CO2浓度升高对半干旱区春小麦光合作用及水分生理生态特性的影响
2021-04-12张凯张勃王润元王鹤龄赵鸿赵福年齐月陈斐
张凯 ,张勃,王润元,王鹤龄,赵鸿,赵福年,齐月,陈斐
1. 西北师范大学地理与环境科学学院,甘肃 兰州 730070;2. 中国气象局兰州干旱气象研究所/甘肃省干旱气候变化与减灾重点实验室/中国气象局干旱气候变化与减灾重点开放实验室,甘肃 兰州 730020;3. 中国气象局定西干旱气象与生态环境野外科学试验基地,甘肃 定西 743000
大气 CO2浓度升高已成为世界范围内重要环境问题。由于煤炭、石油等化石燃料的过度使用以及人类对植被生态系统的不合理利用,使得全球范围内大气环境CO2浓度显著升高。政府间气候变化专门委员会(IPCC)在第五次评估报告中指出,大气CO2浓度(摩尔分数)已经从1960年西方工业革命开始时的 310 μmol·mol-1升高至当前的 400 μmol·mol-1(IPCC,2013),且以每年约2 μmol·mol-1的速度在增加,预计21世纪末大气CO2浓度将达到 800 μmol·mol-1(Yasuor et al.,2017)。CO2是作物生境的重要因子,是绿色植物光合作用的反应底物,在作物光合作用中起着至关重要的作用。绿色植物通过光合作用将大气 CO2转化为储存能量的有机物质,提供给人类生存所必须的粮食、油料和纤维等(潘瑞炽,2001),大气CO2浓度增加势必会对作物的光合作用与生长发育产生影响。近年来大气中 CO2浓度的升高及其影响成为国内外生态和农业领域的热点研究问题(张凯等,2017)。
关于 CO2浓度升高对作物的影响研究,基本上都是通过控制试验进行的。试验装置最常见的有开顶式气室(Open-Top Chamber,OTC)、开放式空气CO2浓度增高系统(Free-air CO2enrichment,FACE)、人工气候室和同化箱等(房世波等,2010;刘超等,2018)。从目前试验研究情况来看,OTC被广泛用于牧草和水稻、玉米等作物的研究,而对于小麦,前人多采用FACE平台(杨连新等,2007)、人工气候室(康绍忠等,1999)和同化箱(王修兰等,1996)等装置进行研究。
小麦是世界上广泛种植的粮食作物,也是我国干旱半干旱地区的最主要粮食作物,在国家粮食安全和社会经济发展中占有举足轻重的地位。中国北方是春小麦的主要种植区。西北地区是我国北方春小麦的主要产区之一,春小麦种植面积约占全国小麦种植面积的20%,占当地耕地面积和粮食作物播种面积分别为40%和51%(吴宏亮等,2013),占有十分重要的地位。春小麦的生长不仅受到自身遗传物质的控制,同时也受到诸多环境因子的影响(张凯等,2018),包括大气中 CO2浓度增加。自 20世纪 70年代以来,国内外学者已经在小麦对 CO2浓度升高的响应方面做过许多相关研究(康绍忠等,1996;Morison,1985),由于所处的研究区不同以及所采用的试验平台和作物品种不一样,在作用程度大小以及机理响应方面的研究结果还不尽一致。例如关于光合生理特性的研究,有的研究认为 CO2的有限升高确实对包括小麦在内的农作物光合作用有一定的促进作用,导致净光合速率增加(郭丽丽等,2019),有的研究则认为长时间高浓度CO2浓度下的生长会导致小麦呼吸增强,净光合速率的降低,出现光合作用对高浓度CO2的适应或下调,即发生了CO2驯化现象(范金杰等,2020);关于水分生理特性的研究,有研究认为CO2的升高导致叶片水平 WUE的提高是净光合速率提高和蒸腾速率降低共同作用的结果(Li et al.,2017;刘月岩等,2013;郑云普等,2019),也有的研究发现CO2升高时净光合速率并未增加,WUE的提高仅是由于蒸腾速率降低造成的(Prior et al.,2011),而有的研究认为CO2升高引起的WUE的提高是由于净光合速率增大所致,而蒸腾速率的降低并不明显(Kimball et al.,2002)。这些不尽一致的研究结果说明CO2浓度增加对小麦光合和水分生理生态特征的影响是一个复杂的过程,有必要通过不同的试验设备和更多的田间模拟试验进行深入系统的研究。
本文采用国际上通用的 OTC试验平台,进一步探索与分析 CO2浓度升高条件下西北半干旱区春小麦的光合和水分生理生态特征的动态变化过程及其影响机理,为未来CO2浓度升高条件下春小麦生产、水资源的高效利用提供理论指导,为深入理解小麦生长过程对全球 CO2升高的反应效应提供参考,而且为农田生态系统气候变化适应性对策的制定提供理论依据。
1 材料与方法
1.1 试验地点
本试验在中国气象局兰州干旱气象研究所定西半干旱气象与生态环境试验基地(104°37′E,35°35′N)进行。该基地位于甘肃省中部(图1),属典型黄土高原半干旱气候区,地处 104°37′E,35°35′N,海拔高度1896.7 m,年平均气温6.7 ℃,年降雨量386.6 mm,且分布不均匀,主要集中在6—8月。年太阳总辐射为5923.8 MJ·m-2,年日照时数2433.0 h,无霜期平均为140 d。土壤为黄绵土,碱性,肥力中等,其中土壤pH值7.8,有机质含量110.7 g·kg-1,有效氮、总氮分别为 51.1 mg·kg-1和0.84 g·kg-1,有效磷、总磷分别为 14.12 mg·kg-1和1.24 g·kg-1(张凯等,2015)。
图1 试验点在甘肃省的位置示意图Fig. 1 Location of test site in Gansu Province
1.2 试验设计
试验春小麦品种为“定西24号”,叶色灰绿,叶片狭长,茎秆细而有弹性,生育期约为120 d左右,为当地主栽品种。播种日期为2016年4月1日,行距0.15 m,播种量为225 kg·hm-2。春小麦生产管理按照当地高产栽培技术规程进行,其中基肥按农家肥 56000 kg·hm-2、磷酸二铵 228 kg·hm-2和尿素 138 kg·hm-2施用。试验过程中水肥条件适宜且一致。
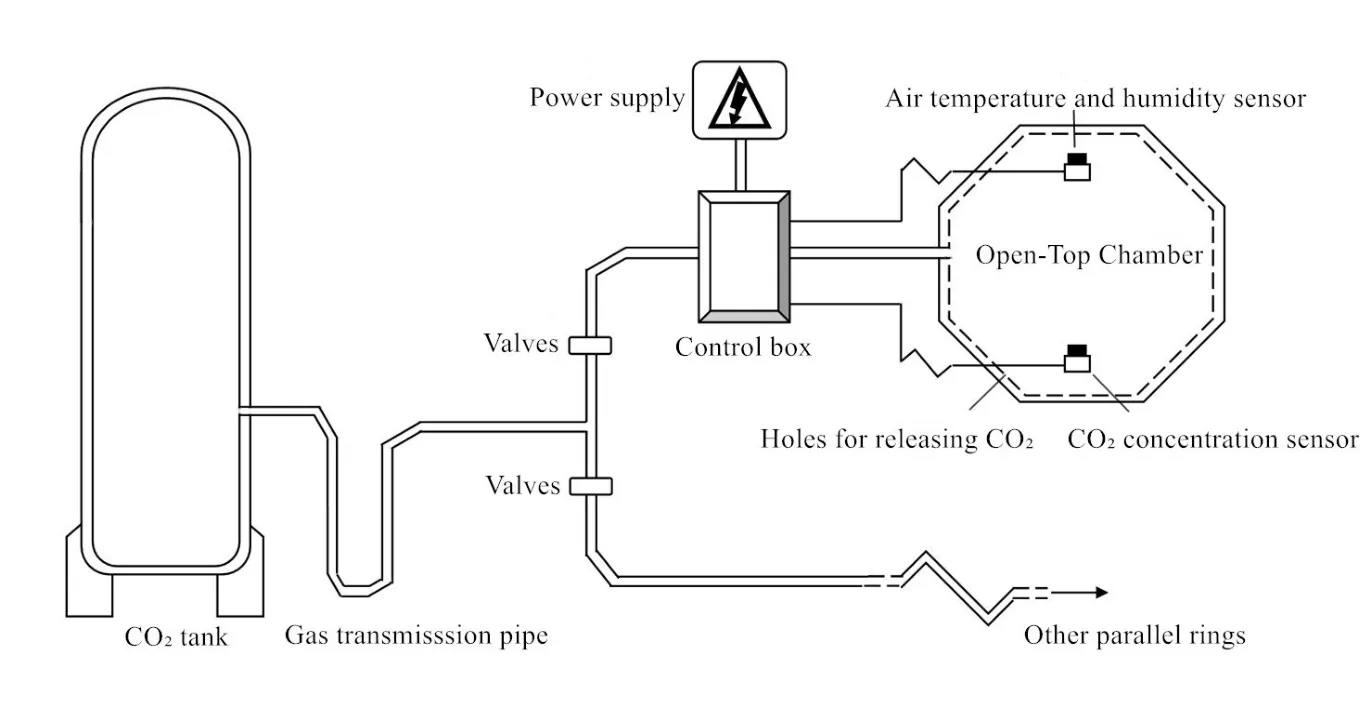
图2 定西OTC田间试验平台主要结构Fig. 2 Main structures of Dingxi cropland OTC experimental platform
试验装置采用OTC系统试验平台(图2),主要由CO2气体供应装置、控制系统、释放系统3大部分组成。试验有3个开顶式气室(边长2.15 m,高2.4 m,玻璃室壁,正八边形),包括2个试验气室和1个对照气室。室内对应由8根不锈钢管围成八边形,不锈钢管面向室内一面每隔100 mm有孔径约0.5 mm的小孔,用以释放纯CO2气体。室内装有1个CO2气体监测器,用于采集CO2气样供控制系统分析圈内CO2浓度分布。此外,还装有温度传感器,实时采集开顶气室内温度数据。开顶式气室通CO2时间从5月初(三叶期)开始到7月底(收获期)结束。根据对未来CO2升高预测的结果以及前人采用的CO2浓度(杨连新等,2007),设计3个开顶式气室分别在对照大气CO2浓度(摩尔分数),在自然条件下该试验地 CO2平均浓度为390 μmol·mol-1的基础上增加 90、180 μmol·mol-1,即 OTC1的浓度为 390 μmol·mol-1(作为对照),OTC2 的浓度为 480 μmol·mol-1,OTC3 的浓度为570 μmol·mol-1。从试验情况来看,各 OTC 内的 CO2浓度基本能达到设计水平,OTC1、OTC2和OTC3内整个生育期实测CO2浓度日平均值分别为389、485、574 μmol·mol-1(图3)。
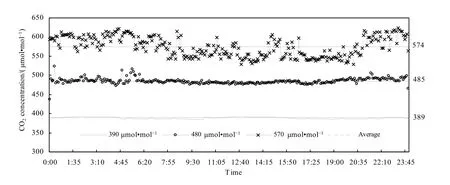
图3 不同OTC内整个生育期日平均CO2浓度Fig. 3 Daily average CO2 concentration during different growth periods in different OTC
1.3 观测项目与方法
采用温度探头自动观测空气温度;按照《农业气象观测规范》(中国气象局,1993),观测不同CO2浓度处理下春小麦的生育期,准确记录春小麦播种、进入出苗、三叶、拔节、孕穗、抽穗、开花、灌浆、乳熟、成熟的时间,以及三叶期、拔节期、孕穗期、抽穗期、开花期、灌浆期、乳熟期的生育时期天数;采用 LI-6400型便携式光合测定仪(LI-COR,USA),分别在春小麦拔节期、孕穗期、开花期、灌浆期和乳熟期选气候条件稳定的晴天,选取生长一致且受光方向相同的上部功能叶10片,测定各处理生长环境和对照环境中春小麦叶片光合的生育期变化和日变化。测定参数包括叶片净光合速率(Pn)、气孔导度(Cs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)和叶温(Tl)。日变化观测从08:00开始,18:00结束,每间隔2 h观测1次。与光合测定系统配套的2 cm×3 cm标准气室可以独立控制光合光量子通量密度(PPFD)、CO2浓度和叶片温度,测量时标准气室参数设定叶室内PPFD为1000 μmol·m-2·s-1PAR , CO2浓 度 为 390 μmol·mol-1,叶片温度25 ℃;采用WP4水势仪在春小麦生长关键期-拔节期对叶片水势日变化进行观测,每隔2 h采样1次,采下样品后立即将叶片剪碎,覆盖水势仪样品杯底部,测定叶片水势。
1.4 水分利用效率的计算
作物的水分利用效率(WUE)反映了 CO2同化作用和水分消耗的关系,通常用瞬时净光合速率与蒸腾速率的比值来表示(王振华等,2015)。

1.5 数据处理
采用Excel 2013软件对数据进行统计分析,利用SPSS 18.0软件的单因素方差分析法(One-way ANOVA)和最小显著差数法(LSD)进行方差分析和差异显著性检验(α=0.05)。
2 结果与分析
2.1 CO2浓度增加对春小麦叶片净光合速率的影响
图4是不同大气CO2浓度下春小麦叶片净光合速率日变化特征。试验结果表明,随着CO2浓度的增加,小麦叶片净光合速率明显提高。在对照大气CO2浓度和增加CO2浓度条件下,小麦叶片净光合速率日变化均呈“双峰型”分布,日出后净光合速率逐渐上升,至12:00达到全天最高值,随后Pn呈下降趋势,16:00出现第二峰值,出现明显的“光合午休”现象。从图4可以看出,观测日上午的Pn明显大于下午,这主要由于经上午光合作用后,叶片中的光合产物有积累而发生反馈抑制的缘故。对不同CO2浓度下小麦叶片净光合速率的日变化做进一步分析,可以发现,与对照相比,480 μmol·mol-1浓度和 570 μmol·mol-1浓度处理下,净光合速率分别平均提高了18.74%和34.86%;两种高CO2浓度条件下春小麦叶片净光合速率的提高,均表现为中午前后最为显著,在早晨和傍晚增强作用相对较小。
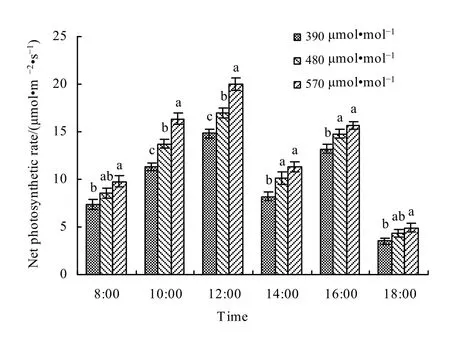
图4 不同大气CO2浓度下春小麦叶片净光合速率日变化Fig. 4 The diurnal variation of net photosynthetic rate in spring wheat leaves under different CO2 levels
图5为春小麦不同生育期叶片净光合速率对大气 CO2增加的响应。可以看出,在对照大气 CO2浓度和增加CO2浓度条件下,净光合速率均表现为开花期最大,乳熟期最小。与对照CO2浓度(390 μmol·mol-1)相比,在 480 μmol·mol-1浓度下,叶片净光合速率在拔节、孕穗、开花、灌浆和乳熟期分别增加了 21.59%、15.65%、7.91%、12.74%和15.50%,平均增加了 14.68%;在 570 μmol·mol-1浓度下,拔节、孕穗、开花、灌浆和乳熟期分别增加了47.19%、28.35%、13.83%、19.24%和32.41%,平均增加了28.20%。
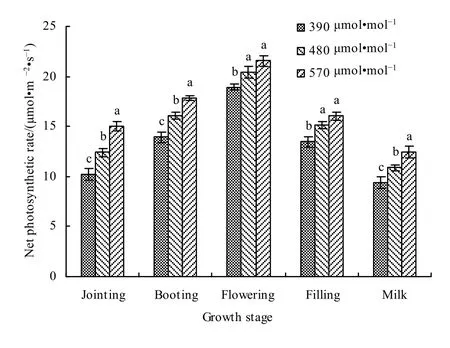
图5 大气CO2增加对不同生育期春小麦叶片净光合速率影响Fig. 5 Effects of different CO2 concentrations on the net photosynthetic rate of spring wheat leaves at different growth stages
分析表明,叶片温度(Tl)与净光合速率的趋势一致,且叶温较高时净光合速率也随之变高,叶温降低时净光合速率也随之降低,具有明显的正相关关系(图6)。由于光合作用过程中的暗反应是由酶所催化的化学反应,而温度直接影响酶的活性,因此,温度对光合作用的影响很大(潘瑞炽,2001),尤其是在高CO2浓度时温度对净光合速率的影响要比低CO2浓度时大。
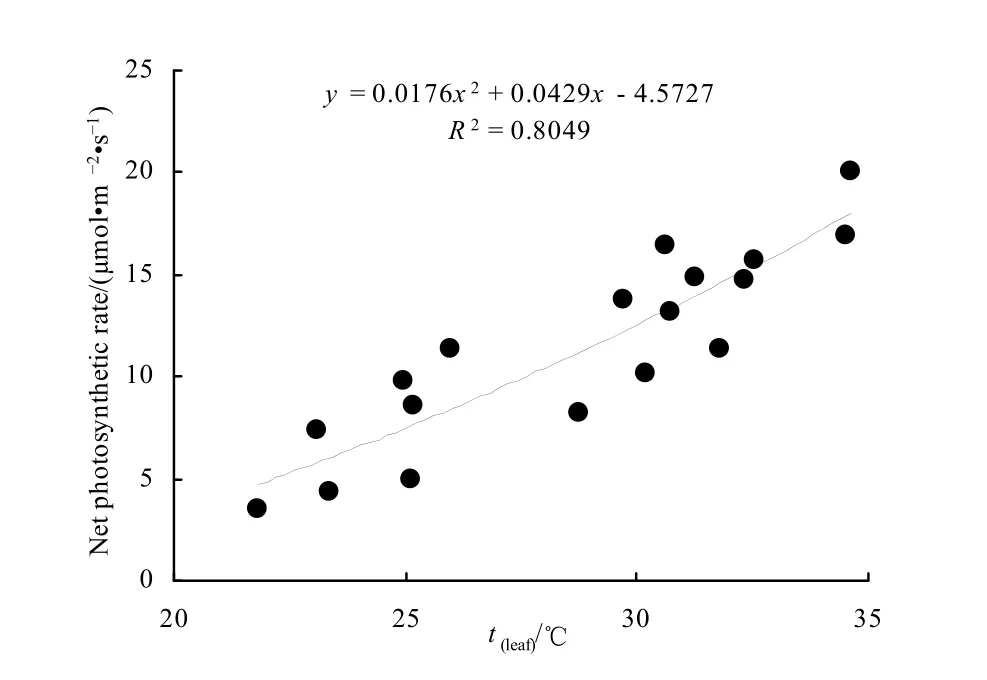
图6 叶温和净光合速率相关关系Fig. 6 Relationship between leaf temperature and net photosynthetic rate
2.2 CO2浓度对春小麦叶片气孔导度的影响
气孔导度(Cs)是衡量气体通过气孔的难易程度,气孔导度越大则气孔开度越大,即气孔阻力小,说明水汽、CO2等可顺利通过气孔进行交换(赵鸿等,2007)。从不同大气CO2浓度下小麦叶片气孔导度日变化可以看出(图7),日出后,随着光照增强,气孔导度增大,08:00达到最大值,之后逐渐减小,到14:00以后,气孔导度又呈上升趋势,16:00出现第二个峰值,随后减小至全天最低。对不同 CO2浓度下小麦气孔导度的日变化做进一步分析表明,与对照相比,480 μmol·mol-1浓度和570 μmol·mol-1浓度处理下,气孔导度分别平均降低了21.05%和36.28%。
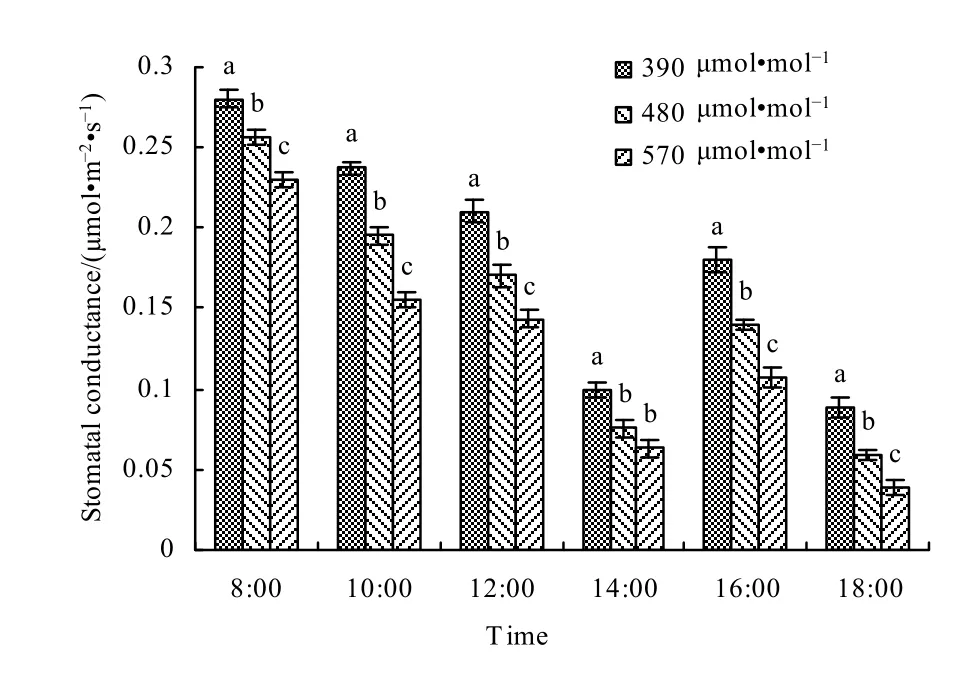
图7 不同大气CO2浓度下春小麦叶片气孔导度日变化Fig. 7 The diurnal variation of stomatal conductance in spring wheat leaves under different CO2 levels
图8为春小麦不同发育期叶片气孔导度对大气CO2增加的响应。可以看出,在对照大气CO2浓度和增加CO2浓度条件下,气孔导度均表现为开花期最大,乳熟期最小。与对照 CO2浓度(390 μmol·mol-1)相比,在 480 μmol·mol-1浓度下,叶片气孔导度在拔节、孕穗、开花、灌浆和乳熟期分别降低了 16.77%、17.22%、18.48%、12.96%和11.02%,平均降低了 15.29%;在 570 μmol·mol-1浓度下,拔节、孕穗、开花、灌浆和乳熟期分别降低了23.87%、31.11%、27.35%、23.15%和18.64%,平均降低了24.83%。

图8 大气CO2增加对不同生育期春小麦叶片气孔导度影响Fig. 8 Effects of different CO2 concentrations on the stomatal conductance of spring wheat leaves at different growth stages
分析表明,气孔导度与空气湿度(RH)的趋势一致,气孔导度随着空气湿度的下降而减小,具有明显的正相关关系(图9)。由于空气湿度与气温直接影响空气的蒸腾能力,因此与气孔导度有较强的关系(司建华等,2008)。
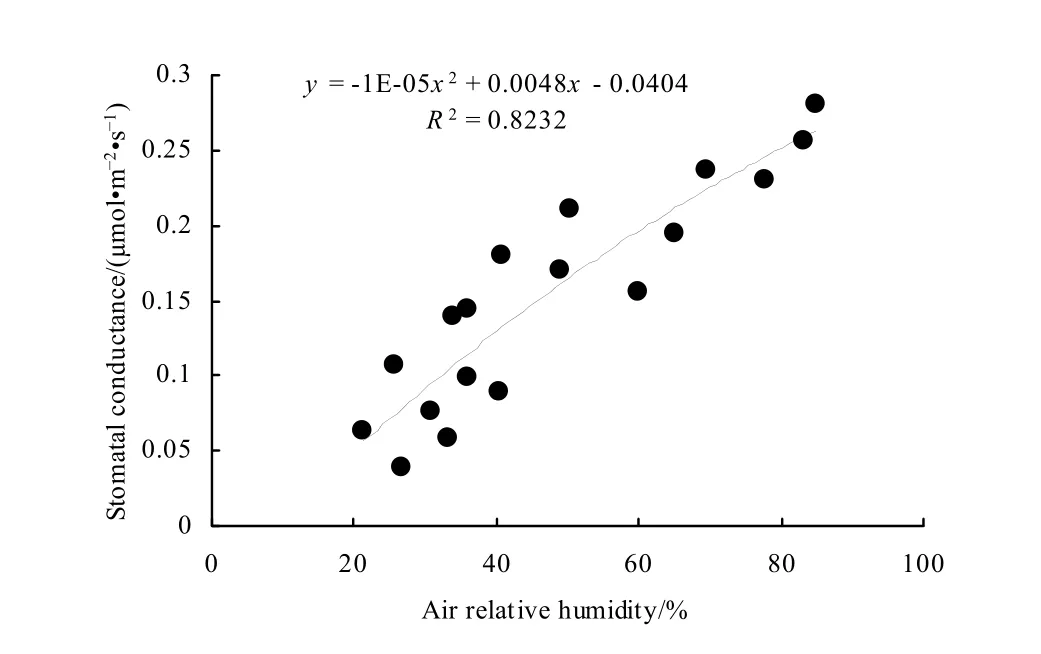
图9 空气相对湿度和气孔导度相关关系Fig. 9 Relationship between air relative humidity and stomatal conductance
2.3 CO2浓度对春小麦叶片胞间CO2浓度的影响
胞间CO2浓度是光合作用的主要原料之一,与净光合速率关系密切,当净光合速率较高时,固定较多的CO2,引起胞间CO2浓度下降(张治安等,2006)。大气CO2浓度的变化也会影响到叶片胞间CO2浓度,大气 CO2浓度增加导致叶片内外 CO2浓度差增大(申双和等,2009)。从图10可以看出,春小麦叶片胞间CO2浓度的日变化特点表现出斜“V”字型曲线。08:00—12:00胞间CO2浓度快速下降,12:00达到全天最低值,12:00之后开始回升。进一步分析不同CO2浓度下小麦叶片胞间CO2浓度的日变化,可以看出,两个增加CO2浓度处理的胞间CO2浓度均高于对照,其中480 μmol·mol-1浓度与 390 μmol·mol-1浓度相比,胞间 CO2浓度平均提高了 9.93%;570 μmol·mol-1浓度与 390 μmol·mol-1浓度相比,胞间 CO2浓度平均提高了33.84%。
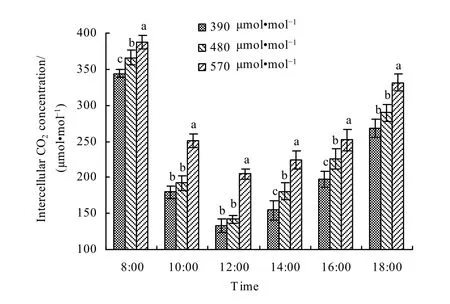
图10 不同大气CO2浓度下春小麦叶片胞间CO2浓度日变化Fig. 10 The diurnal variation of internal CO2 concentration in spring wheat leaves under different CO2 levels
对不同生育期大气 CO2增加对春小麦叶片胞间CO2浓度的影响进行分析(图11)。可以看出,在对照大气CO2浓度和增加CO2浓度条件下,胞间CO2浓度均表现为开花期最大,乳熟期最小。与对照 CO2浓度(390 μmol·mol-1)相比,在 480 μmol·mol-1浓度下,胞间CO2浓度在拔节、孕穗、开花、灌浆和乳熟期分别增加了6.20%、10.56%、10.59%、15.60%和 8.95%,平均增加了 10.38%;在570 μmol·mol-1浓度下,拔节、孕穗、开花、灌浆和乳熟期分别增加了20.44%、24.96%、24.08%、31.71%和29.56%,平均增加了26.15%。
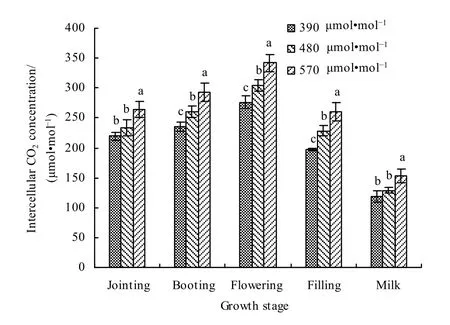
图11 大气CO2增加对不同生育期春小麦叶片胞间CO2浓度影响Fig. 11 Effects of different CO2 concentrations on the internal CO2 concentration of spring wheat leaves at different growth stages
2.4 CO2浓度对春小麦蒸腾速率的影响
研究结果表明,春小麦蒸腾速率的日变化趋势和净光合速率是一致的,也呈“双峰型”分布,属于“蒸腾午休型”(图12)。从08:00起随着光照强度的不断增强,蒸腾速率不断增大,并在 12:00达到峰值,中午由于气温升高,空气相对湿度很低,引起叶片失水过多,促使气孔关闭以减少蒸腾保持水分平衡,蒸腾速率由快速降低,并在14:00出现谷值,此后随着温度的降低和空气湿度的增加,水分胁迫程度减轻,蒸腾速率又开始回升,至 16:00出现第二个峰值,以后随着光照的减少而快速降低。从图12还可以看出,随着大气CO2浓度升高,春小麦叶片蒸腾速率呈减小的趋势。与对照相比,480 μmol·mol-1浓度和 570 μmol·mol-1浓度处理下,蒸腾速率分别平均降低了16.64%和25.36%。主要是高CO2浓度下,春小麦气孔导度降低,水分由叶片向外排放的阻力增大,导致蒸腾速率降低。
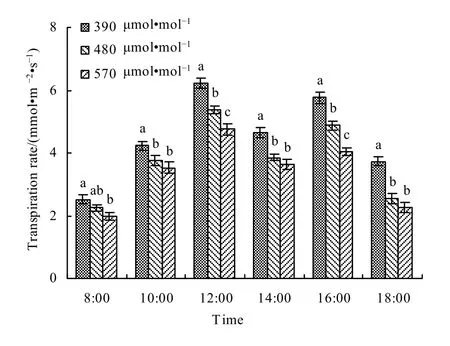
图12 不同大气CO2浓度下春小麦叶片蒸腾速率日变化Fig. 12 The diurnal variation of transpiration rate in spring wheat leaves under different CO2 levels
对不同生育期大气 CO2增加对春小麦蒸腾速率的影响进行分析(图13)。可以看出,在对照大气CO2浓度和增加CO2浓度条件下,春小麦蒸腾速率均表现为开花期最大,拔节期最小。与对照CO2浓度(390 μmol·mol-1)相比,在 480 μmol·mol-1浓度下,叶片蒸腾速率在拔节、孕穗、开花、灌浆和乳熟期分别减小了4.17%、9.92%、5.71%、5.82%和 7.52%,平均减小了 6.63%;在 570 μmol·mol-1浓度下,拔节、孕穗、开花、灌浆和乳熟期分别减小了9.64%、20.87%、6.15%、14.94%和10.46%,平均减小了12.41%。
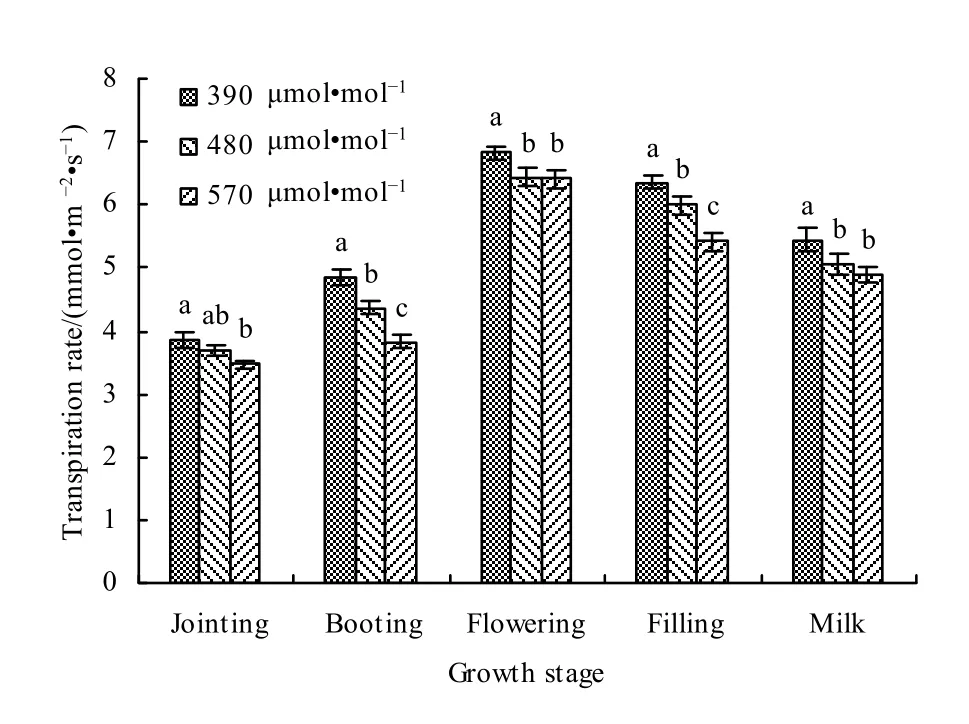
图13 大气CO2增加对不同生育期春小麦叶片蒸腾速率影响Fig. 13 Effects of different CO2 concentrations on the transpiration rate of spring wheat leaves at different growth stages
2.5 CO2浓度对春小麦水分利用效率的影响
不同 CO2浓度下春小麦水分利用效率的日变化表现为,在08:00 WUE达到最大,此后由于光合速率的增幅小于蒸腾速率的增幅,于是WUE逐渐下降,14:00以后又转而上升,至 16:00又达到次高峰,这是因为此时光合有效辐射虽小但蒸腾更小,此后WUE又迅速减小(图14)。从图14可以看出,随着大气CO2浓度的升高,水分利用效率逐渐增大。与对照相比,480 μmol·mol-1浓度和570 μmol·mol-1浓度处理下,水分利用效率分别平均增加了44.25%和82.60%。这主要是因为随着CO2浓度的升高,净光合速率提高,而气孔导度下降引起蒸腾速率的降低,从而促使水分利用效率提高。
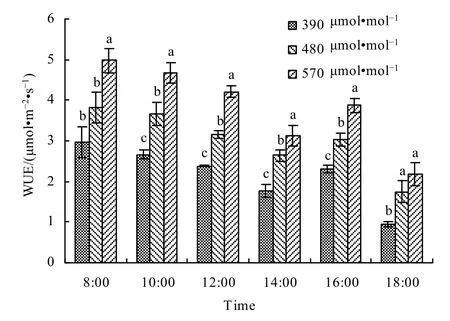
图14 不同大气CO2浓度下春小麦叶片水分利用效率日变化Fig. 14 Diurnal variation of water use efficiency in spring wheat leaves under different CO2 levels
对不同发育期大气 CO2增加对春小麦水分利用效率的影响进行分析(图15)。可以看出,在对照大气CO2浓度和增加CO2浓度条件下,水分利用效率表现为孕穗期最大,乳熟期最小。与对照CO2浓度(390 μmol·mol-1)相比,在 480 μmol·mol-1浓度下,叶片水平WUE在拔节、孕穗、开花、灌浆和乳熟期分别增加了 26.9%、28.3%、14.5%、19.7%和 25.0%,平均增加了 22.9%;在 570 μmol·mol-1浓度下,拔节、孕穗、开花、灌浆和乳熟期分别增加了 62.8%、62.1%、21.3%、40.3%和47.9%,平均增加了46.9%。
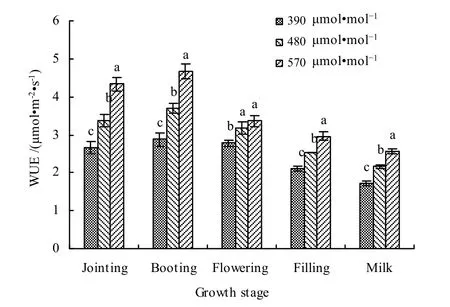
图15 大气CO2增加对不同生育期春小麦叶片水分利用效率影响Fig. 15 Effects of different CO2 concentrations on water use efficiency of spring wheat leaves at different growth stages
2.6 CO2浓度对春小麦叶片水势的影响
从图16可以看出,不同CO2浓度处理下的春小麦在拔节期的叶水势日变化表现为早晨和傍晚较大,中午后较小,呈现反抛物线曲线走向,在中午后出现水势曲线拐点。主要是由于清晨(07:00)太阳辐射弱,气温低,相对湿度大,叶片气孔开度小,蒸腾失水少,叶水势为白天最高值,随着气温的升高、太阳辐射的增强、空气相对湿度降低、叶片气孔开度逐渐加大,由蒸腾作用引起的叶片失水量也逐渐增加,叶水势不断降低(杨培林等,2012),于15:00达到全天最低值,此后,随着气温降低、太阳辐射减少、空气相对湿度增加及叶片气孔开度变小,由蒸腾作用引起的叶片失水也逐步减少,叶水势逐渐回升。从图16还可以看出,随着CO2浓度升高,叶片气孔开度降低,蒸腾失水减少,叶片水势不断增加,从而增强了春小麦对干旱胁迫的抵御能力。与对照相比,480 μmol·mol-1浓度和 570 μmol·mol-1浓度处理下,叶片水势分别平均增加了1.35%和3.64%。
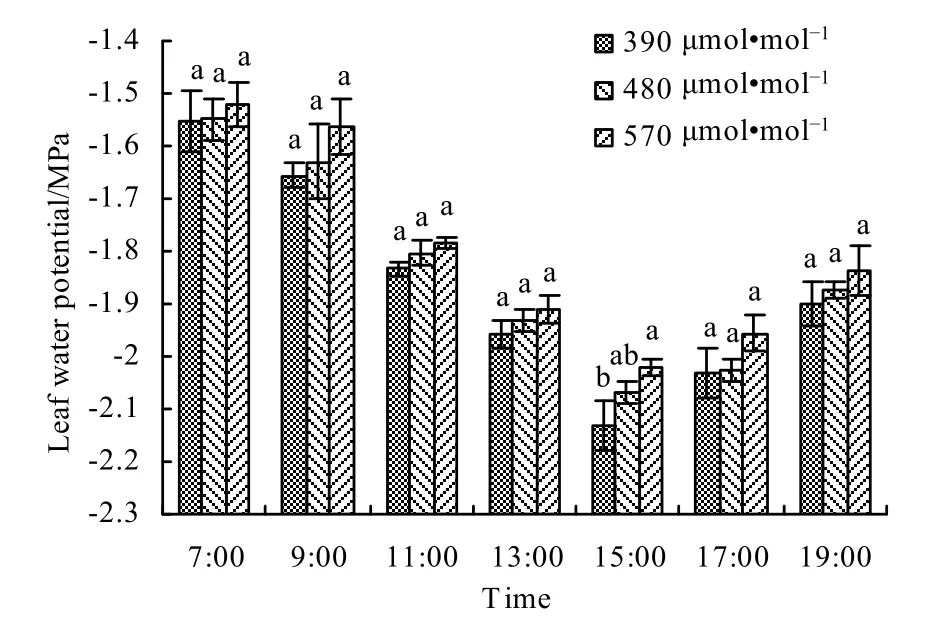
图16 拔节期不同CO2浓度下春小麦叶片水势的日变化Fig. 16 Diurnal variation of leaf water potential of spring wheat under different CO2 concentrations at jointing stage
3 讨论
光合作用是植物重要的生理过程之一,是植物生长发育和产量形成的基础,是外部生态因子和内部生理因子共同作用的复杂过程。
CO2是作物进行光合作用的必要原料,作物吸收太阳能,将CO2和水转化成糖。大气CO2浓度升高对植物最直接、最迅速的影响是植物光合作用(Cyrtis et al.,1998)。净光合速率是植物对高CO2浓度响应最直接最敏感的指标之一。根据前人研究,认为CO2对作物光合作用的影响存在短期和长期效应(李伏生等,2002),当作物长期处于高CO2条件下时,由于作物不能全部利用在高CO2下增加的碳水化合物(如淀粉),因此作物通过反馈抑制,使得CO2对作物光合速率的提高会随着时间的延长而逐渐减缓。本研究从试验周期来看,应属于长期,但是长期效应并不明显,并未出现光合适应现象。主要是因为C3作物出现光合适应现象对应的 CO2浓度约为 1000 μmol·mol-1左右(陈平平,2002;Leakey,2009),而本试验条件下的大气CO2浓度还没有达到这个标准,CO2浓度对春小麦的光合作用是不饱和的。本研究表明,大气 CO2浓度升高使春小麦的净光合速率显著提高。
气孔是水汽和CO2进出植物体内的门户,它同时控制着植物的光合和蒸腾作用(叶子飘等,2008),叶片上气孔对开口大小、数量及其分布状况的调节功能是植物适应环境、抵御外界胁迫的一项重要机制(Zheng et al.,2013)。以往研究表明,CO2浓度增加引起气孔阻力增大(康绍忠等,1995),导致气孔导度减小,进而抑制蒸腾,使作物的蒸腾速率减小,相应的整个生育期的蒸发蒸腾量减少。本试验中高CO2对春小麦的气孔阻力和蒸腾速率的影响也说明了这一点,这也与别人的研究结果一致(王静等,2009;蒋跃林等,2005)。
CO2浓度升高,作物不同水平WUE增加,这个结论大家基本上相对统一(康绍忠等,1995;Sionit et al.,1980)。但是在CO2浓度升高对作物WUE影响机理方面还存在不同的认识,尤其是叶片水平WUE增加的生理机制争议较多。根据以往研究结果,作物的水分利用效率是一个可遗传性状,同一作物的不同品种在需水、耗水及产量形成等方面存在较大差异。另外,CO2浓度、温度、水分条件等环境因子具有一定的协同效应,往往同时发生变化,进而对作物水分利用效率产生综合影响,因而可能导致WUE增加的生理机制方面存在差异(刘玉洁等,2013)。本试验研究认为,半干旱区春小麦叶片水平WUE的提高是由于CO2浓度增加了叶片净光合速率,而气孔导度下降又引起蒸腾速率的降低所致,这跟目前的大多数研究者观点一致。大气CO2升高导致作物生长对水分的高效利用,在因温室效应造成气候变化,从而使水分供应问题日益突出的环境条件下,对包括小麦生产在内的农业可持续发展有积极意义。
虽然春小麦光合及水分生理生态因子随着CO2浓度升高在变化趋势上跟前人研究结果基本一致,但是在增加和降低的幅度上有差异, 造成差异的原因可能是试验设计不同和研究区域的特殊性,也可能与供试小麦品种和试验环境有关。
本文仅探讨了 CO2浓度单因子对半干旱区春小麦光合及水分生理生态特性的直接影响,未综合考虑气候变化对春小麦带来的其他改变(如增温、降水)和CO2浓度增加导致的间接影响。此外,在研究过程中仅采用了1年的试验资料,只得到了一个初步结果,为了更深入认识CO2浓度增加对春小麦光合及水分生理生态特性的影响及其机制,还需要开展进一步的试验和研究。
4 结论
(1)在对照大气CO2浓度和增加CO2浓度条件下,春小麦叶片净光合速率日变化均呈“双峰型”分布,出现明显的“光合午休”现象。两种高CO2浓度条件下春小麦叶片净光合速率的提高,均表现为中午前后最为显著,在早晨和傍晚增强作用相对较小;在不同生育时期内,净光合速率均表现为开花期最大,乳熟期最小。与对照CO2浓度相比,在480 μmol·mol-1和 570 μmol·mol-1浓度(摩尔分数)下,叶片净光合速率在整个生育期平均增加了14.68%和28.20%。
(2)在不同生育时期内,气孔导度均表现为开花期最大,乳熟期最小。与对照CO2浓度相比,在480 μmol·mol-1浓度和 570 μmol·mol-1浓度下,叶片气孔导度在整个生育期平均降低了 15.29%%和24.83%。
(3)在对照大气CO2浓度和增加CO2浓度条件下,春小麦叶片胞间CO2浓度的日变化特点表现出斜“V”字型曲线。在不同生育时期内,胞间CO2浓度表现为开花期最大,乳熟期最小。与对照CO2浓度相比,在 480 μmol·mol-1浓度和 570 μmol·mol-1浓度下,叶片胞间CO2浓度在整个生育期平均提高了10.38%和26.15%。
(4)在对照大气CO2浓度和增加CO2浓度条件下,春小麦蒸腾速率的日变化呈“双峰型”分布,属于“蒸腾午休型”。在不同生育时期内,蒸腾速率均表现为开花期最大,拔节期最小。与对照CO2浓度相比,在 480 μmol·mol-1浓度和 570 μmol·mol-1浓度下,蒸腾速率在整个生育期平均减小了6.63%和12.41%。
(5)在不同生育时期内,在对照大气CO2浓度和增加CO2浓度条件下,叶片水平水分利用效率均表现为孕穗期最大,乳熟期最小。与对照CO2浓度相比,在 480 μmol·mol-1浓度和 570 μmol·mol-1浓度下,叶片水平 WUE在整个生育期平均增加了22.9%和46.9%。
(6)不同CO2浓度处理下的春小麦在拔节期的叶水势日变化呈现反抛物线曲线走向,在中午后出现水势曲线拐点。
