幼苗期摘心对高产春大豆根系生长和产量的影响
2021-04-28楚光红章建新赵占营
楚光红,章建新,王 聪,赵占营
(新疆农业大学农学院,新疆 乌鲁木齐 830052)
幼苗期摘心作为大豆增产的一项有效农艺措施,同时也是一个塑造株型的过程[1-4],根系作为大豆主要的水分和营养吸收器官是产量形成的主要贡献者,根系的时空分布特征决定了植株获得水分和养分的能力,并与其地上部的生长密切相关[5-7]。因此,研究大豆幼苗期摘心后的根系生长特性对大豆高产栽培具有重要意义。以往研究表明,株型是大豆植株整体特征的集中表现,各器官空间结构分布在一定程度上决定了光合产物的分配[8],良好的株型结构将有利于促进大豆群体对光能的吸收利用和群体内的气体交换[9]。分枝数是影响大豆株型的重要因素之一,且与光合效率、结荚特性和抗倒伏性等性状密切相关[10]。强大的根系是作物获得高产的重要保证,强健的根系能够吸收更多的水分和养分,促进地上部叶片的光合作用,增加干物质积累量,而充足的光合产物又将为根系的生长和发育提供必要的营养物质,二者相辅相成[11-14]。董钻等[15]通过对大豆幼苗摘心试验发现,在大豆第一对真叶展开至第一片复叶展开时摘心可促使腋芽形成二主茎、三主茎和四主茎等次生主茎。尚文艳等[16]研究发现,第一复叶期和第二复叶期摘心的产量极显著高于不摘心和真叶期摘心处理,同时第一复叶期摘心和第二复叶期摘心的产量差异不显著(p>0.05),且第一复叶期摘心更有利于形成较多的有效分枝。由此可以看出,摘心改变了大豆株型,改善了群体结构,增加了株叶片数量、单株节数、结荚节数和生殖生长期的干物质积累量,因此实现了最终的增产[17]。然而,大豆摘心后株型结构的改变必然会影响到根系的生长发育,由于根系不可视且研究困难,前人关于大豆摘心后根系的生长变化研究相对较少,特别是对现有较高产量水平下的大豆品种摘心的研究存在不足。鉴于此,本文以主茎型高产春大豆为试验材料,田间研究了大豆幼苗期摘心后根系形态性状和根系活性时空分布特征及产量构成,以期为高产大豆减量播种提供理论依据。
1 材料与方法
1.1 试验材料及试验设计
试验于2018年4—10月在新疆农业大学三坪实习农场进行,试验用地土层深度为0~80 cm,其下为石块。0~20 cm基础肥力为:有机质10.88 mg·kg-1、碱解氮110.0 mg·kg-1、速效磷26.2 mg·kg-1、速效钾328.2 mg·kg-1,土壤pH值 8.17,试验材料选用主茎型高产春大豆品种‘新大豆27号’。
试验一,依据真叶期摘心和复叶期摘心均有利于形成3个分枝而设置真叶期摘心(TP,5月27日)和第一复叶期摘心(CP,6月2日),以不摘心为对照(CK1),共3个处理,理论留苗均为25万株·hm-2(此密度为该地区本品种不摘心栽培最适宜密度),以此探究高产大豆最适合的摘心时间。
试验二,在试验一的基础上设置了5个不同密度处理,分别为15(D1)、18(D2)、21(D3)、25(D4)和30(D5)万株·hm-2,进行第一复叶期摘心,对照为25(CK2)万株·hm-2,以此探究摘心对种植密度的响应规律。
上述2个试验独立设置,小区随机排列,3次重复,行长7 m,每个小区6行,人工摘心时以高于腋芽处0.5 cm为宜(见图1)。
4月28日人工开沟条播,全生育期不覆盖地膜,宽窄行(50 cm+30 cm)种植模式,滴灌毛管铺设在窄行中间,由于播后土壤较为干旱,先滴水1次促进种子萌发出苗,5月15日出苗后定苗。6月5日滴头水,全生育期共滴水7次,总滴水量4 340 m3·hm-2,于始花期、结荚中期分别随水施尿素150 kg·hm-2,另外分别于始花期、始荚期和鼓粒初期各喷施叶面肥1次,9月10—20日不同处理分别成熟,9月20日统一收获。
1.2 测定项目及方法
1.2.1 根系形态特征测定 分别于6月23日(始花期)、7月7日(始荚期)、7月20日(盛荚期)、8月8日(始粒期)、8月25日(鼓粒期)和9月10日(成熟期)进行根系取样,各处理选取代表性样点,自子叶节处剪断植株后依次挖取0~20、20~40 cm和40~60 cm土层根样,取土体积为0.024 m3,40 cm(长,即宽、窄行1/2处间距)×30 cm(宽,即行长方向)×20 cm(高),3次重复。取出的根系用流水冲洗干净,冲洗时在根系下面放置100目筛网,冲完后将主根和侧根分开,用根系扫描分析仪(WinRHIZ0-2004a,Canada)分别扫描各土层侧根,用万深LA-S植物根系分析系统对获得的图像进行分析并获得侧根长、侧根表面积和侧根体积数据。将主根和扫描后的侧根在105℃下杀青30 min后于80℃烘箱中烘至恒重,使用万分之一天平称重并计算侧根干质量密度、侧根长密度和侧根表面积密度。
1.2.2 根系活力测定 采用TTC氧化还原法,准确称取较细的鲜根样0.5 g放入三角瓶中,之后加入含有0.4%TTC溶液和磷酸缓冲液各5 ml的混合液,用封口膜密封瓶口后置于37℃的暗室中反应2 h,之后快速加入浓度为1 mol·mL-1的硫酸2 mL终止反应,用镊子将根取出,用吸水纸轻轻擦拭至根表面水分干燥后放入研钵中,加乙酸乙酯3~4 mL充分研磨,将红色提取液移入刻度试管,用乙酸乙酯冲洗数次并定容至10 mL,用分光光度计在波长485 nm下比色,以空白试验(将根先用硫酸处理)作参比测出吸光度,后查标准曲线即可求出TTC还原量。根系活力计算公式如下:
根系活力(mg·g-1·h-1)=C/(1000W·t)
式中,C为四氮唑还原量(μg);W为根鲜质量(g);t为反应时间(h)。
1.2.3 考种及测产 成熟期(9月10—20日)各小区收取中间4行(1.6 m×3 m = 4.8 m2)植株,人工脱粒称重,随机称取100 g籽粒于80℃烘干至恒重计算含水量,最终折合成含水量13.5%时的产量。另选取具有代表性的连续植株20株于室内进行考种,测定单株荚数、单株粒数、单株腔数和百粒重。
1.3 数据统计与分析
使用Excel 2013、SSPS 19.0和Origin 2018软件进行数据处理、统计分析和图表绘制。
2 结果与分析
2.1 幼苗期摘心对大豆总侧根干质量及侧根干质量密度垂直分布的影响
试验一中(图2A),TP、CP总侧根干质量在6月23日(始花期)显著低于对照(CK1),分别增加了42.62%和10.29%;7月7日,CP高于对照,TP低于对照;7月20日(盛荚期)总侧根干质量达到峰值,此时CP显著高于对照,增加了18.66%,而TP显著低于对照,降低了7.33%;8月25日(鼓粒期)-9月10日(成熟期)TP、CP总侧根干质量的下降速率明显快于对照,且成熟期时CP与对照差异不显著;CP的0~20、20~40 cm侧根干质量密度峰值(7月20日)分别较对照高出24.52%和38.86%,而TP的0~20、20~40 cm侧根干质量密度峰值与对照差异不显著,但TP和CP的40~60 cm侧根干质量密度生育期内均显著低于对照。试验二中(图2B),不同密度摘心处理总侧根干质量在6月23日均显著低于对照(CK2),7月20日(峰值)处理间表现为D3、D4、D2、D5>D1、CK2;各摘心处理0~20 cm侧根干质量密度峰值(7月20日)均显著高于对照,20~40 cm侧根干质量峰值(7月20日)除D1外均显著高于对照,而40~60 cm侧根干质量密度峰值(8月8日)均显著低于对照。两试验结果表明,幼苗期(第一复叶期)摘心先抑制根系生长,之后促进根系生长,增加总侧根干质量,增加0~20、20~40 cm侧根量,降低40~60 cm侧根量,适当降低摘心密度(D3~D2)大豆群体侧根量依然增加。
2.2 幼苗期摘心对大豆总侧根长度及侧根长密度垂直分布的影响
试验一中(图3A),摘心后TP总侧根长度在生育期内均显著低于对照(CK1),其峰值(7月20日)低于对照6.83%;CP总侧根长在7月7日前显著低于对照,之后显著高于对照,8月8日(峰值)较对照高7.35%,但成熟期时CP与对照差异不显著;CP的0~20、20~40 cm侧根长密度峰值(7月20日)显著高于对照14.14%、14.94%,而TP的0~20、20~40 cm侧根长密度峰值与对照无显著差异,但二者40~60 cm侧根长密度生育期内均显著低于对照;试验二中(图3B),不同密度摘心处理总侧根长在6月23日显著低于对照(CK2),之后除D1外均高于对照,且以D3处理数值最大,7月20日(峰值)处理间表现为D3、D4、D2、D5>D1、CK2;摘心后0~20 cm、20~40 cm侧根长密度峰值(7月20日)除D1外,均表现为显著高于对照,而40~60 cm侧根长密度峰值(7月20日)均表现为低于对照。两试验结果表明,幼苗期(第一复叶期)摘心先抑制根系伸长,之后促进根系伸长,增加总侧根系长度,增加0~20、20~40 cm侧根长密度,降低40~60 cm侧根长密度,适当降低摘心密度(D3~D2),群体总侧根长度依然增加。
2.3 幼苗期摘心对大豆总侧根表面积及侧根表面积密度垂直分布的影响
试验一中(图4A),摘心后TP、CP总侧根表面积在6月23日分别显著低于对照(CK1)40.86%和20.43%,之后二者快速增加,7月20日(峰值)TP与对照差异不显著,而CP显著高于对照17.49%; TP、CP处理的0~20 cm侧根表面积密度分别较对照增加了10.04%和25.26%,CP处理的20~40 cm侧根表面积密度较对照高18.44%,而TP处理的20~40 cm侧根表面积密度较对照下降了10.13%,但二者的40~60 cm侧根表面积密度自7月7日后均显著低于对照。试验二中(图4B),不同密度摘心处理总侧根表面积在6月23日均显著低于对照(CK2),7月7日除D5外均低于对照;7月20日总侧根表面积达到峰值,且以D3数值最大,处理间表现为D3、D4、D2、D5、CK2>D1;摘心后0~20 cm和20~40 cm侧根表面积密度峰值(7月20日)除D1处理外均高于对照,而40~60 cm均低于对照。两试验结果表明,幼苗期(第一复叶期)摘心先抑制侧根系表面积增加,之后促进侧根系表面积增加;摘心增加总侧根系表面积,增加0~20、20~40 cm侧根表面积密度,降低40~60 cm侧根表面积密度,适当降低摘心密度(D3~D2),群体根系表面积依然增加,增加根系表面积有利于提高根系对水分和养分的吸收量。
2.4 幼苗期摘心对大豆侧根根系活力的影响
试验一中(图5A),TP、CP的0~20、20~40 cm和40~60 cm根系活力在6月23日表现为显著低于对照(CK1),至8月25日CP处理各层根系活力均表现为显著高于对照,而TP处理各层根系活力依然表现为大多低于对照;7月20日各层根系活力达到最大值,此时CP的0~20、20~40 cm和40~60 cm根系活力较对照分别高出14.00%、15.43%和15.41%,但是峰值过后其各层根系活力的下降速率明显快于对照而表现为成熟期(9月10日)与对照差异不显著,但是TP的0~20、20~40 cm和40~60 cm根系活力较对照分别下降8.46%、5.03%和4.82%。试验二中(图5B),不同密度摘心处理0~20 cm和20~40 cm土层的根系活力在6月23日表现为显著低于对照(CK2),40~60 cm土层的根系活力则高于对照,其中D1~D3与对照差异显著。7月7日后各摘心处理根系活力快速提高。随摘心密度增加根系活力呈先升后降变化,且以D2处理最高,但与D1、D3多表现为差异不显著。两试验结果表明,摘心后0~60 cm土层中各层根系活力短期内先降低后提高,适当降低摘心密度(D3~D2)有利于提高大豆根系的吸收能力。
2.5 幼苗期摘心对大豆农艺性状的影响
由表1可知,摘心后大豆生育周期推迟近一周,TP、CP株高、茎粗和单茎节数较对照(CK1)显著降低,而收获茎数、单株茎数、单株结荚节数和节间长度显著增加,TP、CP收获茎数、单株茎数和单株结荚节数分别较对照增加98.27%、97.66%,93.00%、92.00%和34.92%、31.22%;降低摘心密度(D3~D1)株高、单茎节数和节间长不变,而单株茎数、单株节数显著增加。

表1 幼苗期摘心对大豆农艺性状的影响
2.6 幼苗期摘心对大豆产量及其构成因素的影响
由表2可知,摘心后TP、CP单节荚数、单株荚数和单株粒数较分别较对照(CK1)下降了40.26%、34.29%,19.32%、14.00%和17.42%、14.02%,百粒重分别增加了12.25%、10.08%,籽粒产量分别下降了5.16%、2.64%(p>0.05),CP减产幅度小于TP;降低摘心密度(D3~D1)单节荚数、单株荚数和单株粒数显著增加,增加摘心密度(D5)则显著降低;群体荚数和群体粒数以D2最高,但与对照(CK2)差异不显著,而其籽粒产量显著高于对照5.90%;百粒重除D5外均显著增加;指标间的相关分析表明,产量只与根干质量呈正相关(r=0.018)但不显著,而产量与单株荚数、单株粒数、群体荚数、群体粒数和百粒重的相关系数分别为r1=0.884*、r2=0.875、r3=0.811、r4=0.994**和r5=0.977**(*、**分别表示在α=0.05和α=0.01水平上差异显著)。两试验结果说明,第一复叶期摘心CP群体荚数和群体粒数显著降低而产量不降低的主要原因是百粒重增加补偿荚粒数减少的结果,而D2收获株数显著降低而产量显著增加的主要原因是分枝数显著增加导致单株荚粒数、群体粒数和百粒重较对照显著增加的结果。
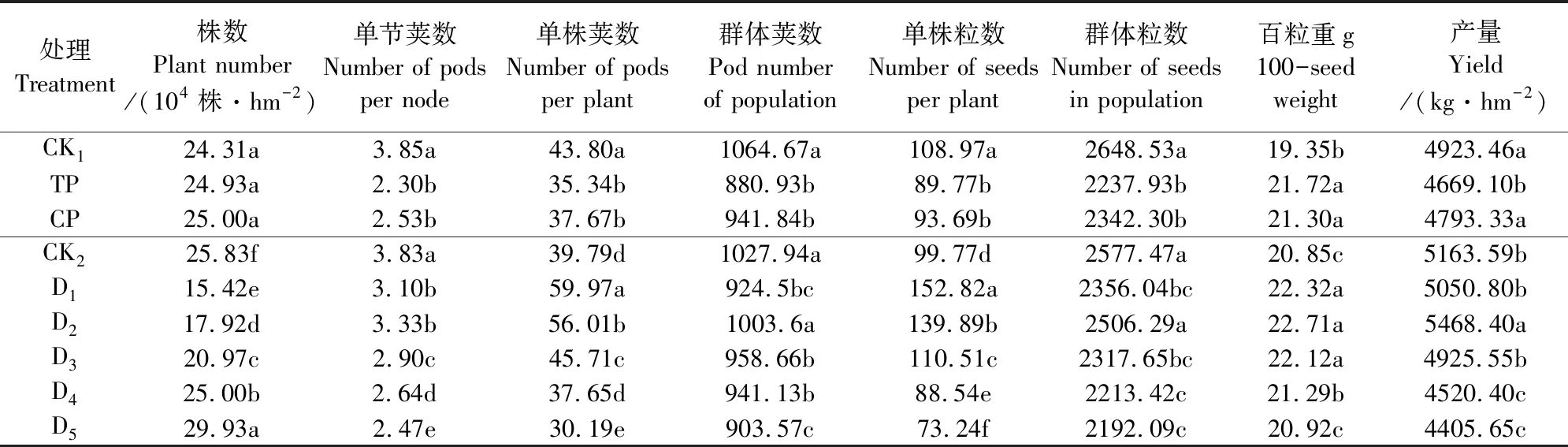
表2 幼苗期摘心对大豆产量及其构成因素的影响
3 讨 论
根系形态性状和根系活力是衡量作物根系对土壤中养分和水分吸收能力的重要指标[18],根系所占据的土壤空间大小和分布状况在很大程度上影响其所获得资源的多少[19-20]。而不同育成年代及不同产量水平结构下的大豆品种根系存在较大差异,近现代大豆品种的根系较当代大豆品种的根系更发达,高产型大豆品种的根系较中、低产大豆品种的根系更发达[21-25]。卢思慧等[1,26]对夏大豆摘心研究表明,夏大豆子叶节摘心形成双茎后对花期以前根系数量增加有一定的抑制作用,但随着地上部的生长和双茎株光合面积增加较多,到荚期以后双茎株根系的数量显著增加,荚期和鼓粒期分别增加19.3%和17.9%,同时第一复叶期摘心和第二复叶期摘心处理分别增产3.2%和5.2%。这与本试验中CP处理相一致,高产型大豆复叶期摘心CP的根量在6月23日(始花期)显著低于对照10.29%,而后在7月20日(盛荚期,峰值)前后又显著高于对照18.66%,这说明摘心后随着分枝的不断生长,根系的生长量也在同步增加,而生殖生长期根系的增加有利于大豆鼓粒和产量提高。但本试验中真叶期摘心处理TP根量生育期内均显著低于对照,这可能与摘心前根系、茎秆、真叶和复叶的生理生化特性以及分枝形成位置有关,有待进一步研究分析。同时本试验结果还显示,摘心处理生育后期根系衰老速度明显快于对照,这可能因为地上形成的较多分枝引起群体密度过大,进而加剧了根系对土壤空间、水分和养分的竞争。
产量的形成是作物在生长发育过程中环境条件对其遗传本性满足程度的综合表现,单位面积内有效分枝数量和生长状况是构成产量的重要因素,且产量补偿结果往往来自于分枝发育增加的荚果数[27],而合理密植是大田栽培技术的关键,一方面通过加大群体的种植密度来实现,而另一方面在适度密度的基础上通过增加个体的有效分枝数量达到高产目的,分枝数增加,单株产量也随之增加[28]。本研究通过对高产春大豆幼苗期进行摘心,较早消除了顶端优势,促进腋芽形成较多分枝,新形成的分枝茎数达原来的近2倍。在最适宜栽培密度下真叶期摘心和复叶期摘心处理TP、CP群体荚数、群体粒数和籽粒产量均表现为下降。而降低摘心密度(D3~D1)单节荚数、单株荚数和单株粒数显著增加,增加摘心密度(D5)则显著降低,随收获茎数增加籽粒产量先增加后降低,而当摘心密度下降30.62%时,籽粒产量增加5.90%,这说明在最适密度条件下摘心虽然可以促进根系生长,形成较多分枝茎数,但分枝茎数的过多增加并不利于大豆产量的形成(r=-0.822),当降低摘心密度时籽粒产量显著高于最适密度条件的产量,这说明主茎型高产春大豆幼苗期摘心后形成的多分枝对产量具有补偿或超补偿效应。然而大豆产量的形成是开花、结荚的结果,同时还受到外部条件(土壤、气候、管理等)的综合影响,因此,对于主茎型高产春大豆摘心与增产关系的研究还需进一步深入。
4 结 论
第一复叶期摘心显著促进主茎型高产春大豆侧根干质量、侧根长度和侧根表面积的提高,且其效果优于真叶期摘心;降低摘心密度使根系根量增加,且单株荚数和单株粒数显著增加,当摘心密度降低30.62%时籽粒产量增加5.90%,此时群体荚数和群体粒数仅分别下降了2.37%和2.76%,而百粒重和产量分别提高了8.92%和5.90%。由此可见,摘心密度显著降低后群体荚粒数和产量不降低的主要原因是摘心显著增加了植株的根量、单株荚数(粒数)和百粒重。
