昆虫毒蕈碱型乙酰胆碱受体的研究进展
2021-03-30李建芳陆永跃齐易香
李建芳,成 杨,陆永跃,齐易香
(华南农业大学植物保护学院昆虫学系,广州 510642)
乙酰胆碱(acetylcholine,ACh)是昆虫重要的神经递质,它通过与不同类型的乙酰胆碱受体(acetylcholine receptors,AChRs)结合发挥作用。与脊椎动物类似,昆虫AChRs分为两类:烟碱型乙酰胆碱受体(nicotinic AChRs,nAChRs)和毒蕈碱型乙酰胆碱受体(muscarinic AChRs,mAChRs),其中nAChRs为离子通道受体,mAChRs为G蛋白偶联受体。昆虫nAChRs广泛分布于中枢神经系统,是以吡虫啉为代表的新烟碱类杀虫剂及生物农药多杀菌素的作用靶标(李健和巨修炼,2007;封云涛等,2009)。目前对昆虫体内mAChRs的研究相对较少,有研究报道mAChRs参与昆虫神经元轴突的延伸、表皮的形成(Clarketal.,2005)、代谢调节(Chowańskietal.,2018)、飞行运动(Buhletal.,2008)及学习记忆等(Ismailetal.,2006;Silvaetal.,2015)。mAChRs是一类潜在的杀虫剂靶标位点,研究该类受体,可为害虫防治提供新思路,具有很大的开发和利用价值。
1 毒蕈碱型乙酰胆碱受体的概述
1.1 mAChRs的结构
mAChRs是膜受体家族成员,通过与异三聚体鸟苷酸结合蛋白(G蛋白)偶联介导信号转导过程,因此属于G蛋白偶联受体家族(G protein coupled receptor,GPCR)。GPCR包括A类视紫红质受体、B类分泌素受体、C类代谢型谷氨酸受体、F类Frizzled/Smoothened受体等6个亚家族,其中mAChRs属于A类视紫红质受体家族(Hauseretal.,2006)。mAChRs的基本结构特征是具有7个跨膜螺旋结构域,其中7个跨膜区(TMⅠ-TMⅦ)由3个胞外环(O1-O3)和3个胞内环(I1-I3)连接而成,其氨基端位于细胞外侧,羧基端位于细胞内侧。mAChRs被激活后通过偶联G蛋白转导后续信号,引起细胞内多种生理生化反应。
1.2 mAChRs的克隆与分类
通过分子克隆的方法已经从脊椎动物中鉴定出5种mAChRs亚型(M1-M5),根据这5个亚型所转导的信号途径不同又分为两大类:M1、M3、M5属于Ⅰ类,M2、M4属于Ⅱ类(Caulfield and Birdsall,1998;Bubseretal.,2012)。然而,脊椎动物mAChRs的分类并不完全适用于昆虫(Eldefrawi and Eldefrawi,1997)。Onai等在1989年通过分子克隆方法从黑腹果蝇Drosophilamelanogaster体内获得了第一个昆虫mAChR基因序列(编码788个氨基酸,分子量大小为85 kDa),命名为DM1。该受体位于果蝇第二条染色体的右臂,其结构和功能与脊椎动物M1、M3、M5受体最为相似(Onaietal.,1989)。
随着分子生物学的深入研究和发展,mAChRs被报道在多种昆虫体内均有表达,包括摇蚊Chironomustentans(Wegeneretal.,1996)、家蚕Bombyxmori(Aizonoetal.,1997)、沙漠蝗Schistocercagregaria(Hintonetal.,1998)、烟草天蛾Manducasexta(Clarketal., 2005)、拟黑多刺蚁Polyrhachisvicina(Luetal.,2011)等,但未从分子水平验证昆虫存在多种mAChRs亚型。直到2013年Collin等人重新克隆黑腹果蝇mAChR(CG4356),发现Shapiro等(1989)发表的cDNA序列并不完整(DM1),并将该受体命名为A型mAChR;同时从黑腹果蝇中克隆了第二个mAChR(CG7918),命名为B型mAChR;B型受体药理学特征与A型受体及脊椎动物的mAChRs的药理学性质有很大的区别,对毒蕈碱敏感性要比乙酰胆碱低1 000倍,而且传统的拮抗剂对其不起作用(Collinetal.,2013)。从赤拟谷盗Triboliumcastaneum中也克隆得到了A型和B型mAChRs;通过生物信息学分析发现,A型和B型mAChRs的同源基因在其它节肢动物的基因组序列库中均能找到(Collinetal.,2013)。2016年从黑腹果蝇基因组中发现一种新的mAChR(CG12796),命名为C型mAChR,该受体的氨基酸序列与人类和昆虫中报道的其他mAChRs的序列相似性较低,其药理学特征与A型受体相近,与B型受体差异较大(Xiaetal.,2016)。因此,根据药理学特性和偶联的G蛋白不同,目前已知的昆虫mAChRs可以分为3大类:mAChR-A、mAChR-B和mAChR-C。为了进一步了解昆虫、线虫与脊椎动物的mAChRs在系统发育上的关系,笔者选取了GenBank中收录的9种昆虫的mAChR-A、8种昆虫的mAChR-B、5种昆虫的mAChR-C、线虫的3种mAChRs亚型以及人类的5种mAChRs亚型的氨基酸序列,利用Mega 7.0软件基于Neighbor-Joining法构建了分子进化树(图1)。
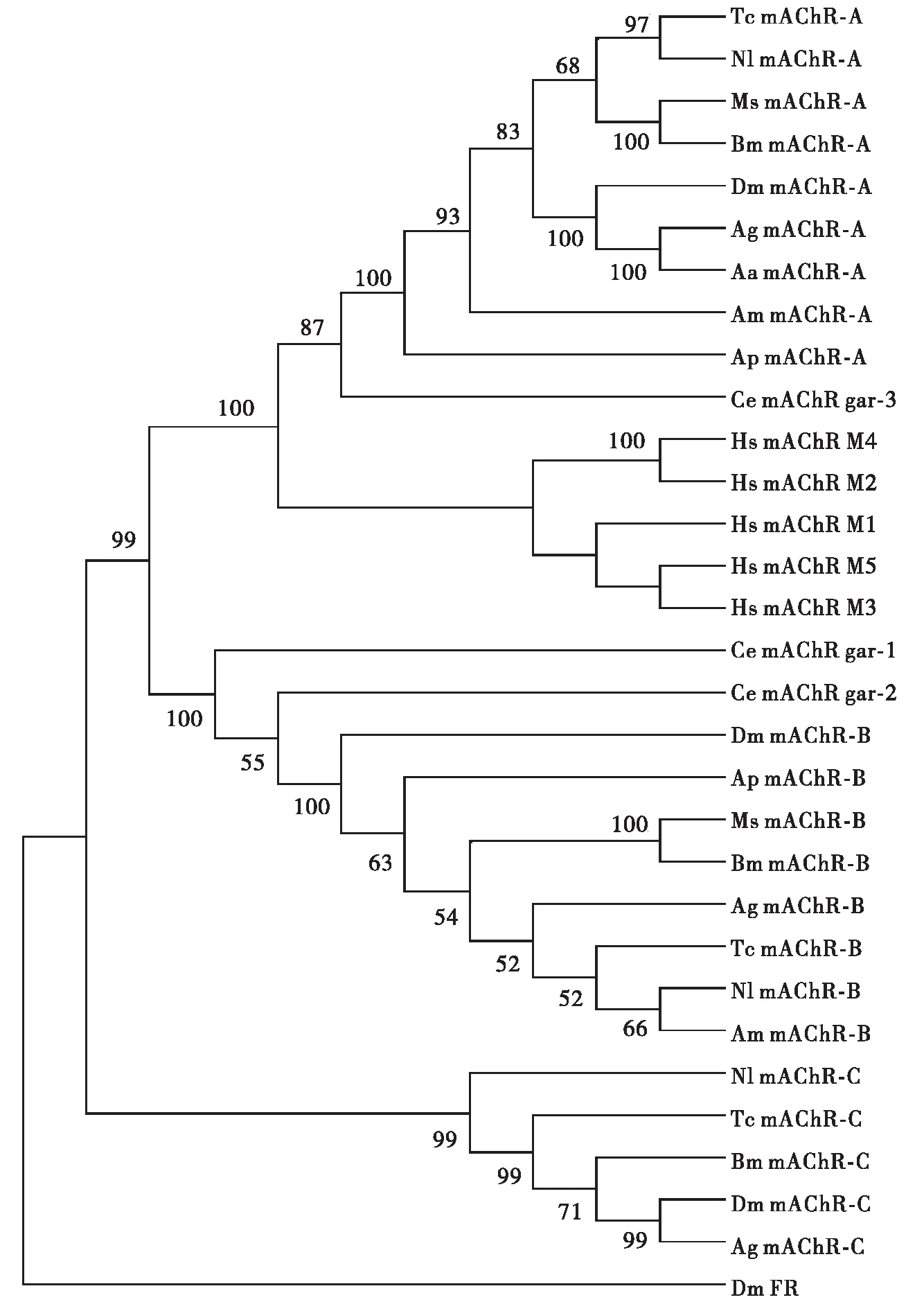
图1 不同物种mAChRs的系统发育分析Fig.1 Phylogenetic tree of mAChRs from different species注:mAChRs来源及GenBank登录号如下。Note: The origin of mAChRs and their GenBank accession numbers. Aa, 埃及伊蚊Aedes aegypti (Aa mAChR-A: XP_021708563.1); Ag, 冈比亚按蚊Anopheles gambiae (Ag mAChR-A: XP_314486.1; Ag mAChR-B: XP_003436890.1; Ag mAChR-C: XP_310742.3); Am, 西方蜜蜂Apis mellifera (Am mAChR-A: XP_395760.4; Am mAChR-B: XP_006558421.1); Ap, 豌豆蚜Acyrthosiphon pisum (Ap mAChR-A: XP_008187332.1; Ap mAChR-B: XP_008178482.1); Bm, 家蚕Bombyx mori (Bm mAChR-A: XP_012545273.1; Bm mAChR-B: XP_021208951.1; Bm mAChR-C: XP_004924179.1); Ce, 秀丽隐杆线虫Caenorhabditis elegans (Ce mAChR gar-1: NP_001024403.1; Ce mAChR gar-2: NP_001022594.1; Ce mAChR gar-3: NP_001024236.1); Dm, 黑腹果蝇Drosophila melanogaster [ Dm mAChR-A: NP_726440.1; Dm mAChR-B: NP_001287221.1; Dm mAChR-C: NP_572358.1; Dm FR (FMRFamide receptor): NP_001261347.1 ]; Hs, 人Homo sapiens (Hs mAChR M1: NP_000729.2; Hs mAChR M2: NP_000730.1; Hs mAChR M3: NP_000731.1; Hs mAChR M4: NP_000732.2; Hs mAChR M5: NP_036257.1); Ms, 烟草天蛾Manduca sexta (Ms mAChR-A: XP_030021104.1; Ms mAChR-B: XP_030032719.1); NI, 褐飞虱Nilaparvata lugens (Nl mAChR-A: XP_022187654.1; Nl mAChR-B: XP_022207514.1; Nl mAChR-C: XP_022186837.1); Tc, 赤拟谷盗Tribolium castaneum (Tc mAChR-A: NP_001280533; Tc mAChR-B: XP_015840901.1; Tc mAChR-C: EFA01319.1)
2 昆虫mAChRs的信号转导途径
GPCR信号转导过程的共同特点是受体被激活后会与G蛋白结合引起细胞内第二信使浓度的改变。G蛋白是由α、β、γ亚基组成,GPCR被配体激活后使得偶联的Gα亚基与β、γ亚基分离,引发Gβ、Gγ参与下游的信号调节,完成信号转导过程(李聪慧等,2019)。脊椎动物Ⅰ类mAChRs通过与Gαq/11结合,激活磷脂酶C(phospholipase C,PLC),PLC能够启动三磷酸肌醇级联反应,水解磷脂酰肌醇二磷酸(phosphatidylinositol 4,5-bisphosphate,PIP2)产生两种第二信使,即1,4,5-三磷酸肌醇(inositol triphosphate,IP3)和二酰基甘油(diacylacerol,DAG);IP3自由扩散并与内质网上的IP3离子通道受体结合,激活Ca2+通道,使Ca2+从内质网流向细胞内,从而使胞内Ca2+升高;DAG激活膜上的蛋白激酶(protein kinase C,PKC),PKC能够使很多蛋白发生磷酸化,从而产生生物学效应(Shapiroetal.,1989; Bubseretal.,2012)。Ⅱ类是通过与Gαi/o结合,降低腺苷酸环化酶(adenylyl cyclase,AC)的活性,抑制胞内ATP转化成cAMP,从而使胞内cAMP的含量下降(Felderetal.,2000; Wenzeletal.,2002)。与脊椎动物类似,昆虫mAChRs也可引发两种不同的信号转导途径(图2)。A型与C型mAChR与Gαq/11偶联,激活PLC,调节肌醇磷酸盐的水解,从而使胞内Ca2+升高;B型mAChR与Gαi/o偶联,抑制cAMP的形成(Renetal.,2015; Xiaetal.,2016)。
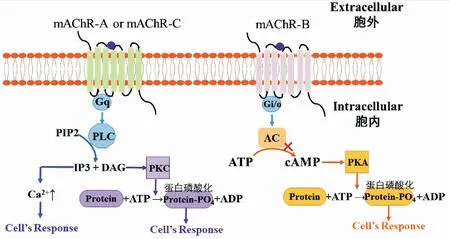
图2 昆虫mAChR的信号转导途径(修改自Felder et al.(1989))Fig.2 Signaling pathways of insect mAChRs(adapted from Felder et al.(1989))注:PLC,磷脂酶C;PIP2,4,5-二磷酸磷脂酰肌醇;IP3,三磷酸肌醇;DAG,二酰甘油;PKC,蛋白激酶C;ATP,三磷酸腺苷;ADP,二磷酸腺苷;AC,腺苷酸环化酶;cAMP,环腺苷酸;PKA,蛋白激酶A。Note: PLC, Phospholipase C; PIP2, Phosphatidylinositol 4,5-bisphosphate; IP3, Inositol triphosphate; DAG, Diacylglycerol; PKC, Protein kinase C; ATP, Adenosine triphosphate; ADP, Adenosine diphosphate; AC, Adenylyl cyclase; cAMP, Cycli adenosine monophosphate; PKA, Protein kinase A.
3 昆虫mAChRs的药理学特征
目前对昆虫mAChRs的研究主要集中在药理学性质上(表3)。脊椎动物5种类型mAChRs均可被传统的激动剂毒蕈碱充分激活,并被拮抗剂阿托品阻断(Caulfield and Birdsall,1998)。非选择性拮抗剂二苯羟乙酸-3-奎宁环酯(3-quinuclidinyl benzilate,QNB)、东莨菪碱和阿托品对果蝇DM1的亲和性最高,但对蝇类的毒性较轻(Hondaetal.,2007)。昆虫mAChRs 根据药理学性质分为三大类:mAChR-A与脊椎动物mAChRs(M1-M5)的药理学特性相似,可以被乙酰胆碱、毒蕈碱激活,也能被传统的拮抗剂阿托品、东莨菪碱及QNB抑制;mAChR-B虽然对乙酰胆碱很敏感,但对毒蕈碱的敏感性要低1 000倍,且对上述传统的拮抗剂均不起作用(Collinetal.,2013);mAChR-C的药理学特征与A亚型相近,与B亚型不同,可以被氧化震颤素M激活,传统的拮抗剂阿托品和东莨菪碱在100 μM时可以完全阻断100 μM乙酰胆碱引起的反应(Xiaetal.,2016)。通过与烟草天蛾幼虫高亲和QNB结合位点的结合分析,发现烟草天蛾mAChRs具有异质性,其中一些受体对吡嗪敏感,一些对吡嗪不敏感(Qazietal.,1996)。在摇蚊的药理学实验中发现与放射标记QNB的竞争顺序为QNB>阿托品>氧化震颤素,哌仑西平>美索曲明(Wegeneretal.,1996)。1,2,4-恶二唑和1,2,4-噻二唑作为mAChR激动剂,对褐飞虱Nilaparvatalugens等所测昆虫均表现良好的杀虫活性,衍生物3-甲基-1,2,4-恶二唑-2,2,1-四氢吡啶在蝇类神经组织中起着毒蕈碱激动剂的作用(Dicketal.,1997);1,2,4-噻二唑的衍生物3-甲基-5-(1,2,5,6-四氢吡啶-3-基)-1,2,4-噻二唑对褐飞虱的杀虫活性最高(Suzukietal.,2011)。目前没有针对mAChRs开发的农药,mAChR的激动剂和拮抗剂及其衍生物可作为杀虫剂的潜在资源。
4 昆虫mAChRs的功能
目前主要通过免疫组化的方法研究mAChRs在昆虫体内的分布,该类受体广泛存在于昆虫的神经系统中。Blake等发现黑腹果蝇mAChR主要位于大脑的触角叶,在肌肉组织中不表达(Blakeetal.,1993)。烟草天蛾mAChR在触角中有表达(Torkkelietal.,2004),并且触角中除了感觉神经元,胶质细胞和表皮细胞中也存在mAChR(Clarketal.,2005)。拟黑多刺蚁mAChR在卵期及整个幼虫期均有表达,并且各个品级蚂蚁的脑部都广泛表达mAChR,脑部又以蘑菇体、触角叶和视叶中表达量较高(Luetal.,2011)。关于昆虫mAChRs功能方面的研究,以下从运动行为、生长发育、感觉系统及学习记忆等方面进行总结。
4.1 运动行为
通过药理学的方法证明了用mAChR激动剂处理美洲大蠊Periplanetaamericana可以使触角长时间有节奏的运动,而拮抗剂可以抑制激动剂诱导的节律运动(Okadaetal.,2009)。同样在竹节虫Carausiusmorosus上用激动剂处理也发现mAChR对其持续、有节奏的触角运动有影响(Krauseetal.,2013)。章鱼胺等胺类物质和毒蕈碱激动剂可以激活沙漠蝗的飞行启动神经元,毒蕈碱的飞行激活不依赖于生物胺,而胺能的飞行启动需要mAChR(Buhletal.,2008)。毒蕈碱激活黑腹果蝇mAChR能降低幼虫的运动速度,而mAChR拮抗剂抑制这种作用。利用RNAi发现黑腹果蝇运动神经元中mAChR-A水平的降低并没有显著改变运动行为,但干扰运动神经元中mAChR-C的表达,以及干扰泛神经元中mAChR-C或者mAChR-A的表达均显著增强黑腹果蝇运动能力,说明mAChR-A和mAChR-C在调节昆虫运动行为方面发挥作用(Malloyetal.,2019)。
4.2 生长发育
雄性劳氏粘虫Mythimnaloreyi咽侧体中保幼激素的释放可被卡巴胆碱而非尼古丁显著激活,表明这种胆碱能效应可能是由mAChR引起的。此外通过药理学方法证明粘虫雄性成虫咽侧体的胆碱能调节是由类似于M2和M3受体的mAChR介导的(Kou,2002)。研究报道mAChR在家蚕的心侧体/咽侧体(corpus cardiacum/corpus allatum,CC/CA)复合体中有表达,该结构参与多种昆虫神经激素的分泌,如促前胸腺激素(prothoracicotropic hormone,PTTH)或者家蚕素(一种昆虫胰岛素样肽,insulin-like peptides,ILPs,在决定昆虫生长速率和大小、代谢特性、抗逆性和繁殖力等方面起着关键作用)(Shiraietal.,2001)。家蚕产生PTTH的神经分泌细胞中表达一种mAChR(Aizonoetal.,1997),且CC/CA复合体中ILPs的分泌受mAChR的调控(Shiraietal.,2001)。通过研究黄粉虫Tenebriomolitor营养组织中代谢物水平,发现mAChRs在糖原水平、血淋巴总糖浓度、ILPs水平等代谢调节中起重要作用,但不排除烟碱型受体的参与(Chowańskietal.,2018)。
另外,mAChR在沙漠蝗前肠的收缩中起重要作用(Hintonetal.,1998)。研究发现mAChR存在于黑腹果蝇幼虫的心脏组织中,毒蕈碱在低浓度和高浓度均可增加心率,表明mAChR在调节幼虫心率方面具有重要的作用(Malloyetal.,2016)。腹中神经节(mid-ventral ganglion,mVG)的谷氨酸能神经元在营养胁迫条件下通过mAChR调节昆虫化蛹,所以当食物中蛋白质缺乏时,mVG中mAChR或者IP3R缺失会影响果蝇幼虫化蛹(Jayakumaretal.,2016)。
4.3 感觉系统及学习记忆
西方蜜蜂Apismellifera和收获蚁Pogonomyrmexbarbatus能够区分同巢者和非同巢者与mAChR有关。药理学实验证明mAChR的拮抗剂处理蜜蜂和收获蚁都会干扰其对同巢者的识别能力,增加对同巢者的攻击,但是这种药物处理不影响嗅觉,也没有改变实验昆虫的运动能力,说明mAChR起气味记忆检索的作用(Ismailetal.,2008;Coates and Robertson,2013)。
用mAChR激动剂匹罗卡品处理有一周自然觅食经验的蜜蜂,可使其蘑菇体神经纤维网的体积增加(Ismailetal.,2006)。Dobrin等用匹罗卡品和东莨菪碱处理幼蜂和觅食蜜蜂,证明mAChR可以影响蜜蜂蘑菇体的生长,揭示了毒蕈碱在调节大脑可塑性方面的进化保守作用(Dobrinetal.,2011)。研究者发现黄斑黑蟋蟀Gryllusbimaculatus的mAChR可能与嗅觉联想学习中长期记忆的形成有关(Terazima and Yoshino,2010;Hasebe and Yoshino,2016)。



通过免疫组化发现,黑腹果蝇幼虫mAChR-A主要定位于中枢神经系统蘑菇体的蕈体萼和蕈体叶上,通过RNAi特异性干扰蘑菇体中mAChR-A的表达可以完全抑制幼虫新厌恶性嗅觉记忆的形成,并降低学习能力,表明mAChR-A是果蝇幼虫厌恶性嗅觉学习所必需的(Silvaetal.,2015)。Bielopolski等也证明了黑腹果蝇成虫的厌恶性嗅觉学习和短期记忆需要mAChR-A,同时发现mAChR-A在Kenyon细胞的树突中起作用,抑制气味反应(Bielopolskietal.,2019)。有研究报道,mAChR-A在黑腹果蝇的触角叶中广泛表达,作用于GABA抑制性局部神经元(inhibitory local neurons,iLNs)的一个亚群,通过药理学和遗传学手段证明mAChR-A可以增加iLNs的兴奋性,并增加iLN对气味的反应和减少嗅觉受体神经元(olfactory receptor neurons,ORNs)和iLNs之间突触所产生的短期抑制;还可以间接抑制兴奋性投射神经元(excitatory projection neurons,ePNs)对气味的作用(Rozenfeldetal.,2019)。
mAChR通过激活NO/cGMP/PKG信号级联调节Kenyon细胞(Terazima and Yoshino,2010),而NO不仅参与嗅觉记忆(Su and O’Dowd,2003),NO/cGMP/PKG在中央复合体内参与果蝇的视觉图形记忆(Wangetal.,2008),说明mAChR可能与视觉相关。通过药理学和钙成像技术发现,A型或者C型mAChR有助于增强若隐若现物体的辨别能力(Zhuetal.,2018)。黑腹果蝇mAChR-B在视觉回路的谷氨酸能中间神经元(glutamatergic interneuron,glu-lOLP)中表达,而glu-lOLP只对光线的减弱(OFF)有Ca2+立即升高的反应,这表明glu-lOLP是被OFF激活的;敲除mAChR-B可以消除glu-lOLP诱导OFF的Ca2+瞬变,表明mAchR-B/Gαo信号调节glu-lOLP中OFF响应(Qinetal.,2019)。
4.4 其它
在绿牧草蝗Omocestusviridulus和异翅雏蝗Chorthippusbiguttulus的脑部中央复合体小剂量注射mAChR激动剂会导致蝗虫鸣声更加尖锐且持续时间更长,并伴随发声的间隔时间缩短,表明蝗虫的鸣声与mAChR的激活有关(Wenzeletal.,2002)。研究还发现昆虫mAChR信号与其它神经递质系统之间的串扰,如毒蕈碱拮抗剂阿托品可以阻断黑腹果蝇中枢神经系统中由章鱼胺引起的持续性神经兴奋(Gross and Bloomquist,2018)。
5 小结与展望
新烟碱类杀虫剂如吡虫啉和噻虫嗪等作为nAChR激动剂,对多种昆虫都有杀虫活性,世界范围内已经有多种新烟碱类杀虫剂投入使用(李健和巨修炼,2007)。随之而来的害虫抗药性问题有待解决,因此开发新型杀虫剂非常必要。mAChRs是昆虫神经系统中重要的神经递质受体,属于GPCR家族。昆虫GPCRs在调控生殖、渗透、生长发育等方面具有重要作用(Jiangetal.,2017),也是最常被开发的药物靶点之一(Audsley and Down,2015;Hilletal.,2018)。因此,mAChR是潜在的杀虫剂作用靶标(Dicketal.,1997)。已有一些研究利用mAChR防治害虫,例如,驱蚊剂N,N-二乙基-3-甲基苯甲酰胺(N,N-Diethyl-3-methylbenzamide, DEET)对昆虫mAChRs具有选择性高亲和力,进而增强氨基甲酸酯残杀威的活性,因此mAChRs能够促进DEET和残杀威之间的协同作用,从而提高雌性埃及伊蚊Aedesaegypti的死亡率(Abd-Ellaetal.,2015)。另外,杀虫剂IR3535(10 nM)通过mAChRs的原位激活提高Ca2+浓度,增强新烟碱类杀虫剂噻虫啉的效果(Moreauetal.,2020)。
脊椎动物mAChRs的作用是众所周知的,它是多种药物的靶标位点(Felderetal.,2000;Kruseetal.,2014;Navarriaetal.,2015)。脊椎动物mAChRs的拮抗剂可减轻炎症反应,被广泛应用于人类疾病的治疗(Sales,2010)。M3受体调控小鼠对巴西日圆线虫Nippostrongylusbrasiliensis和肠道沙门氏菌Salmonellaenterica的适应性免疫反应(Darbyetal.,2015)。而在牡蛎Crassostreagigas和秀丽隐杆线虫Caenorhabditiselegans中也发现毒蕈碱信号在宿主免疫反应中起关键作用(Liuetal.,2016;Labedetal.,2018)。这为昆虫mAChRs的功能研究提供了一个新的方向。
目前有关昆虫体内mAChRs的研究还不够,研究的昆虫种类还不多。早期人们对昆虫mAChRs的研究都没有明确的分类,直到2016年有研究报道昆虫mAChRs分为A、B和C 3种类型,C型与A型的药理学性质相似;与脊椎动物相比,B型是无脊椎动物所特有的类型(Collinetal.,2013;Xiaetal.,2016)。昆虫(原口动物)和脊椎动物(后口动物)的mAChRs大约在7亿年前分裂(Renetal.,2015)。B型mAChR与脊椎动物mAChRs的药理学特性存在较大差异,脊椎动物和昆虫mAChRs之间的药理学性质差异可能是发现特异作用于昆虫mAChRs的新化学物质的关键(Gross and Bloomquist,2018)。因此加强对mAChRs药理学、信号转导和功能的研究,特别是无脊椎动物特有的B型mAChR的功能研究,可为新型杀虫剂的开发提供思路。
