长期增温对短花针茅荒漠草原3种主要植物光合日动态特性的影响
2021-03-25雷雪峰
雷雪峰,王 妍,李 杨,梁 燕,白 龙
(内蒙古农业大学农学院,内蒙古呼和浩特 010019)
按照政府间气候变化专门委员会(IPCC)的预测结果,到21 世纪末,全球平均气温将升高1.5~4.8℃[1],全球气候变暖将对植物、动物、微生物及各生态系统功能产生怎样的影响,早已引起相关学者的广泛关注[2-5]。植物作为陆地生态系统的重要组成部分,对全球气候变化的响应最为突出和显著[6]。研究植物对气候变化的响应与适应对策,对了解植物演化、预测气候变化条件下植物的变化特征对生态系统的影响具有重要意义[7-8]。关于预测气候变化对生态系统影响的研究,目前,涉及的气候变化影响因子以温度、氮沉降、CO2、水分、酸雨[9-11]等为主。温度是影响植物生长、发育和功能的重要环境因子,温度变化不仅会影响地表植被的种群及群落结构,还会影响各种植物的生长发育与繁衍。温度对植物的直接影响主要表现在植物的光合、呼吸、代谢等生理方面,从而改变植物的生长速率及物候等特性。针对全球变暖对陆地生态系统的影响,近年来,生态学者们开展了大量关于增温的试验研究,研究尺度主要集中在对生态系统气体交换、群落结构与功能、土壤特性、物种组成等宏观尺度上,对植物方面的研究主要是植物物候及群落特征、生产力变化等[12-14]。关于植物光合特性对增温响应的研究报道相对较少。
位于内蒙古中部、阴山山脉以北乌兰察布高原地区的荒漠草原是我国北方重要的生态屏障,也是内蒙古草原的重要组成部分,其独特的地理分布以及物种组成已成为亚洲中部一种特殊的、十分重要的生态系统类型。在这种环境影响下形成的荒漠草原生态系统极其脆弱,对人类活动和全球气候变化极其敏感。荒漠草原生态系统一旦遭到破坏则难以恢复,具有发生荒漠化的潜在危险[15]。因此,研究气候变暖对荒漠草原植物的影响及生理机制,对保护内蒙古西部地区的草地生态系统和畜牧业可持续发展具有重要意义。本试验以内蒙古短花针茅荒漠草原3种主要植物——短花针茅(Stipa breviflora)、冷蒿(Artemisia frigida)、木地肤(Kochia prostrata)为研究对象,采用野外模拟控制性增温的研究方法,测定短花针茅荒漠草原3种主要植物的光合特性日动态,分析长期增温对荒漠草原主要植物产生的影响及规律,旨在为今后解释和预测未来全球气温变化对荒漠草原植物光合特性的影响提供基础理论依据。
1 材料和方法
1.1 试验地概况
试验地位于内蒙古自治区乌兰察布市四子王旗内蒙古自治区农牧业科学院科研基地(41°47′17″N,111°53′46″E),海拔1 456 m。该地区多年平均气温为3.7℃,多年平均降水量为311 mm,降水主要集中在6—9月。年均水分蒸发量约为2 947 mm,气温平均日较差为13~14℃,≥10℃年积温为2 200~2 500℃,平均无霜期为90~120 d,平均日照时数为3 117.7 h。该试验区属于短花针茅荒漠草原地带性植被,草地类型为短花针茅+冷蒿+无芒隐子草(Cleistogenes songorica)。建群种为短花针茅,优势种为冷蒿、无芒隐子草,主要伴生种有木地肤、银灰旋花(Convolvulus ammannii)、阿尔泰狗娃花(Heteropappus altaicus)、栉叶蒿(Neopallasia pectinata)、狭叶锦鸡儿(Caragana stenophylla)和羊草(Leymus chinensis)等。
1.2 试验设计
试验基地的增温装置2006年5月安装,常年持续使用。本试验是在试验样地长期增温的基础上于2017年进行的测定。试验采用完全随机裂区设计,样地设增温和不增温(对照)2种处理,设置长3 m、宽2 m的样方,每个处理6次重复,共12个样方,面积共72 m2。在各处理样方间,设置了3 m×3 m的缓冲带,以消除试验过程中的人为破坏。6个增温区内各安装一个红外线辐射器进行加热。辐射器规格为165 cm×15 cm,安置于增温样方上部,距地面2.25 m,辐射器功率设置为2 000 W,全年不间断加热。每个对照区安置“假灯”消除辐射器遮阴等造成的误差。
1.3 试验材料
试验选取样地内有代表性的3种植物进行光合日动态测定,分别为建群种短花针茅、优势种冷蒿和主要灌木木地肤。
1.4 植物光合日动态测定指标及方法
试验时间为2017年7月,选择晴朗无风天气,使用便携式光合仪Li-6400(LI-COR lnc.Nebraska,USA)测定各植物的光合参数。选用开放气路,自然光源下进行测定。在每个测定样方内选择3株健康、成熟的植株叶片进行测定,每株3次重复。从6:00 开始,每2 h 测定1次,20:00 结束,每种植物各测定7次。主要测定指标包括光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci),并计算水分利用效率(WUE)。
水分利用效率(WUE)的计算公式
WUE=Pn/Tr
式中,Pn表示光合速率;Tr表示蒸腾速率。
1.5 数据处理方法
采用Excel 2016 软件对数据进行整理、计算分析、制作曲线图,采用SPSS 22.0 软件制作柱形图及进行差异显著性分析。
2 结果与分析
2.1 增温对荒漠草原主要植物光合速率日动态的影响
由图1、图2、图3可知,短花针茅、冷蒿和木地肤3种植物的光合速率日动态具有相同的变化趋势,即均为双峰曲线,基本为上午峰值高于下午峰值。3种植物的光合速率日动态对增温的响应存在一定差异,其中,短花针茅光合速率日动态对增温的响应基本为整体增加;冷蒿光合速率日动态对增温的响应略有降低,与对照(不增温)相比,上午波峰后移变宽,下午曲线波动趋势基本一致;木地肤光合速率日动态对增温的响应表现为上午波峰升高,下午波峰降低,并且光合日动态曲线整体向左推移。
从光合速率日动态均值来看(图4),3种植物光合速率日动态均值存在明显差异,其大小关系为短花针茅>木地肤>冷蒿。其中,短花针茅与木地肤光合速率日动态均值差异不显著(P>0.05),冷蒿与短花针茅、木地肤光合速率日动态均值差异显著(P<0.05)。与对照相比,增温提高了短花针茅和木地肤的光合速率日动态均值,但差异不显著(P>0.05)。增温对冷蒿光合速率日动态均值的影响极小。
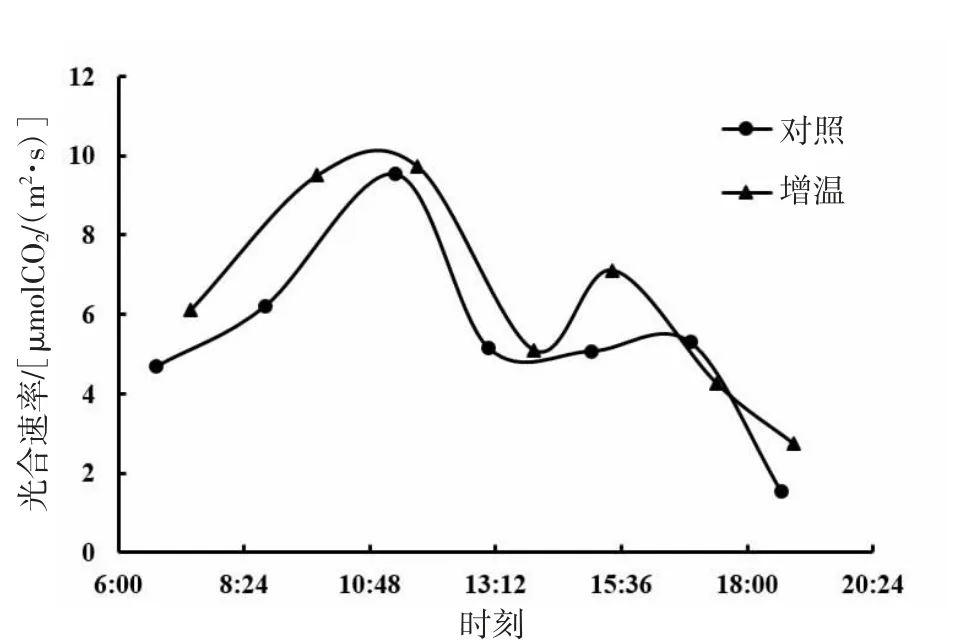
图1 短花针茅光合速率日动态

图2 冷蒿光合速率日动态
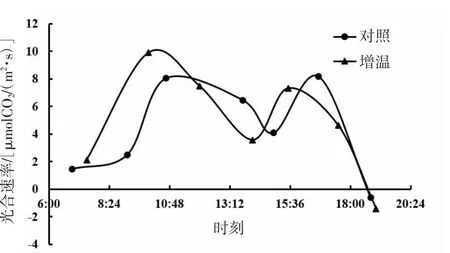
图3 木地肤光合速率日动态

图4 各植物光合速率日动态均值比较
2.2 增温对荒漠草原主要植物蒸腾速率日动态的影响
由图5、图6、图7可知,短花针茅、冷蒿和木地肤3种植物的蒸腾速率日动态基本具有相同的变化趋势,即均为双峰曲线。3种植物的蒸腾速率日动态对增温的响应存在一定差异,其中,短花针茅和木地肤的蒸腾速率日动态对增温的响应为明显整体增加,且木地肤蒸腾速率日动态曲线波峰提高尤为明显;冷蒿蒸腾速率日动态曲线对增温的响应与另两种植物相比差异最大,上午蒸腾速率基本保持一个水平不变,下午较上午明显降低,但也保持在一个较稳定的水平上。
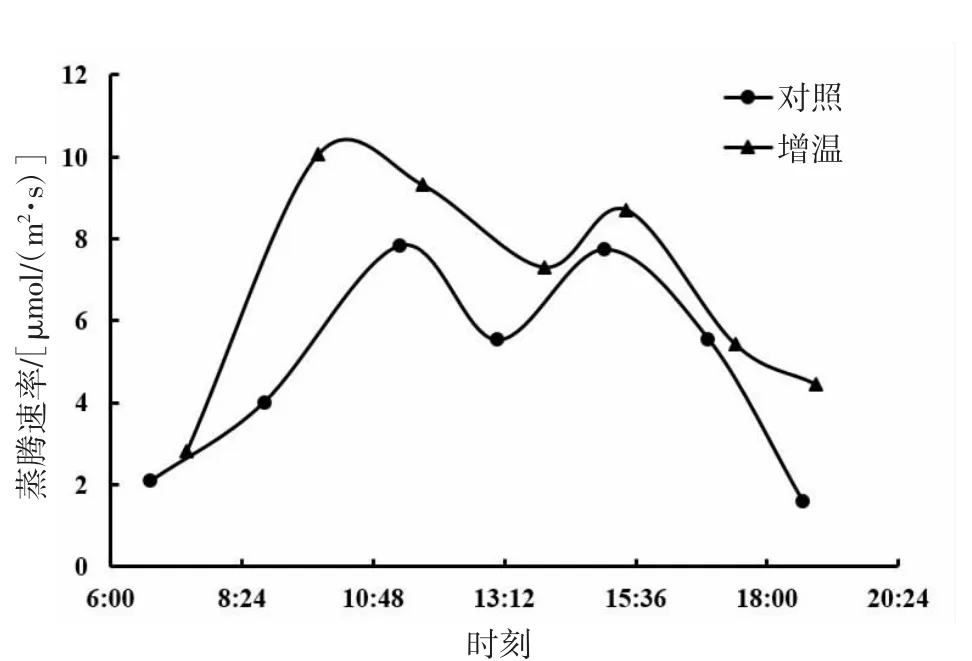
图5 短花针茅蒸腾速率日动态
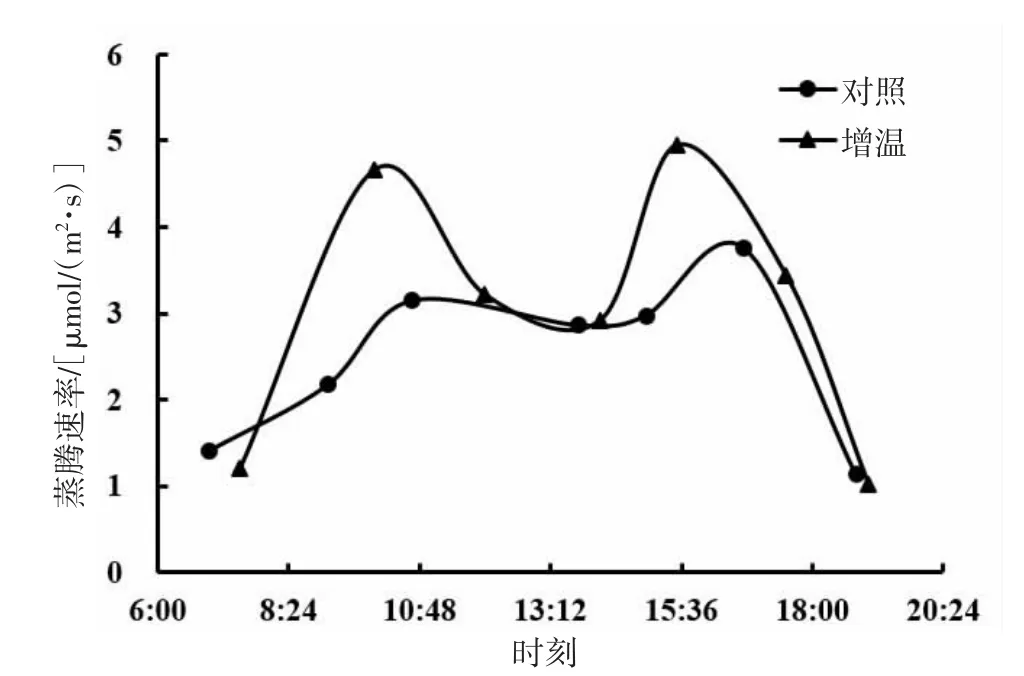
图6 冷蒿蒸腾速率日动态

图7 木地肤蒸腾速率日动态
从蒸腾速率日动态均值来看(图8),短花针茅的蒸腾速率日动态均值明显大于木地肤和冷蒿,且差异达到显著水平(P<0.05)。冷蒿与木地肤蒸腾速率日动态均值大小接近,差异不显著(P>0.05)。与对照相比,增温均提高了3种植物的蒸腾速率日动态均值,其中,短花针茅的蒸腾速率日动态均值增加最明显,达到了显著水平(P<0.05);冷蒿及木地肤蒸腾速率日动态均值增幅较小,差异不显著(P>0.05)。
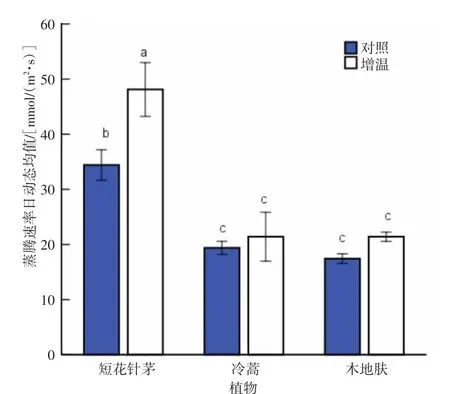
图8 各植物蒸腾速率日动态均值比较
2.3 增温对荒漠草原主要植物胞间CO2浓度日动态的影响
由图9、图10、图11可知,短花针茅、冷蒿和木地肤3种植物的胞间CO2浓度日动态具有相同的变化趋势,基本呈现“W”形曲线。3种植物的胞间CO2浓度日动态对增温的响应存在一定差异,其中,短花针茅胞间CO2浓度日动态对增温的响应表现为两端较低,冷蒿胞间CO2浓度日动态对增温的响应为中段右移,木地肤胞间CO2浓度日动态对增温的响应为中段提升。整体来看,增温对3种植物的胞间CO2浓度日动态影响幅度不大。
从胞间CO2浓度日动态均值来看(图12),3种植物胞间CO2浓度日动态均值存在一定差异,大小为冷蒿>短花针茅>木地肤,其中,短花针茅与冷蒿胞间CO2浓度日动态均值差异不显著(P>0.05),木地肤与短花针茅、冷蒿胞间CO2浓度日动态均值差异显著(P<0.05)。增温降低了短花针茅的胞间CO2浓度日动态均值,提高了冷蒿和木地肤的胞间CO2浓度日动态均值,但差异均未到达显著水平(P>0.05)。

图9 短花针茅胞间CO2浓度日动态
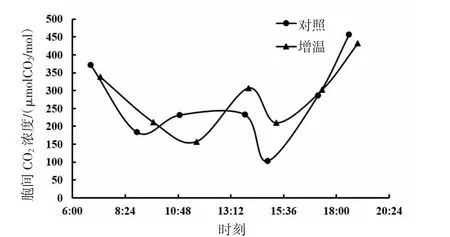
图10 冷蒿胞间CO2浓度日动态

图11 木地肤胞间CO2浓度日动态

图12 各植物胞间CO2浓度日动态均值比较
2.4 增温对荒漠草原主要植物气孔导度日动态的影响
由图13、图14、图15可知,短花针茅、木地肤的气孔导度日动态具有相同的变化趋势,基本呈现为双峰曲线;冷蒿的气孔导度日动态与前者相比差异较大。3种植物的气孔导度日动态对增温的响应存在一定差异,其中,短花针茅气孔导度日动态对增温的响应表现为两端较低,冷蒿气孔导度日动态对增温的响应为中段右移,木地肤气孔导度日动态对增温的响应为中段提升。整体来看,增温对3种植物的气孔导度日动态影响幅度不大。
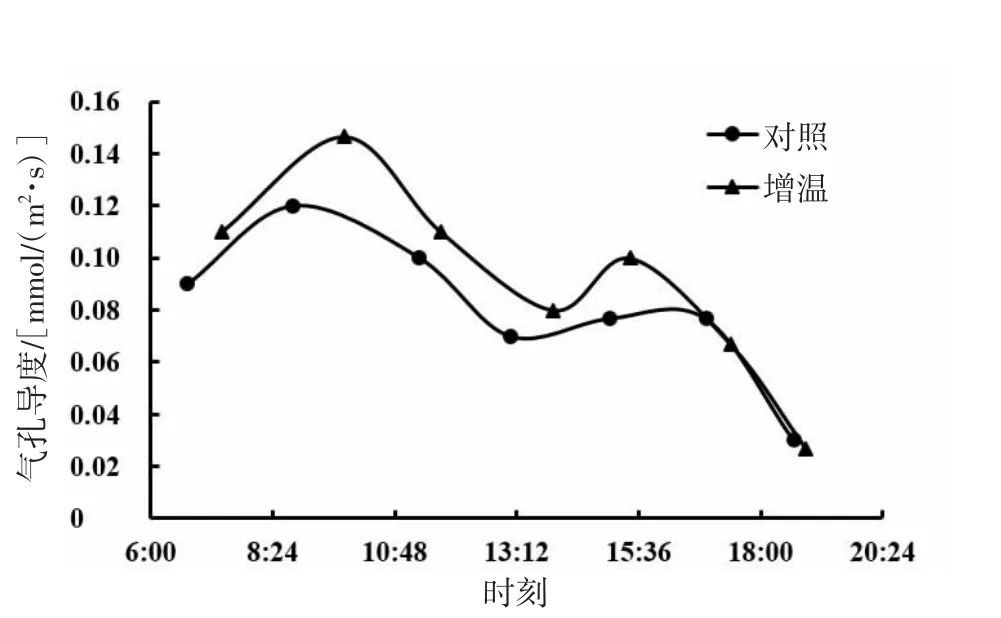
图13 短花针茅气孔导度日动态

图14 冷蒿气孔导度日动态

图15 木地肤气孔导度日动态
从气孔导度日动态均值来看(图16),3种植物中短花针茅的气孔导度日动态均值最大,与木地肤、冷蒿气孔导度日动态均值差异达到了显著水平(P<0.05);木地肤与冷蒿的气孔导度日动态均值接近,且差异不显著(P>0.05)。增温提高了短花针茅和冷蒿的气孔导度日动态均值,短花针茅增幅大于冷蒿,但差异均未到达显著水平(P>0.05)。
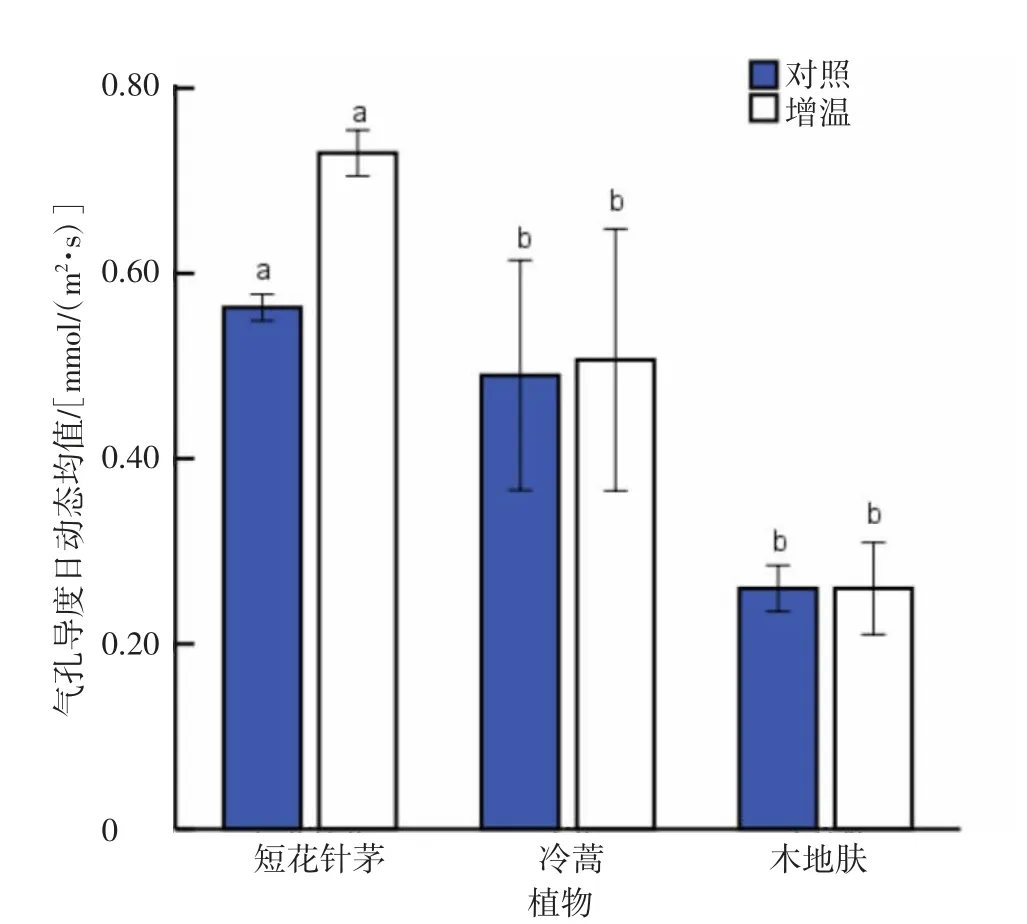
图16 各植物气孔导度日动态均值比较
2.5 增温对荒漠草原主要植物水分利用效率日动态的影响
由图17、图18、图19可知,短花针茅、冷蒿和木地肤3种植物的水分利用效率日动态具有相同的变化趋势,基本呈现为双峰曲线。3种植物的水分利用效率日动态对增温的响应基本表现为降低的趋势,但不同植物间也存在一定差异。其中,短花针茅的水分利用效率日动态对增温的响应表现为上午明显较低,下午略有降低;冷蒿水分利用效率日动态对增温的响应为上午略有增加,下午明显降低;木地肤水分利用效率日动态对增温的响应为上、下午均降低。
从水分利用效率日动态均值来看(图20),3种植物水分利用效率日动态均值存在一定差异,大小为木地肤>短花针茅>冷蒿。其中,短花针茅与冷蒿水分利用效率日动态均值差异不显著(P>0.05),木地肤与短花针茅、冷蒿水分利用效率日动态均值差异显著(P<0.05)。增温处理均降低了短花针茅、冷蒿和木地肤的水分利用效率日动态均值,但差异均未到达显著水平(P>0.05)。
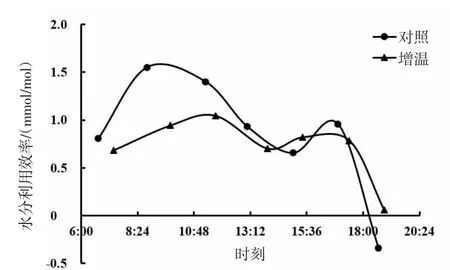
图17 短花针茅水分利用效率日动态
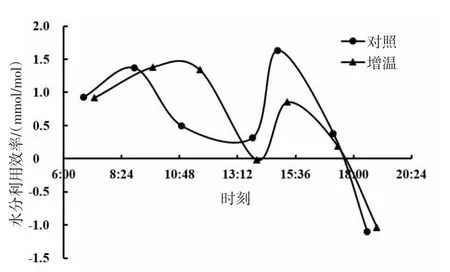
图18 冷蒿水分利用效率日动态
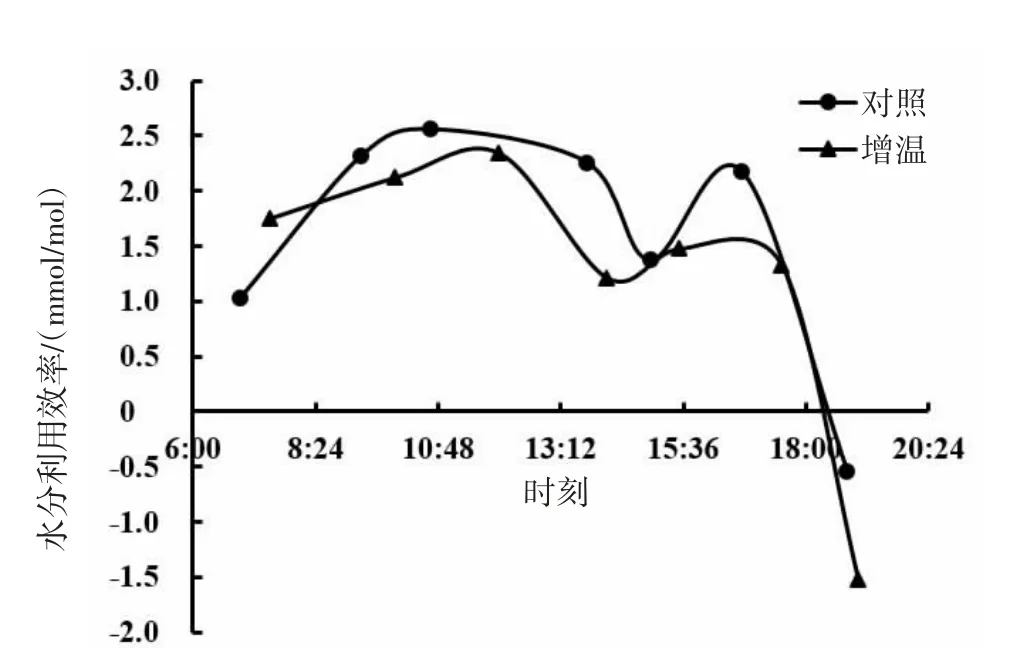
图19 木地肤水分利用效率日动态
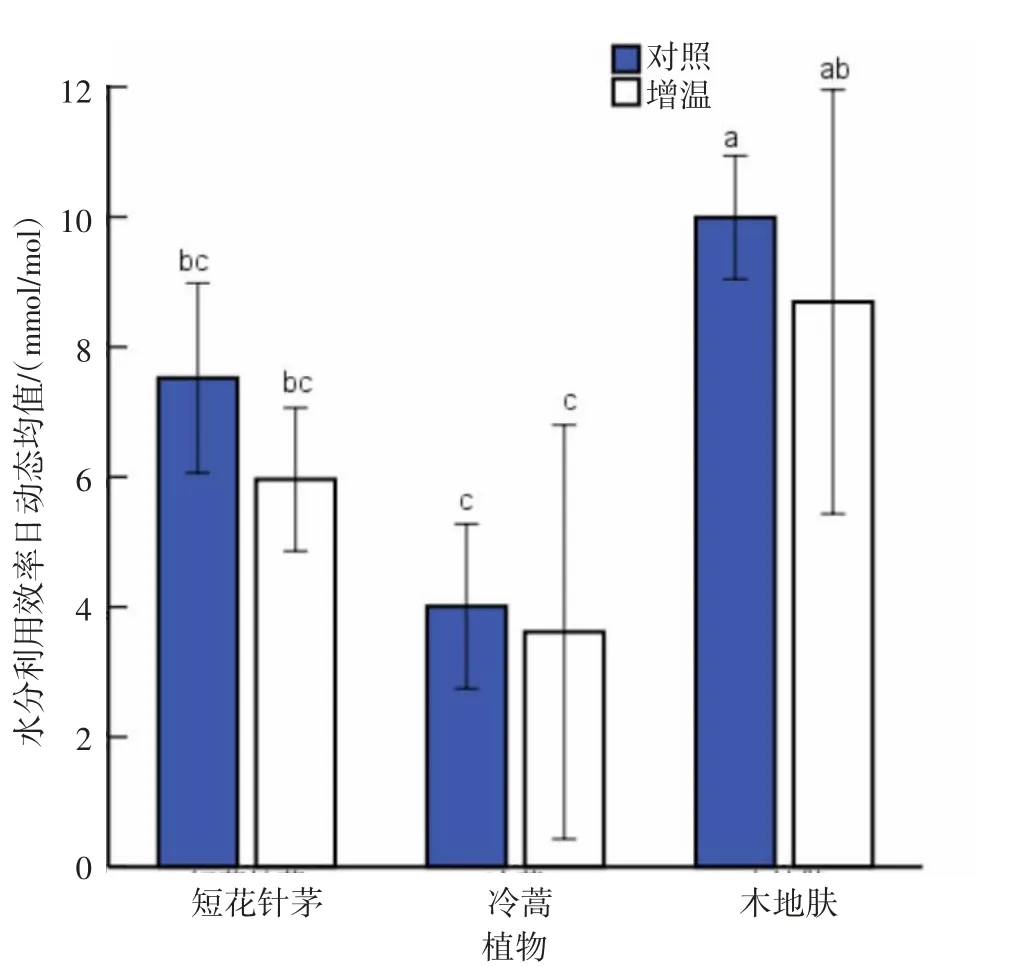
图20 各植物水分利用效率日动态均值比较
3 讨论
植物的光合作用是一系列酶促反应过程,其效率的高低受温度的影响较大。植物只有处于最适温度附近时,光合速率才能达到最大,低于或高于这个最适温度时光合速率都会降低。即使生长在同一环境下的不同植物,其遗传背景不同,光合特性的最适温度也存在明显差异。加之水分、养分、土壤微生物等其他环境因子的影响,更是加大了分析植物对增温响应规律的难度。目前,学术界关于增温对植物光合特性影响规律的研究存在较大争议。其结果可概括为3种类型:其一,增温促进了植物的光合作用[16];其二,增温抑制了植物的光合作用[17-18];其三,增温对植物的光合作用无明显影响[19]。出现这样的不同研究结果,可能与植物的不同分布、植物种类以及不同的增温强度等因素有关。因此,有必要继续对不同地理分布的不同植物种类进行系统科学的增温模拟试验,为进一步揭示导致植物对增温产生不同响应的原因提供基础依据。本试验为了探讨内蒙古荒漠草原植物光合特性对增温的响应,选择了3种具有代表性的典型植物进行光合特性日动态测定,结果显示,3种植物光合速率日动态、蒸腾速率日动态及水分利用效率日动态均显示为双峰曲线,胞间CO2浓度日动态表现为“W”形曲线。增温处理后,短花针茅和木地肤光合特性日动态曲线趋势变化幅度较小,与对照曲线相比,只是出现了上下或左右的波动,没有出现明显的曲线趋势改变;与短花针茅和木地肤相比,冷蒿光合特性日动态曲线趋势受增温影响相对较大,尤其是蒸腾速率日动态曲线趋势,在增温的影响下彻底改变了双峰曲线模型,表明增温扰乱了冷蒿的正常蒸腾规律。
温度可以通过多个方面影响植物叶片的光合特性,如温度会对光合酶活性、类囊体膜功能、叶绿素含量、胞间CO2浓度及气孔导度等产生影响。不同的植物种类遗传背景不同,对环境温度的适应方式各不相同,有时同样的增温强度可导致不同植物产生不同的响应。本试验显示,不同植物种类,光合特性日动态平均值对增温的响应趋势存在一定的差异,基本表现为短花针茅对增温处理响应最大,表现为促进的变化趋势;增温对木地肤、冷蒿的光合特性也产生了一些积极的影响,但影响程度较小。
有学者研究表明,随着叶片温度的升高,植物的气孔导度下降、叶片蒸腾速率下降、叶肉胞间CO2浓度升高,水分利用率增加[20-21]。但本试验结果显示,增温基本提高了短花针茅荒漠草原3种主要植物短花针茅、冷蒿、木地肤的光合速率,这可能与提高植物的气孔导度有直接关系。增温也提高了短花针茅荒漠草原3种主要植物短花针茅、冷蒿、木地肤的蒸腾速率,同时降低了植物的水分利用效率。这与前人的研究结果差异较大,原因可能与模拟增温的强度、植物类型等因素有关。另外,影响植物光合特性的气候因子很多,尤其是当地的降雨频率及强度随着年份、季节的变化会有较大差异,必然影响植物光合特性对增温的响应。因此,在不同的生长期及不同年份对植物进行光合特性的测定,对于掌握增温条件下植物光合特性的响应规律具有更重要的意义。
4 结论
长期增温对短花针茅荒漠草原3种主要植物短花针茅、冷蒿、木地肤光合特性影响较小。不同植物对增温的响应存在一定差异,其中,增温对短花针茅光合特性的影响相对较大、冷蒿光合特性日动态曲线对增温的响应相对较为明显。
