微红梢斑螟(鳞翅目:螟蛾科)成虫触角感器的扫描电镜观察
2021-03-30王海燕胡振亮马梦然李继泉崔建州
靳 轩,王海燕,胡振亮,马梦然,李继泉,崔建州*
(1.河北农业大学林学院,河北保定 071000;2. 南水北调中线干线工程建设管理局河北分局,石家庄 050000 )
昆虫触角是接受外来化学信号的主要器官,其上分布着昆虫的大量嗅觉感器。昆虫通过这些感器来感知周围环境、寻偶、寻找取食寄主、选择产卵场所等(Gotzeketal., 2011)。嗅觉感器是一类特化的表皮细胞组织,不同的感器具有不同的功能(Altneretal., 1985)。研究昆虫触角感器的种类、形态、分布和功能,已经成为昆虫触角研究的重点,可以为探索昆虫的行为学奠定基础。
微红梢斑螟Dioryctriarubella属于鳞翅目Lepidoptera螟蛾科Pyralidae梢斑螟属Dioryctria,是一种主要危害松属植物的枝梢和球果的重要害虫,国内外均有分布(田恒德和严敖金, 1989)。其寄主非常广泛,在河北省北戴河联峰山主要危害油松,一年发生2代,主要以幼虫蛀害寄主主梢和幼树枝干,使枝梢折断枯死,影响树冠的整体形态,降低了树木的利用价值(李箐, 2003; 王荣国等, 2003),影响林木种子质量(常华, 2013)。由于微红梢斑螟幼虫钻蛀危害的特点,对于其防治技术主要采用化学防治为主,这不仅增加了害虫的抗药性,同时也带来了杀伤天敌及环境污染等问题。目前,根据行为学及化学生态学理论来讨论害虫的防治技术日益得到重视。利用害虫自身的生理生化微观特性来探索其宏观行为机制,可以为害虫防治提供新的理论依据。近些年,国内外的一些专家学者很重视在触角感器显微结构方面的研究(Schneider, 1964; 马瑞燕和杜家纬, 2000; 李竹和陈力, 2010)。现已对大蜡螟Galleriamellonella、松果梢斑螟Dioryctriapryeri、黄翅绢野螟Diaphaniacaesalis、二点螟Chiloinfuscatellus等多种螟蛾科的昆虫的触角感器进行了超微结构研究(杨立军和李新岗, 2007; 杨爽等, 2017; 孟倩倩等, 2017; 王锦达等, 2018),但国内外尚未见关于微红梢斑螟触角感器研究的报道。本试验利用扫描电镜对微红梢斑螟成虫触角的感器类型、形态和分布特点进行观察和研究,以期为微红梢斑螟的化学生态调控提供基础资料。
1 材料与方法
1.1 研究地概况
北戴河联峰山公园位于位于北戴河海滨中心西部,公园傍海,其占地约400 ha,东西横列约5 km。该地区受沿海季风影响,海洋性特征明显,湿度较大。年平均日照时间在2 700~2 850 h;年平均气温在8.8~11.3℃;盛夏日平均气温在22~25℃;年平均降水量为650~750 mm;年平均湿度在65%。园内地被杂草丛生,灌木穿插其中,主要以针叶树油松为主,兼有部分栎类、黄栌Cotinuscoggygria、碧桃Amygdaluspersicavar.duflex、刺槐Robiniapseudoacacia、毛白杨Populustomentosa和松科其它树种。
1.2 供试虫源
2019年6月,在河北省秦皇岛市北戴河区联峰山公园采集油松带虫枝果带回实验室于40 cm×40 cm×40 cm的养虫笼中进行饲养,饲养条件:温度为25±1℃,相对湿度为75%±5%,待其化蛹后,根据蛹期的形态特征将其雌、雄虫分开,选取初羽化且生长一致的雌、雄虫各10头。
1.3 样品的制样与观察
首先,在解剖镜下用镊子将微红梢斑螟的触角取下,将其浸泡在70%乙醇溶液中,利用超声波清洗仪进行清洗15 s,用2.5%戊二醛固定。24 h后用75%乙醇溶液冲洗,用80%、85%、90%、95%和100%乙醇溶液依次梯度脱水,各浓度脱水10 min,用乙酸异戊酯置换乙醇溶液,处理过夜,最后进行临界点干燥。
将干燥好的触角样品用双面胶粘在样品台上,喷金处理后用HITACHI SU8010扫描电镜进行观察、拍照。对已观察到的感器主要参照Schneider(1964)的标准进行鉴定和命名,并略加改进。
1.4 数据处理
感器的测量和数量统计的样本数量为6个触角或同种感器。所有数据均使用SPSS 22.0软件计算平均值和标准误。
2 结果与分析
2.1 微红梢斑螟触角的微形态
通过扫描电镜发现,微红梢斑螟的触角呈线状。雌、雄个体间的触角长度略有差异,雌虫触角长度为6 310±25.1 μm,雄虫触角长度为6 775±33.8 μm,雄虫触角略长于雌虫。微红梢斑螟触角背面均覆盖鳞片,鞭节的每个亚节均有两排鳞片,呈层层相叠的状态。触角感器大多分布在鞭节的腹面和侧面,柄节和梗节较少(图1-b)。
2.2 触角感器的类型和结构
微红梢斑螟触角上有10类感器,其中7类感器是雌、雄虫共有的,分别为毛形感器(Ⅰ、Ⅱ型)、刺形感器(Ⅰ、Ⅱ型)、耳形感器、腔锥形感器和Bǒhm氏鬃毛。栓锥形感器仅在雌虫触角上被发现,鳞形感器和钟形感器仅位于雄虫触角上;其中数量最多的是毛形感器。

图1 微红梢斑螟触角(a)和鞭节亚节形态(b)Fig.1 Morphology of Dioryctria rubella Hampson antenna (a)and subsegments of flagell (b)
2.2.1毛形感器(Sensilla trichoidea, ST)
基部无臼状窝,着生于触角表面的圆窝内,外形细长,前倾,向顶端渐细,触角各节均有分布,鞭节最多,按其形状不同分为Ⅰ型、Ⅱ型(图2-A, B)。Ⅰ型感器表面有螺旋形的环状纹,与触角表面成30°~70°夹角,Ⅱ型感器穿插分布于Ⅰ型之间,与Ⅰ型感器相比较短,基部较细,表面有竖向条纹且多孔,从基部往上开始弯曲,角度小于30°。雌虫的Ⅰ型毛形感器长为53.69±2.81 μm, 雄虫的Ⅰ型毛形感器长为57.27±3.71 μm,基部直径为3.24±0.04 μm;雌、雄虫的Ⅱ型感器长为30.2±0.20 μm,基部直径为2.03±0.3 μm。雄虫触角的毛形感器数量与雌虫相比较多,且Ⅰ型感器长度较长。
2.2.2刺形感器(Sensilla chaetica, SCH)
刺形感器基部几乎与触角表面垂直并且有突起的光滑臼窝,中部或中部偏上部位弯曲向前倾,顶部钝圆向上翘起,表面有螺旋状的环状纹。按其大小可分为Ⅰ型、Ⅱ型(图2-C,D,E),Ⅰ型分布于鞭节各亚节腹面,Ⅱ型分布于侧面,与Ⅰ型感器相比较细小。雌虫刺形感器Ⅰ型长为45.37±0.26 μm,基部直径为3.09±0.03 μm,雄虫刺形感器Ⅰ型长为49.00±1.07 μm,基部直径为2.69±0.03 μm;雌、雄虫Ⅱ型感器长为23.52±1.74 μm,基部直径为2.17±0.08 μm,雄虫触角的Ⅰ型刺形感器与雌虫相比较长,但是基部直径较小,Ⅱ型刺形感器的长及基部直径在雌、雄虫触角上无明显差异。
2.2.3耳形感器(Sensilla auricillica, SA)
耳形感器在雌、雄触角上均有较多分布,主要分布于鞭节各亚节上,其中在侧面的分布多于腹面(图2-G, I)。其外形类似于禾本科植物卷心叶状,有耳状凹槽,除此之外,表面还有纵向条纹且为多孔的结构,基部着生于触角圆窝内,顶端尖锐,耳形感器长为21.97±0.25 μm,基部直径为2.22±0.10 μm。
2.2.4腔锥形感器(Sensilla coeloconica, SCO)
生于忧患,死于安乐。席丰履盛,在一般人看来,那是多有面子呀!多享受啊!事实并非如此,中国自古就有物极必反、骄奢必纵,由俭入奢易、由奢入俭难等古训。“盖席丰履厚,其心易于放逸。”一旦身陷席丰履盛之地,滋生暴殄天物之心态,养成浮华奢华的习惯,最终不是走向危险之地,就是走向覆灭之地。
腔锥形感器在触角鞭节各亚节均有分布,是触角表面向下凹陷形成的浅圆腔(图2-F, H),直径10.96±0.32 μm,腔中央着生着一个直立的栓状突起,基部粗大,端部钝圆,由基部向端部聚拢,表面有纵沟,圆腔周围有13~15个的缘栓,端部尖锐,长短不一,向中央直立的栓状突起聚拢,缘栓表面也具有纵沟。
2.2.5Bǒhm氏鬃毛(Bǒhm bristles, BB)
Bǒhm氏鬃毛成群分布于触角柄节基部,外形短小尖细,呈椎形,表面光滑无孔,大多垂直于触角表面,不具基窝(图2-M),此类感器长为6.97±0.22 μm,基部直径为1.65±0.06 μm。
2.2.6鳞形感器(Sensilla squamiformia, SQ)
鳞形感器外形似鳞片,但比鳞片细长(图2-J),长为41.86±0.66 μm,基部直径为2.41±0.09 μm,呈梭型,表面有较深的纵向沟槽,基部有特化的臼状窝,分布于触角侧面紧挨背部鳞片处,分布不均匀,数量较少,观察时很难被发现,并且该感器只在雄虫触角上被发现。
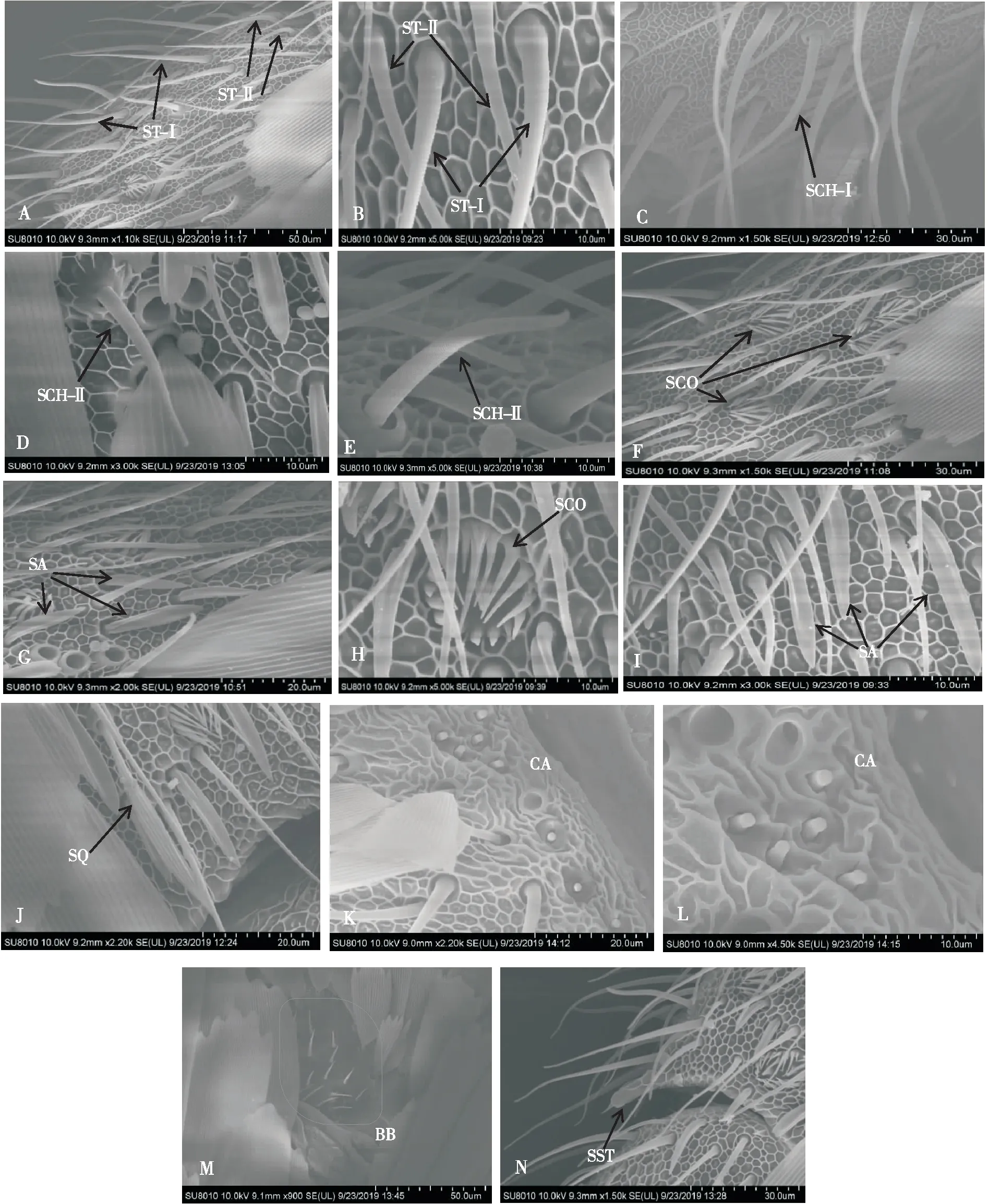
图2 微红梢斑螟触角感受器的扫描电镜照片Fig.2 Photograph of antennal sensilla of Dioryctria rubella by scanning electron microscopy注:A和B,毛形感器Ⅰ型(ST-Ⅰ)、毛形感器Ⅱ型(ST-Ⅱ);C,刺形感器Ⅰ型(SCH-Ⅰ);D和E,刺形感器Ⅱ型(SCH-Ⅱ);F和H,腔锥形感器;G和I,耳形感器;J,鳞形感器;K和L,钟形感器;M,Bǒhm氏鬃毛;N,栓锥形感器。Note: A and B, Sensilla trichoideaⅠ (ST-Ⅰ), Sensilla trichoideaⅡ (ST-Ⅱ); C, Sensilla chaeticaⅠ (SCH-Ⅰ); D and E, Sensilla chaeticaⅡ (SCH-Ⅱ); F and H, Sensilla coeloconica (SCO); G and I, Sensilla auricillica (SA); J, Sensilla squamiformia (SQ); K and L, Sensilla campaniformia (CA); M, Bǒhm bristles (BB); N, Sensilla styloconica (SST).
钟形感器着生于触角第一鞭节基部(图2-K, L),基部直径为1.74±0.05 μm,形状像一个倒扣的钟,稍突出于触角表面,表面光滑,周围有一圈光滑的稍向下凹陷的厚壁边缘,此感器只在雄虫触角上被发现。
2.2.8栓锥形感器(Sensilla styloconica, SST)
栓锥形感器(图2-N)均匀分布于触角腹面鞭节每亚节的端部,每亚节各一个,长为18.31±0.26 μm,基部直径为5.37±0.05 μm,从触角表面隆起延大约45°向外延伸,端部顶生一个小椎,基部至中上部有触角表面网状纹的延伸,中上部表面光滑。
2.3 雌、雄虫各类型感器的数量及差异
雌虫触角的8类感器中,毛形感器Ⅱ型的数量最多,可达1 009.4±9.11个,明显多于其他感器的数量;其次是毛形感器Ⅰ型,数量为949.3±13.02个,明显少于毛形感器Ⅱ型的数量;再次是耳形感器,数量为776.0±7.69个;刺形感器Ⅰ型的数量为56.3±8.75个,栓锥形感器的数量为43.0±0.52个,刺形感器Ⅰ型的数量多于栓锥形感器的数量,但两者的数量无明显差异;氏鬃毛的数量为18.8±0.31个,明显少于其他感器数量。雌虫各感器数量由多到少的排序为:毛形感器Ⅱ型、毛形感器Ⅰ型、耳形感器、腔锥形感器、刺形感器Ⅱ型、刺形感器Ⅰ型、栓锥形感器、Bǒhm氏鬃毛。
雄虫触角的9类感器中,数量最多的是毛形感器Ⅱ型,数量为1 820.0±8.34个,明显多于其他感器;其次是毛形感器Ⅰ型,数量为1 317.3±11.28个,明显少于毛形感器Ⅱ型的数量;鳞形感器的数量最少,数量为4.2±0.75个,与Bǒhm氏鬃毛和钟形感器的数量相比,三者数量无明显差异。雄虫各感器的数量由多到少的排序为:毛形感器Ⅱ型、毛形感器Ⅰ型、耳形感器、腔锥形感器、刺形感器Ⅱ型、刺形感器Ⅰ型、Bǒhm氏鬃毛、钟形感器、鳞形感器。
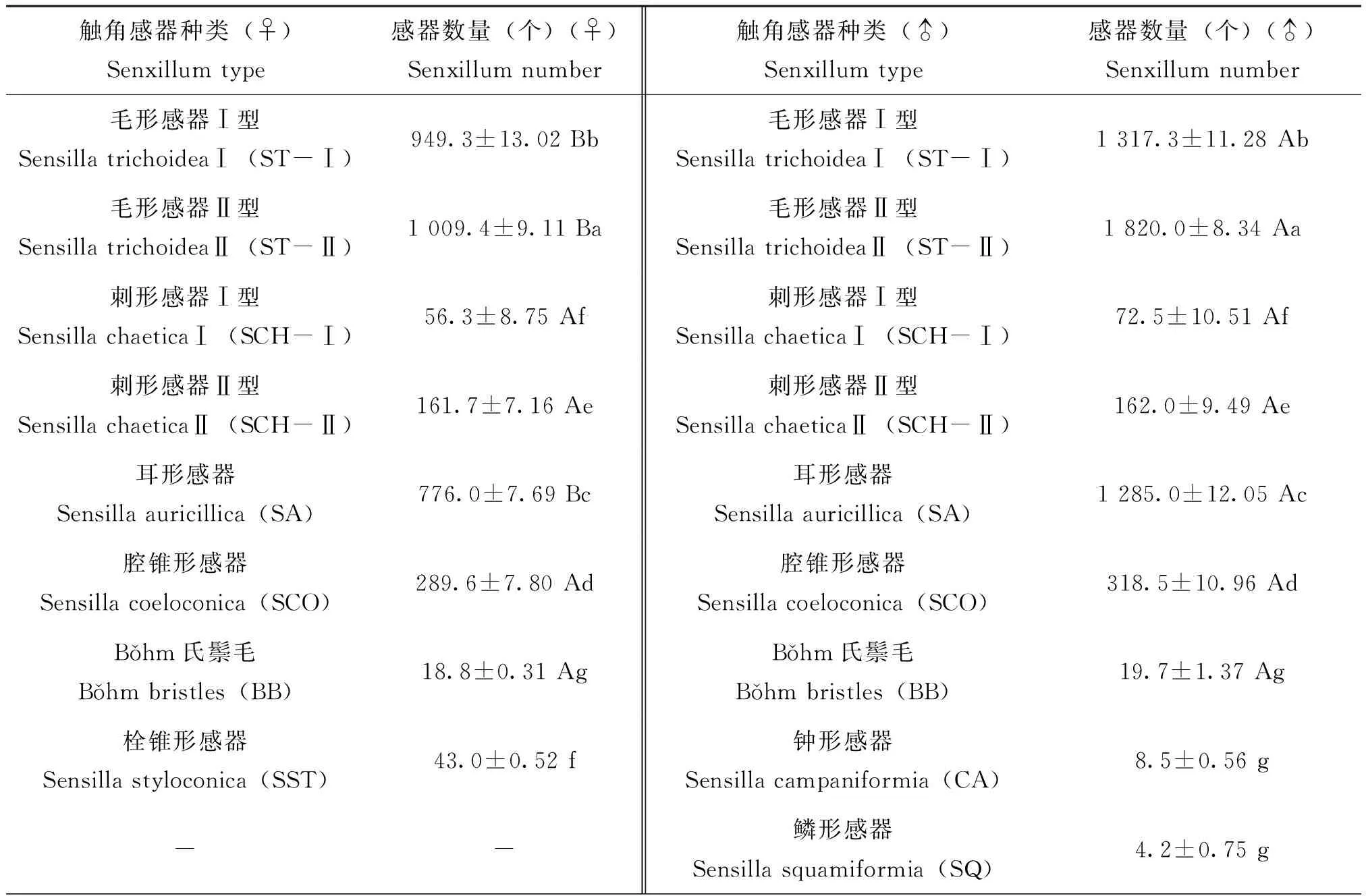
表1 微红梢斑螟雌、雄虫各类型感器的数量
将雌、雄虫触角上相同的感器进行T检验分析发现,相同感器的数量在雌雄虫触角上的分布是不相同的,其中雄虫触角的毛形感器Ⅰ型、Ⅱ型和耳形感器的数量明显多于雌虫;刺形感器和腔锥形感器及Bǒhm氏鬃毛在雌雄虫触角上的数量无明显差异。
2.4 常见的鳞翅目螟蛾科昆虫触角感器的类型及差异
不同的螟蛾科昆虫触角感器不同,微红梢斑螟与同科其他属的感器类型还是较为接近。毛形感器存在于大多鳞翅目昆虫中,并且按长短分为Ⅰ、Ⅱ型,微红梢斑螟也与之相似,但微红梢斑螟的Ⅰ、Ⅱ型的表面结构也存在差异,Ⅰ型表面具螺旋形的环状纹,与大多鳞翅目的昆虫相类似,但是Ⅱ型表面有竖向条纹且多孔,在螟蛾科的已发表的文章中未见报道,因此又有其自身独特性;刺形感器、腔锥形感器、耳形感器也是螟蛾科昆虫常见的几种感器类型,刺形感器有的表面光滑,如二点螟(王锦达, 2018),有的具有螺旋形的环状纹,如微红梢斑螟;腔锥形感器类型几乎相同,都具有菊花状的腔缘毛;耳形感器形与禾本科植物卷心叶类似,根据顶端尖锐或钝圆又可分为Ⅰ、Ⅱ型,有的只有Ⅰ型,如微红梢斑螟,有的只有Ⅱ型,如黄翅绢野螟(孟倩倩, 2017)、井上蛀果斑螟Assarainouei(邵淑霞, 2008);钟形感器分布较少,只在松果梢斑螟和微红梢斑螟触角上发现;鳞形感器外形都很相似,只在大蜡螟(杨爽, 2017)和二点螟上未见分布;栓锥形感器在大蜡螟、黄翅绢野螟和微红梢斑螟上有分布,其中大蜡螟和微红梢斑螟的栓锥形感器最为相似,但在微红梢斑螟中其只在雌虫触角上有分布,而在另两者中则是雌雄虫都有分布;存在性二型的昆虫有松果梢斑螟、微红梢斑螟、井上蛀果螟;通过以上比较发现,微红梢斑螟与同属的松果梢斑螟(杨立军, 2007)的感器类型、形态分布最为接近,不同的是刺形感器Ⅱ型与栓锥形感器未在松果梢斑螟上被发现。

表2 常见的鳞翅目螟蛾科触角感器
3 结论与讨论
本研究通过对微红梢斑螟雌、雄成虫触角进行扫描电镜观察,发现触角呈线状,雄虫触角略长于雌虫。在雌雄触角上共发现10类感器,其中毛形感器Ⅰ、Ⅱ型,刺形感器Ⅰ、Ⅱ型、耳形感器、腔锥形感器和氏鬃毛这7类感器在雌雄虫触角上均有分布,而鳞形感器和钟形感器只在雄虫触角上被发现,及栓锥形感器仅在雌虫触角上被发现,并且雄虫毛形感器、耳形感器数量多于雌虫,存在性二型现象。这一研究结果与松果梢斑螟最为相似,不同的是刺形感器Ⅱ型与栓锥形感器未在松果梢斑螟上被发现。
昆虫触角有多种感器,且感器的类型及功能多样。每种感器的类型、功能、数量及分布等不仅与其种类、性别有关,还与其生活环境及与寄主植物的相互作用相关。因此明确昆虫触角感器的种类、分布及数量,对研究各感器的功能与寄主植物互作方面具有重大意义。对于鳞翅目昆虫触角显微结构的研究已较多。在鳞翅目昆虫中,毛形感器是普遍存在的一类感器,一般具有性二型现象,且该感器现已被证实不仅能感受性信息素,而且还具有触觉、嗅觉、味觉、机械感受等功能(Castrejón Gómezetal., 2003;Dolzeretal., 2003),雄性昆虫对于性信息素的敏感程度高于雌虫(Zhaoetal., 2018),这或许也是微红梢斑螟雄虫触角上毛形感器明显多于雌虫的原因。刺形感器基部几乎与触角表面垂直,整个感器延伸的高度比其他感器高许多,并且基部有特化的白臼窝,基于刺形感器的这些形态特征,被认为具有感受机械刺激的功能(Schneider, 1964)。
Ebbinghaus等(1998)报道在卷蛾科的苹果蠹蛾Cydiapomonella上发现表面多孔的耳形感器具有感受微量信息素的功能,在张建(2017)的报道中,也认为花布灯蛾Camptolomainteriorata的此种感器可能也有类似功能。国外学者Ammagarahalli (2015)使用单感器记录仪发现表面多孔的耳形感器对植物气味和信息素化合物起反应,微红梢斑螟中耳形感器表面具有纵向条纹且多孔,或许也具有以上功能。Alter(1983)做电生理反应试验发现,有腔缘毛的腔锥形感器不仅具有嗅觉功能,而且还能感知温度和湿度的变化;也有研究发现,这类感器上具有感觉神经元,既能对短链脂肪酸及单萜醇类物质有一定的感觉反应,也对雌虫寄主产卵地选择上起作用(Altneretal., 1983; Cossetal., 1998; Hunger and Steinbrecht, 1998),微红梢斑螟的腔锥形感器具有菊花状的腔缘毛,因此,推测其具有以上功能。Bǒhm氏鬃毛在鳞翅目昆虫中也较为常见,Schneider(1964)认为其是一种机械感受器,另有国外学者Krishnan等(2012)通过行为学试验发现,此感器在蛾子飞行中能够调节其触角的定位。
鳞形感器数量较少、紧邻鳞片且在鞭节不均匀分布,很难观察到,目前关于此感器的功能还未见报道,因其无孔的表面,Krishnan 等(2012)推测此感器可能具有机械感受功能。钟形感器被认为能够感受角质层的机械形变,是一种本体感受器(Mclver, 1975),也有研究表明其对气味、二氧化碳、温湿度敏感(Dietz and Humphreys, 1971)。
栓锥形感器仅在雌虫触角上被发现。此类型感器在鞘翅目枣食芽象甲Scythropusyasumatsui中也仅分布在雌虫触角上;在功能上,张峰等(2019)认为枣食芽象甲雌虫对枣树的挥发性气味更为敏感,可能与栓锥形感器仅在枣食芽象甲雌虫上分布有关。在关于寄主油松挥发物对微红梢斑螟成虫生测试验研究中,我们发现雌虫对寄主油松挥发性气味更为敏感,因此可推测栓锥形感器具有味觉、嗅觉的功能,对雌虫的取食及寄主选择方面至关重要。
微红梢斑螟属于蛀干害虫,其幼虫钻蛀枝干危害寄主植物,但在幼虫期防治害虫难度较大。因此,成虫期的防治尤为重要,其成虫触角感器在感知周围环境、寻偶、寻找取食寄主、选择产卵场所等有重要作用。本研究通过对微红梢斑螟触角扫描电镜的观察,发现其触角感器类型丰富,但由于多数的感器功能只是靠前人的推测而来,并未得以证实,今后还需要利用单细胞记录仪结合透射电镜、触角电位等生理技术对各感器功能做进一步研究,为明确微红梢斑螟触角感器与其对寄主植物的选择机理之间的关系奠定基础。
