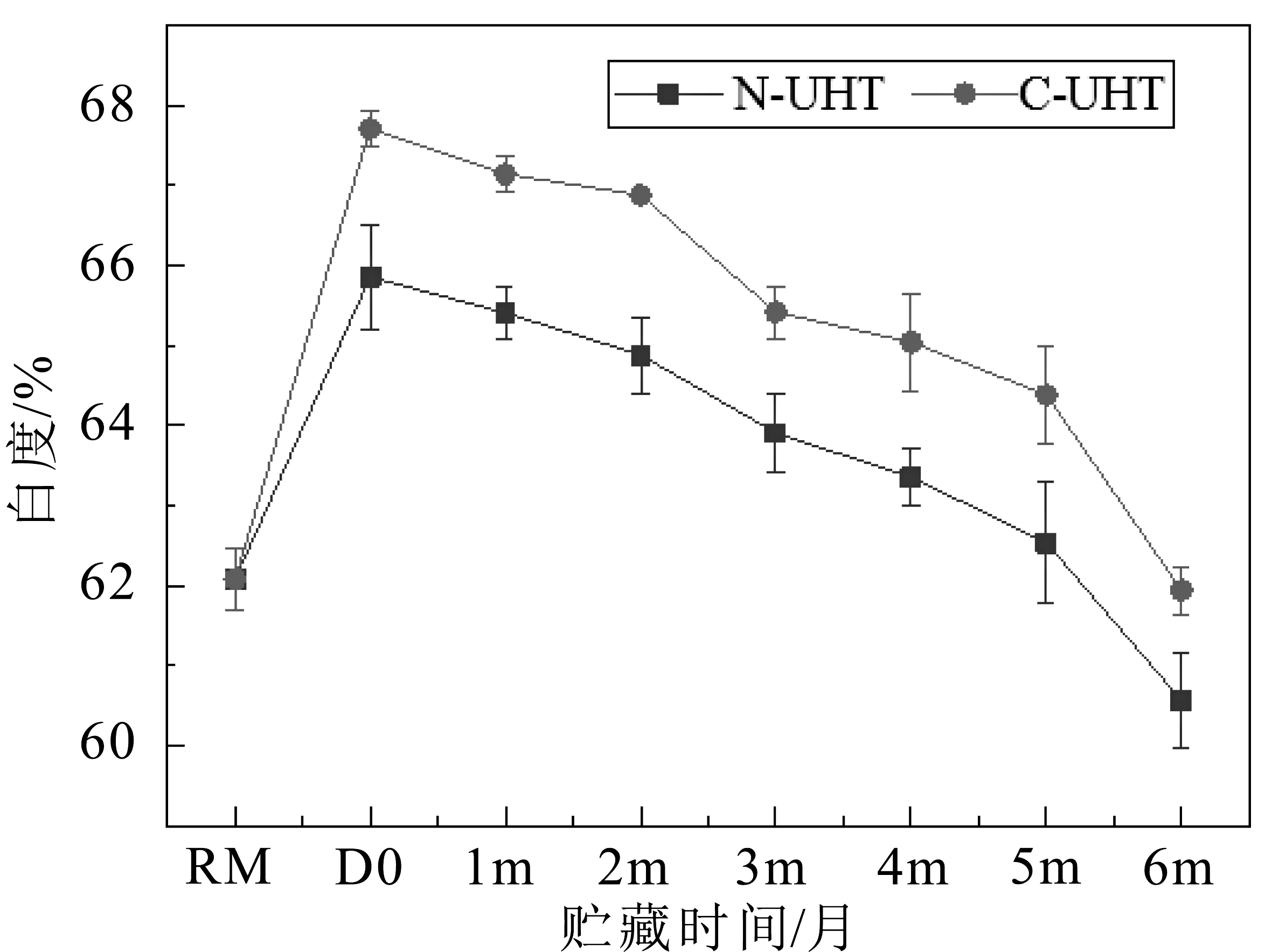
图1 UHT乳贮藏期间白度值变化
与原乳(RM)比较,两种UHT乳的白度值增加,说明热处理后UHT乳颜色变白。两种UHT乳在贮藏过程中白度值逐渐降低,N-UHT乳的白度值显著低于C-UHT乳,即N-UHT乳比C-UHT乳颜色深(图1)。杀菌后UHT乳的白度值高于原乳,是由于热处理导致乳清蛋白变性,变性乳清蛋白与酪蛋白胶束聚集,胶束粒径增大,乳颗粒对光反射率增强,使乳白度增加[9]。在贮藏过程中蛋白质和乳糖之间的美拉德反应持续进行,进而产生糠氨酸、羟甲基糠醛以及类黑精等其他褐色物质,影响了UHT乳的色泽[10]。贮藏过程中褐变程度受热处理强度的影响,热处理强度较高的牛乳即使在热处理后立即变白,在贮藏过程中也会以更快的速度变褐[11]。UHT乳中残余脂肪酶和蛋白酶将蛋白质和脂肪部分被水解,影响了反射粒子的数量[12]。离心除菌工艺将原料乳中体细胞和90%微生物除去,C-UHT乳中微生物源蛋白酶和脂肪酶数量低N-UHT乳,由残余酶间接引起的色泽改变程度低于N-UHT乳。因此,离心除菌UHT乳比常规UHT乳在贮藏期间褐变程度较低。
2.1.2 粒度和Zeta电位
牛乳中95%的酪蛋白以胶束的形式存在,直径约为40~300 nm。胶束粒径的大小反映了乳蛋白的聚集程度,聚集程度越大胶束粒径越大。Zeta电位是表征胶体分散体系稳定性的重要指标,是对颗粒之间相互排斥或吸引力强度的度量。分子或分散粒子越小,Zeta电位的绝对值(正或负)越高,体系越稳定,即溶解或分散可以抵抗聚集。反之,Zeta电位(正或负)越低,越倾向于凝结或凝聚,即吸引力超过了排斥力,分散被破坏而发生凝结或凝聚。N-UHT和离C-UHT乳在6个月贮藏期内粒度和Zeta电位变化如图2。
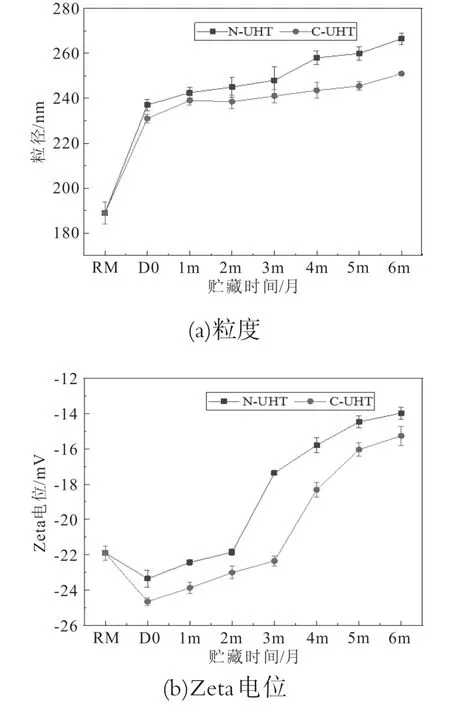
图2 UHT乳贮藏期间酪蛋白胶束粒度和Zeta电位的变化
与原乳比较,两种UHT乳酪蛋白胶束粒径显著增大,随着贮藏时间的延长,酪蛋白胶束粒径逐渐增加。C-UHT乳的粒径显著小于N-UHT乳,并且在贮藏过程中粒径增大的程度也低于N-UHT(图2a)。热处理后乳清蛋白首先变性,变性的乳清蛋白与κ-CN通过二硫键生成稳定的复合物聚集在酪蛋白胶束表面,导致酪蛋白胶束粒径增大[13]。乳蛋白的聚集受乳蛋白质组成、酸度、热处理温度和持续时间、体系内蛋白酶类型及活力等因素的影响,微生物分泌的耐热性蛋白酶是UHT乳贮藏期间不稳定的主要原因之一[14]。Bagliniere等人研究发现,原料乳接种荧光假单胞菌的UHT乳比原料乳不含荧光假单胞菌的UHT乳贮藏期内粒径增幅大[15]。C-UHT乳在原料乳处理过程中除去了大部分微生物,源于微生物菌体的蛋白酶含量显著降低,这也是C-UHT乳贮藏期内粒径低于N-UHT乳的因素之一。
UHT乳的Zeta电位均为负数,说明牛乳是带负电的体系。热处理后两种UHT乳的Zeta电位负值增加,C-UHT的Zeta电位绝对值大于N-UHT(图2b)。随着贮藏时间的增加,两种UHT乳的Zeta电位绝对值逐渐降低,说明乳稳定性下降。N-UHT乳在2个月内Zeta电位变化不显著,C-UHT乳的Zeta电位在3个月内变化不显著,然后两种UHT乳的Zeta电位绝对值显著降低。C-UHT乳的Zeta电位绝对值始终高于N-UHT乳,说明C-UHT的稳定性高于N-UHT。热处理引起蛋白糖基化反应,κ-CN的部分糖基化产生净负电荷,该电荷在酪蛋白胶束之间引起静电排斥[16],导致UHT乳的Zeta电位绝对值提高。O’connell等人研究发现热处理后酪蛋白分子的净负电荷增加[17]。随着贮藏时间的增加,乳体系发生了一系列的物理化学变化,乳糖分解产生有机酸使乳体系的p H值降低,酪蛋白磷酸钙从胶束中溶出影响了体系盐类平衡[18]。在贮藏期间乳蛋白水解并释放出含有负电荷基团的肽,如谷氨酰基,天冬氨酰基,磷酸丝氨酸和糖苷残基等都可以降低胶束的Zeta电位[19]。C-UHT乳由于微生物酶的含量较低,由微生物酶引起的乳体系失稳程度低于N-UHT,体现在离心除菌UHT乳的Zeta电位绝对值高于常规UHT乳。
2.1.3 流变性
黏度系数K反映流体的表观黏度,而n反映流体的剪切能力,n值越大,流体呈现出越强的剪切变稀能力[20]。测定了N-UHT和C-UHT乳在6个月贮藏期内黏度系数K和流动行为指数n变化(图3)。
热处理对乳流动行为指数n的影响不显著,但随着贮藏时间增加,N-UHT和C-UHT的流动行为指数n显著增加,并且C-UHT的n值显著高于N-UHT(图3a)。热处理后,N-UHT和C-UHT乳的黏度系数K增大,并且随着贮藏时间的增加K值呈线性增加,N-UHT的K值显著大于C-UHT(图3b)。乳的K值变大,n值变小,说明其表观黏度变大,剪切变稀能力变强。C-UHT乳的K值始终小于N-UHT乳,n值始终大于N-UHT乳,说明C-UHT乳的黏度增加和变稀能力低于N-UHT乳。
热诱导的变性乳清蛋白结合在酪蛋白胶束表面,增加了胶束粒径,黏度增加。贮藏过程中,酸度增加,胶束粒径增大,蛋白凝聚等因素都会导致K增大。n<1说明乳是剪切变稀型流体,在稳定的剪切流动下,流体黏性随剪切速度的增加而减少。随着贮藏时间的延长呈现出更明显的剪切变稀能力,说明整个乳体系稳定性下降。N-UHT的n值始终小于C-UHT乳,这与贮藏过程蛋白的水解,胶束呈现更加松散结构有关[21]。乳中残余的内源性耐热酶,如组织蛋白酶G,B和D和纤溶蛋白酶是引起蛋白水解的主要酶[22]。UHT乳中残存的微生物蛋白酶(如假单胞菌产生的蛋白酶)在贮藏过程中也会引起蛋白质水解和结构变化,降低乳体系稳定性[23]。C-UHT乳的黏度系数K值高、流动行为指数n较低,表明其在贮藏期维持较好的稳定性。

图3 UHT乳贮藏期黏度系数K和流动行为指数n变化
2.2 贮藏期间UHT乳的化学性质变化
高温杀菌导致乳蛋白变性、乳蛋白氧化、脂质氧化、维生素降解等一系列化学反应,贮藏过程中一些化学反应持续进行[24-25]。贮藏过程发生的变化严重地影响了UHT乳的货架期,并对产品的营养性产生了一定的影响。
2.2.1 酸度和过氧化值
乳酸度是指自然酸度和发酵酸度之和,正常牛乳的酸度是16~18°T。过氧化值是表示脂肪酸被氧化程度的指标。脂肪酸氧化后生成过氧化物、醛、酮等,过氧化值越高说明其酸败和氧化程度越严重。N-UHT乳和C-UHT乳在6个月贮藏期内酸度和过氧化值变化如图4所示。
热处理前后UHT乳酸度变化不显著,N-UHT和C-UHT在3.5个月内酸度变化不显著,N-UHT酸度略高于C-UHT乳,3.5个月以后两者酸度差异不显著,并且随着贮藏时间延长酸度显著增加,整个贮藏期内酸度都低于18°T,符合产品国家质量标准(GB25190-2010,灭菌乳)要求(图4a)。原料乳的酸度是由于微生物分解乳糖产生乳酸,微生物可将一分子乳糖转化为四分子乳酸。UHT乳中一些内源性耐热酶和微生物酶在贮藏中能被激活并发挥作用,乳脂肪和蛋白被水解成脂肪酸和氨基酸,使UHT乳酸度增加。
N-UHT乳在贮藏4.5月时检出过氧化物,C-UHT乳贮藏期5个月时检出过氧化物,贮藏6个月时N-UHT和C-UHT乳的过氧化值高达12.7~12.9 mg/100g,两者差异不显著(图4b)。脂肪氧化产物对贮藏期内的UHT乳风味、色泽、质地、组织等带来不良的影响,降低其营养价值。脂肪氧化发生在原料乳中,并在热处理后脂肪氧化程度增加[26]。脂肪氧化受多种因素的影响,原料乳质量、脂肪酸的组成、脂肪球膜组成和结构、脂肪球膜所带的电荷等[27]。
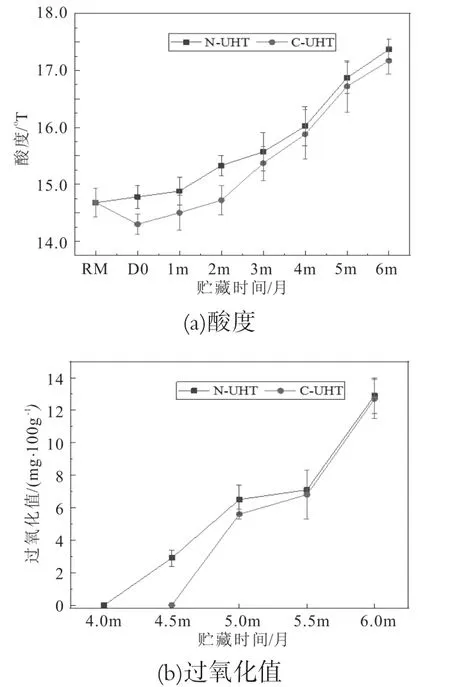
图4 UHT乳贮藏期间酸度和过氧化值的变化
2.2.2 纤溶蛋白酶和非蛋白氮的变化
牛奶中含有少量胰蛋白酶样丝氨酸蛋白酶,负责对酪蛋白的有限水解,乳蛋白的有限水解表现在非蛋白氮(NPN)增加。牛奶蛋白酶被认为是来源于血液,其鉴定为纤溶蛋白酶及其不活跃的纤溶蛋白酶原[28]。纤溶蛋白酶及其酶原纤溶蛋白酶原具有很高的耐热性,UHT杀菌强度不能使得其完全失活,是引起UHT乳货架期内不稳定性的主要因素[29]。研究了N-UHT和C-UHT乳在6个月贮藏期内纤溶蛋白酶活性变化及非蛋白氮含量的变化(图5)。
热处理后UHT乳中纤溶蛋白酶活性显著降低,在贮藏过程中纤溶蛋白酶活性逐渐升高,N-UHT乳的纤溶蛋白酶活性显著高于C-UHT乳(P<0.05),贮藏6个月后N-UHT乳纤溶蛋白酶活性与原料乳相近(图5a)。纤溶蛋白酶耐热性比较强,经高温处理后依旧具有一定的活性[29]。贮藏过程中纤溶蛋白酶活性增加与体细胞数量有关,研究显示体细胞数量与纤溶蛋白酶活性呈正比,体细胞中含有纤溶蛋白酶原激活剂,可增强非活性纤溶蛋白酶原向纤溶蛋白酶的转化,并且高体细胞乳中纤溶蛋白酶原向纤溶蛋白酶的转化率较高[30]。离心除菌技术可以除掉原料乳中大部分体细胞,因此,源于体细胞的纤溶蛋白酶会减少。
非蛋白氮(NPN)是指乳中溶于12%三氯乙酸中可溶性氮的总称,主要是由尿素、尿酸、肌酸、肌酐、多肽、氨基酸、氨等物质组成,约占牛乳总氮的5%~6%[31],NPN是评价乳中真蛋白含量变化非常重要的指标,NPN含量的增加也间接反映了乳蛋白体系的不稳定。热处理导致NPN含量的增加,N-UHT中NPN由原乳的4.2%增加到5.3%,C-UHT中NPN则增加到4.6%,N-UHT中NPN的增加幅度高于C-UHT。随着贮藏时间的增加,NPN含量逐渐增加;贮藏6个月后,NPN从N-UHT-D 0的5.3%增加到N-UHT-6m的8.7%,C-UHT-D0从4.6%增加到C-UHT-6m的7.9%;贮藏过程中C-UHT中NPN的变化程度显著低于N-UHT(P<0.05)(图5b)。热处理后残余的纤溶蛋白酶和细菌蛋白酶引起蛋白水解,产生游离氨基酸、肽以及蛋白降解产物[25]。贮藏过程中,UHT乳中残留耐热酶导致的蛋白水解水平上升,由于离心除菌工艺除掉了原料乳中大部分体细胞和微生物,外源性的细菌蛋白酶,以及体细胞分泌的胞内酶显著降低[32]。Bagliniere等人研究发现,含有200μg/L细菌蛋白酶Ser2的UHT乳贮藏90 d后NPN含量显著高于普通的UHT乳[15]。因此,N-UHT乳中蛋白酶含量高,更大程度的水解乳中蛋白产生含氮的小分子,如尿素,肌酸,小肽和游离氨基酸。此外,热处理和贮藏过程中NPN的增加与热诱导的蛋白降解、脱酰胺作用和美拉德反应的发生有关[24]。
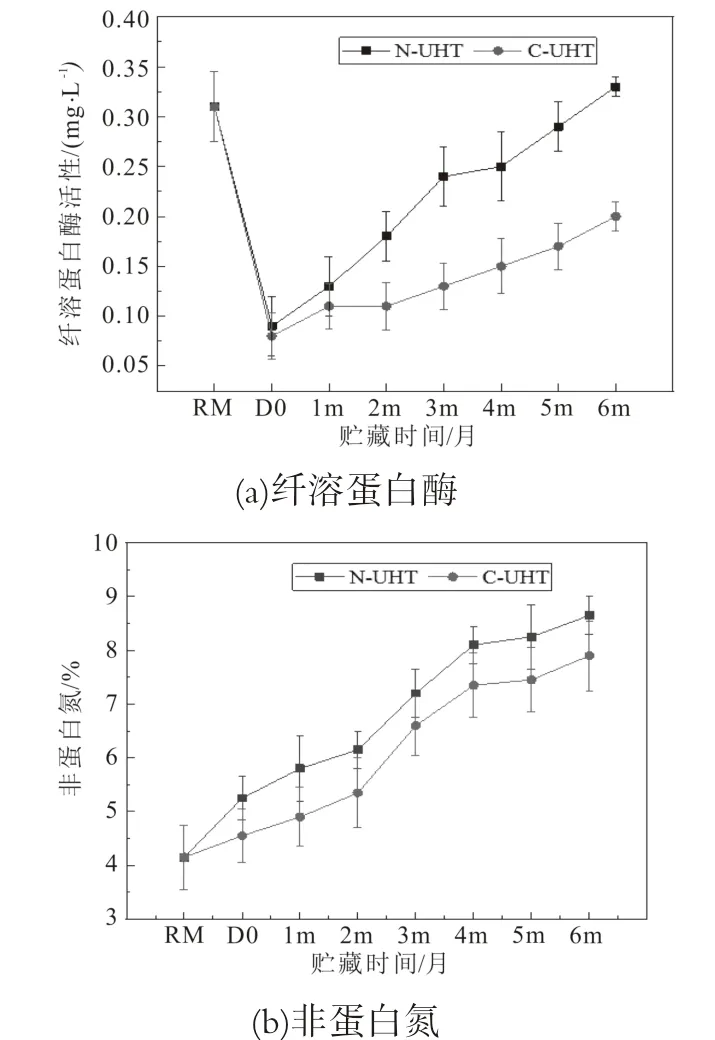
图5 UHT乳贮藏期间纤溶蛋白酶含量变化
2.2.3 贮藏期间乳蛋白组成的变化
乳中总氮(TN)是酪蛋白氮(CN)和乳清蛋白氮(WN)以及非蛋白氮(NPN)之和,CN和WN是真蛋白。正常牛乳中CN、WP、NPN三者保持一定的比例,热处理后乳清蛋白变性,进入胶束相增加了胶束相CN的比例,热诱导的乳蛋白降解增加了NPN的含量。热处理导致乳中氮分布比例发生了改变,影响了乳胶体的稳定性。在贮藏过程中,残余酶导致的蛋白水解、热诱导复合物的产生以及美拉德反应的持续进行,导致了氮分布比例的改变(图6)。
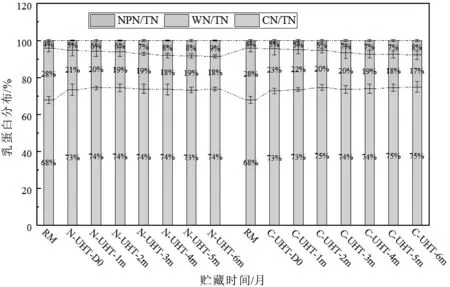
图6 UHT乳热处理和贮藏期氮分布特征
加热和贮藏期乳的总氮(TN)不会变化,但是,CN、WN、NPN三者的含量及占TN的比例能发生改变。与原乳(RM)比较,热处理后N-UHT-D0中WP-N含量由28.1%减少到21.5%,NPN由4.2%增加到5.3%,CN由67.8%增加到73.3%;C-UHT-D0中WN含量由28.1%减少到22.8%,NPN由4.2%增加到4.6%,CN由67.8%增加到72.7%(图6)。乳清蛋白是具有4级结构的球状蛋白,热处理破坏其结构,分子展开,游离巯基暴露出来,通过分子间或分子内二硫键相互作用形成凝聚物,进入胶束相[33]。β-Lg和α-La是乳清蛋白的主要成分,变性β-Lg和α-La与胶束表面的κ-CN通过分子间二硫键作用形成乳清蛋白/κ-CN复合物聚集到酪蛋白胶束表面[34]。热诱导的蛋白降解以及美拉德反应和脱酰胺作用的发生,产生了游离氨基酸、肽、以及一些蛋白降解产物,这些产物以非蛋白氮形式存在[25]。因此,WN含量降低,NPN含量增加。
N-UHT和C-UHT乳在贮藏过程中CN含量变化不显著,两者之间差异不显著。随着贮藏时间的增加WN含量逐渐降低,N-UHT中WN由N-UHTD0的21.5%降低到17.6%(N-UHT-6m),C-UHT的WN由C-UHT-D0的22.8%降低到17.2%(C-UHT-6m),两者之间的变化差异不显著;NPN随着贮藏时间的增加而显著增加,从N-UHT-D 0的5.3%增加到N-UHT-6m的8.7%,C-UHT的NPN从C-UHTD0的4.6%增加到7.9%(C-UHT-6m)。结果表明,在贮藏过程中NPN的增加主要与WN有关。热诱导的蛋白质氧化及蛋白质之间的相互作用在贮藏过程中持续发生,蛋白水解、蛋白的氧化降解、美拉德反应等是引起贮藏期间蛋白含量改变的主要原因[24]。原料乳中嗜冷菌分泌的耐热性蛋白酶和内源性纤溶蛋白酶在UHT贮藏过程中持续水解蛋白,造成UHT乳苦味和凝胶形成已经被大量研究所证实[2]。非酶脱酰胺是将蛋白质的天冬酰胺和谷氨酰胺残基中的酰胺基水解为天冬氨酸和谷氨酸以及氨水,UHT热处理引起的非酶脱酰胺程度较小,但在贮藏过程中能够发生明显的脱酰胺作用[24],并且是随着贮藏时间的延长而增加。UHT杀菌强度引发美拉德反应,贮藏过程中美拉德反应持续进行生成糠氨酸及褐色色素[24],体现在UHT长期贮藏颜色变褐色。蛋白水解产物在乳中最直观的体现是NPN的增加。
3 结 论
UHT乳贮藏期乳蛋白发生了褐变反应,酪蛋白胶束发生了凝聚,牛乳流体的表观黏度变大,剪切变稀能力变强,乳胶体稳定性降低,C-UHT乳的物理性和物理稳定性优于N-UHT乳。随着贮藏时间的增加,UHT乳酸度和纤溶蛋白酶活性增加;贮藏6个月后,N-UHT和C-UHT乳过氧化值高达12.7~12.9 mg/100g,N-UHT乳纤溶蛋白酶活性与原料乳相近,C-UHT乳纤溶蛋白酶活性是N-UHT的60%。贮藏过程中CN含量变化不显著,WN含量逐渐降低,NPN含量逐渐增加;贮藏6个月后,N-UHT和C-UHT的NPN从4.2%分别增加到8.7%和7.9%。热处理和贮藏对N-UHT乳的物理和化学稳定性的影响高于C-UHT乳。离心除菌技术对改善UHT乳的质量具有积极的作用。
