昆虫假死行为研究进展
2021-03-30李会娟杨开朗温俊宝
李会娟,杨开朗,王 倩,温俊宝
(北京林业大学林学院,北京 100083)
动物受到外界刺激如天敌刺激时,会自发进入强直静止状态,身体蜷缩静止不动呈“死亡”状,或从原停留处落下(Miyatake, 2001a; Ruxton, 2006),这一现象被称为假死(Death-feigning、Thanatosis、Tonic immobility、Animal hypnosis、Playing dead、Immobilization catatonia、Play possum)(Humphreys and Ruxton, 2018),属于次级防御行为(Matthews and Matthews, 2010; Ruxtonetal., 2018),是先天性的,是长期进化中保留下的适应特征(Edmunds, 1975)。
假死行为广泛存在于动物界各个门类,包括节肢动物门,脊索动物门中哺乳纲、两栖纲、鱼纲、爬行纲等,软体动物门等,有文献记载的共71个物种(见表1)。节肢动物门中除蛛形纲和软甲纲有假死行为外,昆虫纲中最为普遍(Alonsozarazaga and Lyal, 1999),有40种昆虫被明确报道具有假死行为,其中鞘翅目有22种,而象甲科最多,包括甘薯象甲Cylasformicarius(Acheampong and Mitchell, 2010)、松树象甲Hylobiusabietis(Sibuletal., 2013)、臭椿沟眶象Eucryptorrhynchusbrandti(Lietal., 2019)、沟眶象Eucryptorrhynchusscrobiculatus(张艳,2015)、女贞粗腿象Ochyromeraligustri(侯冬梅和涂小云,2014)、车轴草籽象Tychiuspicirostris(Duporte and Melville, 1916)等。
昆虫的假死行为对提高逃逸率和存活率等有着十分重要的作用,近年来,逐渐受到国内外研究者的重视。本文拟综述昆虫假死行为的研究现状,重点探讨假死行为的发生机制,分析现存问题,展望今后研究方向。
1 昆虫假死行为研究概况
1.1 相关文献资源和机构
从1906年开始,到2019年10月,以“假死”(Death-feigning、Thanatosis、Tonic immobility)为关键词,通过谷歌学术(英文)、中国知网(中文)数据库共检索出1 350篇相关文献。深入筛选发现其中仅279篇涉及昆虫,2000年后发表的有229篇。进一步统计2000-2019年国内外涉及昆虫假死行为的论文(图1),20年间有46篇是明确研究昆虫假死行为的,其中国内6篇。涉及昆虫假死行为的文献呈逐年递增趋势,但深入研究昆虫假死行为的论文增加趋势不明显(图1-a)。37篇文章探究昆虫假死行为特征,探索假死行为发生机制的仅9篇,每年发表数量见图1-b。
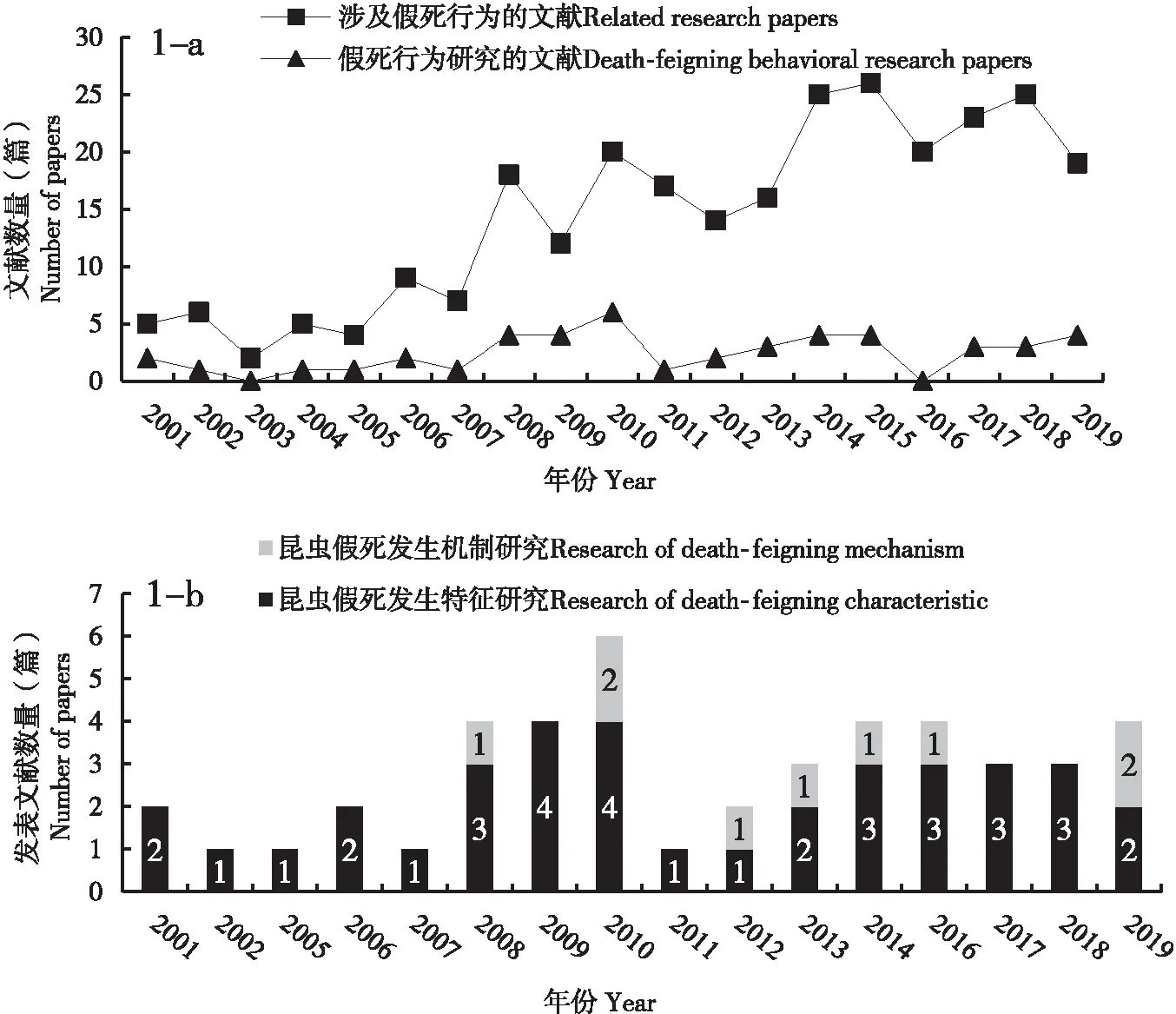
图1 假死行为的学术论文发表情况(1-a,2001-2019年与假死行为研究的学术论文发生趋势;1-b,昆虫中假死行为发生时间和发生机制研究内容的文献数量。)Fig.1 Number of papers in death-feigning(1-a, 2001-2019, the trend of academic papers on the study of death-feining behavior; 1-b, the number of papers with different content of death-feigning in insect.)

表1 具有假死行为的物种统计Table 1 Data on different species that performed death-feigning behaviour

续表1 Continued table 1

续表1 Continued table 1

续表1 Continued table 1
研究昆虫假死行为的机构主要在国外,如日本冈本大学进化生态学实验室、英国圣安德鲁斯大学生物学院、爱沙尼亚农业大学植物保护学院和塔尔图大学生态与地球科学研究所等。2019年北京林业大学林木有害生物防治北京市重点实验室首次发文确认臭椿沟眶象的假死行为。
1.2 国外主要研究工作回顾
昆虫的假死行为研究最早可追溯至1906年,Holmes首次研究了几种蝎蝽的假死行为并测试假死发生时间,发现温度的高低影响假死发生时间(Holmes, 1906)。
爱沙尼亚的Metspalu和Krams等人从生理方面对马铃薯叶甲Leptinotarsadecemlineata的假死行为进行了探讨,发现假死前后代谢率变化显著,正常状态下的代谢率是假死状态下的1.5倍(Metspaluetal., 2002),但未考虑虫体大小的影响,数据结果或存在争议。
英国圣安德鲁斯大学生物学院的Ruxton博士则从进化生态学角度认为,假死行为是一种重要的天敌防御模式,有效躲避天敌(Ruxton, 2006; Ruxtonetal., 2018)。
日本冈本大学的进化生态实验室Miyatake等人,2001年至今共发表19余篇昆虫假死行为的论文,主要研究对象为赤拟谷盗Triboliumcastaneum、四纹豆象Callosobruchusmaculatus、绿豆象Callosobruchuschinensis等仓储害虫,从验证假死行为发生特征入手,包括性别、虫体大小、饥饿、飞行力和活动力、温度、光照等方面,探究昆虫假死行为生理、基因和分子基础。他们证实假死时间在雌雄虫之间存在差异,温度影响假死发生时间,昆虫的活动力和飞行力与假死存在基因负相关性(Ohno and Miyatake, 2007; Miyatakeetal., 2008b)。近年,Miyatake等人认为假死或与脑部生物胺密切相关(Miyatakeetal., 2008b; Matsumuraetal., 2016),但未明确假死前后生物胺表达量是否存在差异,又从基因层面对比分析赤拟谷盗活动力与假死行为发生的联系(Uchiyamaetal., 2019),开始探究假死行为的发生机制。另外,Miyatake(2019)首次从觉醒方面探究昆虫假死行为特征,发现假死过程中的赤拟谷盗受到震动会觉醒,假死时间与震动强度正相关,认为假死觉醒或有遗传基础。
国外假死行为研究已经逐渐多样化,从生理生化、基因、生态学等角度探究假死行为特征和发生机制。
1.3 国内主要研究工作回顾
2010年开始有中国学者探究昆虫的假死行为,涉及种类如长壮蝎蝽Laccotrephesrobustus(孟凡明和梁醒财,2010)、毛健夜蛾Brithyscrini(朱敏等,2016)、丝棉木金星尺蛾Calospilossuspecta(曹亚汝等,2018)、女贞粗腿象甲(侯冬梅和涂小云,2014)、臭椿沟眶象(Lietal., 2019)等,主要研究其假死行为特征。
孟凡明(2009)将假死行为分为3种类型,伪装死亡型假死、逃避搜捕型假死、抑制进食型假死,并认为应将假死归为拟态行为;侯冬梅等人认为假死的影响因素与背景色相关;李会娟等(2019)明确了饥饿对臭椿沟眶象假死的影响,初步完成近缘种沟眶象和臭椿沟眶象假死行为特征的对比分析。
总体来说,我国的研究工作主要探讨了昆虫假死的发生特征,起步时间晚,研究基础较薄弱。
2 昆虫假死行为特征研究
2.1 发生特征
昆虫的假死行为(Death-feigning)可以定义为:当昆虫接收到外界刺激信号后,身体各系统做出反应,呈现特定的姿态,如足或触角收缩、躯体弯曲,自发地进入强直静止的状态。昆虫假死时大脑正常支配功能不受干扰,是一种低耗能、高警惕性的防御模式,在形态和生理上与真正的死亡有着本质区别。
假死发生时,多数昆虫保持特有的假死姿态。在形态上,虫体保持僵直不动,触角和附肢强烈收缩并紧贴于身体腹面,肌肉紧张(Humphreys and Ruxton, 2018)。有的昆虫幼虫,如细胸金针虫Agriotessubrittatus,假死时常弯曲身体(Ritteretal., 2016)。除此,象甲类昆虫,如臭椿沟眶象假死时喙完全收缩于胸部腹面(Lietal., 2019)。若不继续施加刺激,几分钟或十几分钟后,昆虫假死觉醒后恢复正常体态并迅速逃跑。不同昆虫之间的假死行为特征既有相似性,又有其特有的固定模式,受环境和自身因素的影响。依据动物假死发生后的形态特征,划分为蜷缩型、弯曲型、强直型、静止型等4种类型,具体见表2。目前,昆虫的假死发生形态多为蜷缩型和弯曲型,而强直型假死多发生在爬行类、鱼类,静止型假死多发生在鼠、穴兔类等高等哺乳动物,形态上的不同是昆虫与其他动物假死行为特征的主要差异。

表2 不同物种的假死发生特征
此外,昆虫假死后的内部生理活动也有显著变化。Sibul等人运用红外心电图和呼吸计记录松树象甲假死期间呼吸模式,证明假死期间松树象甲依然保持标准的气体交换循环(closed, flutter, ventilation periods, CFV cycle)(Sibuletal., 2013),但是假死结束时出现CO2爆发点,或许假死期间昆虫正常呼吸,但假死前后的代谢有差异;而从大脑活跃度来看,假死状态下的动物依然保持警觉性。Davie等人认为,动物进入假死状态后,假死大脑和清醒时一样活跃,对外界时刻保持警惕(Davieetal., 1993)。或许,假死行为仅单纯迷惑天敌,利于逃避防御。比如,沟眶象和臭椿沟眶象假死结束后,迅速从侧翻假死状态恢复正常体态逃逸(张艳,2015)。细胸金针虫假死觉醒后,会立即挖掘掩埋自身虫体,躲避天敌(Ritteretal., 2016)。
2.2 发生强度
关于假死行为发生的程度,前人多用假死强度(Death-feigning intensity)一词描述(Ohno and Miyatake, 2007),主要由假死发生时间和假死频率定量评估。已知的昆虫假死发生强度如表3所示,不同物种之间的假死发生时间和频率差异较大。Matsumura等人认为不同物种甚至是同一物种雌雄间出现的假死强度变异现象或可用假死深度(Depth of death feiging)解释(Matsumuraetal., 2016),但未具体证实这一定义。
2.3 诱发因素
假死行为可由多种方式诱发,如触觉、视觉、听觉或混合刺激。不同物种具有不同的效应刺激方式,如光束可以引发马铃薯叶甲的假死行为,声波刺激双斑蟋发生假死反应(Nishino and Sakai, 1996; Metspaluetal., 2002);意大利蜜蜂受到一定频率的声波刺激后会假死(Acheampong and Mitchell, 2010);高温可诱发负子蝽假死行为发生(Holmes, 1906)。但是,多数昆虫均可用镊子触碰腹部产生机械刺激达到假死状态,包括臭椿沟眶象、沟眶象、甘薯象甲、绿豆象、四纹豆象和赤拟谷盗等(Lietal., 2019; Miyatakeetal., 2019)。假死行为最直接有效的诱发因素或是机械刺激。
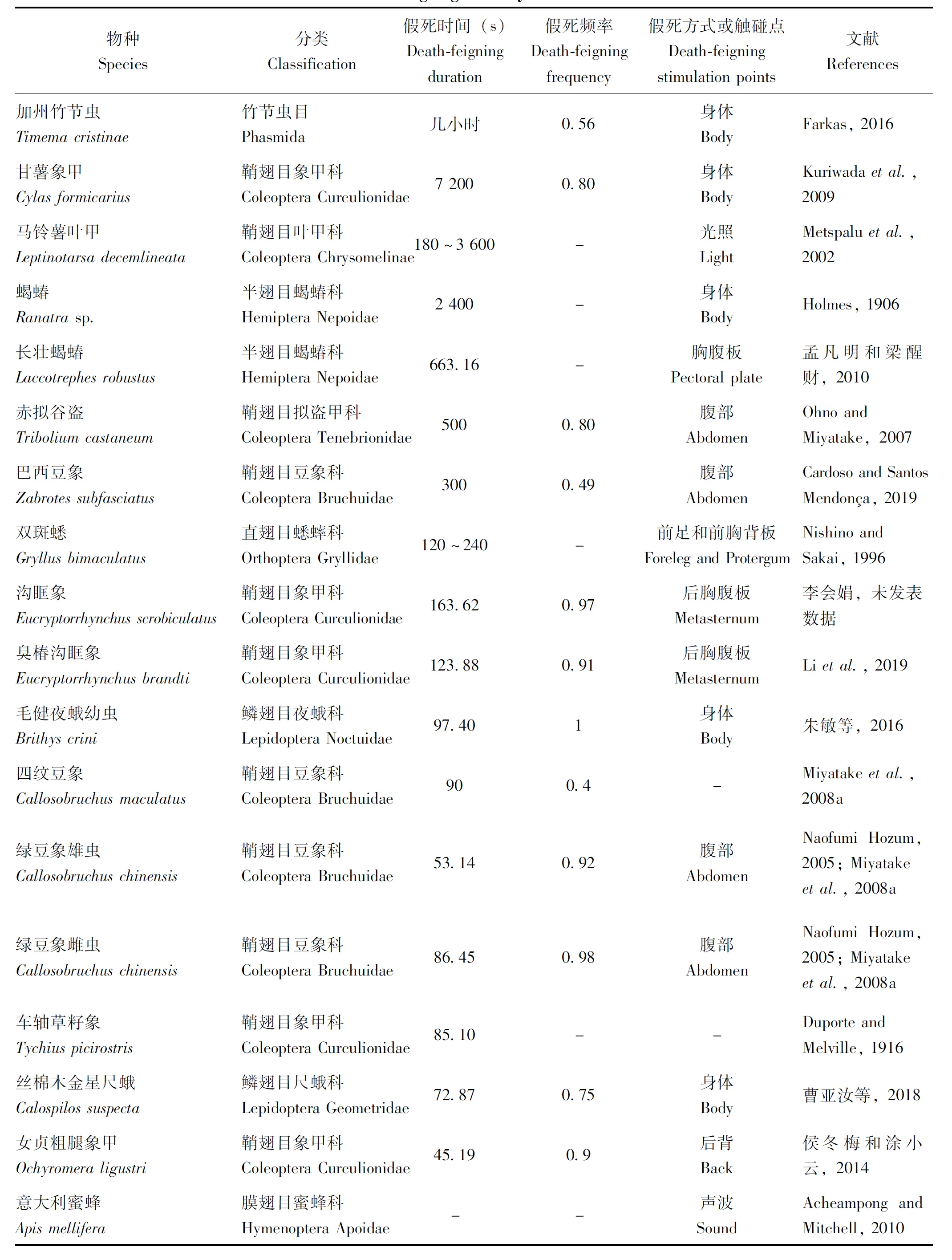
表3 不同昆虫的假死发生强度Table 3 Death-feigning intensity of different insects
2.4 影响因素
不同昆虫之间的假死行为特征既有相似性,又有其特有的固定模式,受环境和自身因素的影响(Humphreys and Ruxton, 2018)。环境中的背景颜色可影响丽蝇蛹集金小蜂雌蜂的假死,毛健夜蛾幼虫和女贞粗腿象甲的假死发生率在红、黄、绿背景下有显著差异(侯冬梅和涂小云,2014;朱敏等,2016);四纹豆象和绿豆象在高温环境下减弱假死发生频率,假死发生时间与温度负相关(Hozumi and Miyatake, 2005);而低频的声音刺激可以减弱沟眶象和臭椿沟眶象的假死发生时间(李会娟,未发表数据)。
昆虫自身内部影响因素包括性别、体重或虫体大小、龄期、饥饿程度等(Yasuda, 1998; Ritteretal., 2016)。绿豆象中,虫体大小与假死时间显著相关(Hozumi and Miyatake, 2005),且雌性假死时间显著低于雄性。然而,四纹豆象的假死时间雌性高于雄性(Miyatakeetal., 2008a)。加州竹节虫Timemacristinae假死行为也受虫体大小影响,且日龄低的竹节虫较日龄高的竹节虫易假死(Farkas, 2016)。饥饿是一种重要的内部生理影响因素,Miyatake等人在甘薯小象甲饥饿试验中发现,饥饿处理的甘薯象甲的假死比例显著低于未饥饿处理组,且雌虫的假死时间长于雄虫(Miyatake, 2001b),此外饥饿的臭椿沟眶象假死发生频率随饥饿时间的增加而降低(Lietal., 2019)。
综上所述,昆虫假死行为的诱发和影响因素,主要有机械刺激、光照、声音、温度、背景色以及虫体自身条件等。明确参与调节昆虫假死行为的因素,利于假死行为的定性分析,为进一步的发生机制研究奠定基础。
3 昆虫假死行为发生机制研究
当前昆虫假死行为研究多集中于假死行为的发生特征、诱发因素,与发生机制相关的生理生化、神经生物学、基因层面等的研究不足,尚无法详细解释昆虫假死行为发生机制。
不同物种具有不同假死诱发因素,且假死行为不同物种甚至个体间差异显著,导致宏观假死行为研究较难控制变量,试验繁琐复杂,工作量大。近年来,生理学、神经生物学和分子生物学层面的技术手段逐渐成熟,为揭秘昆虫假死行为的发生机制提供了条件。
3.1 激素调控
在无脊椎动物中,生物胺是一类非肽类神经激素,在调控感觉器官、影响中央神经系统的状态与起始行为的改变上扮演了相当重要的角色(Pirrietal., 2009),最常见的5种生物胺是章鱼胺(Octopamine, OA),酪胺(Tyramine, TA),多巴胺(Dopamine, DA)以及5-羟色胺(Serotonin, 5HT)和组胺(Histamine)。许多研究发现昆虫在不同的外界刺激下,体内的生物胺浓度会有显著变化。例如,当柞蚕Antheraeapernyi受到外界刺激时,章鱼胺会促进昆虫的脂肪体进行糖解作用,使血液中的海藻糖浓度上升,以满足能量需求应付紧急的逆境状况(潘灿平等,2005)。热刺激处理下,果蝇Drosophilavirilis、蝗虫Schistocercagregaria与美洲蜚蠊Periplanetaamericana体内的的章鱼胺显著上升(Davenport and Evans, 2010; Maetal., 2015)。
假死行为是大多数昆虫重要的应激反应,然而,这一行为是否受生物胺调控依然未知。Miyatke(2008)检测到了赤拟谷盗脑部生物胺表达量变化活跃度高和低的两个种群假死状态下的多巴胺表达量变化有差异,活跃度大的赤拟谷盗多巴胺含量低;Nakayama(2012)和Uchiyama(2019)经过实验获得了类似结果,假死时间长的赤拟谷盗脑内多巴胺含量低。但是,Matsumura(2016)年测定了赤拟谷盗脑部4种生物胺(章鱼胺、多巴胺、5-羟色胺、N乙酰碱胺),未发现生物胺和活跃度有相关性。
生物胺及其受体是否对假死行为有调控作用(Miyatakeetal., 2008b),为假死行为发生机制的研究提供了新思路。目前,已经在很多昆虫中克隆到了编码章鱼胺受体OARs(Octopamine receptors)和酪胺受体TARs(Tyramine receptors)的核苷酸序列,包括黑腹果蝇Drosophilamelanogaster、桔小实蝇Bactroceradorsalis(Robyn and Schafer, 2009; Claßen and Scholz, 2018)等。该类受体调控生物体多种至关重要的生理学功能,关于其分子生物学特性以及药剂靶标潜力的研究颇多。如果找出假死行为和生物胺的联系,克隆出其受体基因,从基因层面探索假死的发生机制,必将是一项重要突破。
3.2 神经调控
关于昆虫假死行为的神经调节作用的研究,一直未有较大进展。昆虫中,切除车轴草籽象头部后,虫体不发生假死(Duporte and Melville, 1916),但这无法充分证明假死由神经中枢调控;又或许昆虫中的剑鞘器(FCOs)直接参与了假死刺激的诱导(Nishino and Sakai, 1996; 孟凡明和梁醒财,2009),但与之相关的神经中枢的活动情况较模糊。
然而,在以黑鲨等其他物种为研究对象的实验中,认为交感神经系统(VVS)或副交感神经系统主要参与调控(Klemm, 1977; Davieetal., 1993; Albonietal., 2008)。此外,基于光遗传学和电生理学研究,已验证SC-丘脑枕核(Pulvinar nucleus)-杏仁核(Amygdala)皮层下通路介导小鼠的假死反应(Weietal., 2015; Münchetal., 2020),详细解析了假死行为发生时的特定神经元的细胞机制及神经环路机制。这些关于假死行为的神经生物学研究可为从脑神经层面探讨昆虫假死行为神经调控机制提供思路启发和理论参考,有助于进一步分析昆虫假死行为发生机制。
3.3 遗传基础
假死行为同其他行为(捕食、交配、产卵等)相同,均具有遗传基础(Miyatakeetal., 2008a; Miyatakeetal., 2019),其行为特征同时受到遗传和环境两方面的影响(Nakayama and Miyatake, 2009; Nakayama and Miyatake, 2010)。Miyatake等人的人工选择实验中,实验室培养8代以上的绿豆象和赤拟谷盗,和不同地域采集的绿豆象对比实验中,假死特性、飞行能力和假死强度的负相关性大体一致,表明假死具有遗传性(Ohno and Miyatake, 2007; Matsumuraetal., 2016),从基因层面探索昆虫假死行为或是今后研究的必由之路。
不同物种之间的假死强度有很大的不同,如假死时间普遍有差异,假死时间几秒到几小时不等。竹节虫可以伪装小树枝几个小时,负子蝽假死时间也长达1 h,而长蝽科的若虫仅有几秒的假死(孟凡明和梁醒财,2010; Farkas, 2016)。同种昆虫不同个体间的假死时间也存在差异,甘薯象甲雌虫假死12 min,雄虫18~25 min,其原因可能是假死异源进化(孟凡明和梁醒财,2009; Ritteretal., 2016)。
截止目前,国内外尚无关于假死行为和遗传进化关系的明确报道。
4 结语与展望
4.1 小结
假死行为是昆虫重要的反捕食防御行为,和生存策略密切相关。近年来,与昆虫假死行为相关的研究逐渐增多,明确了多种昆虫的发生时间、发生频率、刺激方式和诱发因素等特征;但昆虫假死行为的发生机制仍未明确。关于假死行为研究的存在问题总结如下:
具有假死现象的昆虫种类多但被深入研究种类少:自然界中,有大量昆虫存在假死行为。目前的研究对象仅有赤拟谷盗、甘薯象甲、绿豆象、臭椿沟眶象等40余种。研究更多具有假死行为的昆虫,将有助于深入理解昆虫假死行为的特点和发生机制。如何解释昆虫假死行为的普遍性,仍是一项挑战。
研究层次不高:当前研究多集中于昆虫假死行为的发生特征,包括假死强度、诱发因素等,很少涉及研究假死发生机制。常规行为分析手段,比如直接观察、基础行为实验是第一个研究层次,初步解释假死昆虫行为发生反应的基础特征。然而,每一次假死行为,都需要虫体各个系统的互相协调,从而实现假死的发生。从虫体内外系统研究假死行为的发生是第二个研究层次,如外界因素光照、温度、声音、背景色等,到内部因素如虫体大小、雌雄、饥饿探究假死发生的影响,目前多数研究集中在这个层次。运用生理学、神经生物学、分子生物学的技术手段,从生理生化甚至基因层次探究假死行为的发生是第三个研究层次,但鲜有涉及。
研究学者少:关于昆虫的假死行为研究已取得较大进展,但研究学者较少,导致仍有很多基础科学问题尚未涉及,如已知明确具有假死行为的昆虫仅几十种,必然低于自然界中的实际物种数量;赤拟谷盗、绿豆象、竹节虫、臭椿沟眶象等昆虫,有的假死时间最长可达几小时,有的仅几分钟,差异显著。总之,不经大量试验验证,无法直接断言物种是否具有假死行为,其发生机制如何。但是,实际上至今未有理论指导如何直接断定哪种昆虫偏好假死,哪种昆虫不喜假死,哪种昆虫干脆拒绝假死。假死作为昆虫重要的防御行为,希望今后更多的研究学者探索其中奥秘。
4.2 展望
哺乳类、鱼类、鸟类等假死行为的诱发同样受机械刺激、温度、声音等,但是没有明显的假死姿态,较难从外观直接判断是否假死,且实验操作较复杂。与之相比,昆虫的假死行为相对简单,易在实验室进行操作,了解昆虫的假死行为发生特征和机制,有助于深度解析其他动物的假死行为。不同昆虫的假死状态相似,均静止不动,然而即使同一物种间的昆虫,假死发生时间出现较大差异,因此以多种昆虫为对象,对比不同物种的假死行为,有助于深入理解假死行为的发生特征。而假死行为降低被捕食率、增加逃逸率、提高自身适合度,和生存策略有密切联系,未来从进化生态学角度研究假死行为发生和生存策略之间的联系也很有价值。另外,具有假死行为昆虫,如臭椿沟眶象、甘薯甲虫、尺蠖Geometridae、赤拟谷盗等,是重要的林业和农业害虫,对我国的农林生产和社会经济造成严重损失。文献报道利用昆虫假死或可防治害虫(张霞等,2001;张民照等,2009),如通过全基因转录组分析和RNAi干扰技术筛选与假死行为相关的基因,开发调控剂,干扰这类害虫的生理和行为,达到防治目的。还有学者提出,未来或可人为控制假死发生时间,进行活体运输(Brooksetal., 2011)。Claudia等认为细胸金针虫近缘种间假死时间差异显著,可用于分类鉴定(Farkas, 2016; Ritteretal., 2016)。
近几年,现代新技术的发展开辟了昆虫假死行为发生机制的研究新途径,利用纳米级电镜和CT可以探索感器分布脑结构等与假死行为神经控制相关的组织,LC-MS/MS、RNAi干扰以及基因编辑等新技术实现了从生理学和分子生物学角度探索基因对假死行为的调控的可能。综合运用生理学、神经生物学、分子生物学的技术手段,从假死行为的发生特征入手,重点阐述假死行为发生时的各系统内分泌激素和各组织器官间的基因转录、蛋白质翻译的变化,以及各神经系统之间的调节机制,并深入探索这一行为的遗传进化基础,相信不久将来假死行为的发生机制最终会被揭晓。
