短花针茅叶片热值及还原糖含量对载畜率的响应
2021-11-03郑佳华赵萌莉
郑佳华,高 峰,张 峰,张 彬,赵萌莉
(1.内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010010;2.内蒙古自治区水利科学研究院,内蒙古自治区,内蒙古 呼和浩特 010051)
草地生态系统占全球陆地总面积的16.4%,在人类生产生活中起着举足轻重的作用,不仅可以为人类提供各种畜产品,还具有调节气候的作用[10]。放牧是草原常见的利用方式之一,由于长期过度放牧,导致草原生态系统逐渐退化。目前,已有研究发现放牧会不同程度地破坏草原,比如导致草地生物量[11]、物种多样性[12]和土壤肥力下降[13]等。但也有研究表明,适度放牧可以提高草地生态系统的净初级生产力和物种多样性[14]。位于内蒙古四子王旗的荒漠草原处于我国农牧交错带,是草原向荒漠的过渡性草原。由于受到气候和过度放牧等因素的影响,生态系统脆弱,抗干扰能力差,植物多样性低。目前,关于该区的草地群落特征、生产力、物种多样性及土壤养分的研究较多[12,15-16],但是对建群种短花针茅(Stipa breviflora)叶片热值及还原糖的研究相对较少。探究放牧对短花针茅叶片热值和还原糖含量的影响可以深入理解不同载畜率水平下短花针茅光合能力和能量流动,探索如何通过合理的放牧制度保护该系统的建群植物种,提高生态系统的稳定性。因此,本研究以内蒙古荒漠草原建群种短花针茅为研究对象,分析其叶片热值、还原糖和干物质对载畜率的响应规律以及相互关系,旨在为科学利用和保护草地资源提供理论指导。
1 材料与方法
1.1 试验区概况
试验区位于内蒙古自治区乌兰察布市四子王旗内蒙古农牧科学院草原研究所实验基地(111°53′46″ E,41°47′17″ N;海拔约1 456 m),为典型的中温带大陆性季风气候,夏季炎热干旱、少雨,冬季大风、寒冷,四季更替明显;年均降水量约253 mm,多集中在植物生长季(5月 − 9月),约占全年降水量的80%;年均气温3.6 ℃,最高温在7月 − 8月,约为24 ℃,最低温在1月,约−17 ℃;全年无霜期约为150 d。土壤主要为淡栗钙土,且具沙壤土结构[17]。试验区为典型的荒漠草原,植被低矮,草层稀疏,植被盖度低,以短花针茅为建群种,以无芒隐子草(Cleistogenes songorica)和冷蒿(Artemisia frigida)为主要优势种,常见伴生植物种有银灰旋花(Convolvulus ammannii)、阿尔泰狗娃花(Heteropappus altacius)、栉叶蒿(Neopallasia pectinata)、木地肤(Kochia protrata)、小叶锦鸡儿(Caragana microphylla)和狭叶锦鸡儿(Caragana stenophylla)等[12]。
1.2 试验设计
试验基于内蒙古四子王旗荒漠草原长期放牧监测平台开展,该平台成立于2004年。试验区建立起初为天然草地,总面积50 hm2,采用完全随机区组设计,设置不放牧和3 种载畜率水平处理,分别为每月0、0.15、0.30 和0.45 个羊单位·hm−2。放牧家畜选用当地成年绵羊,4 个处理分别为对照(control,CK)、轻度放牧(light grazing,LG)、中度放牧(moderate grazing,MG) 和重度放牧(heavy grazing,HG),每处理3 个重复。不同载畜率处理草地的利用率分别为0、30%、42%和67%,重度放牧处理的草地利用率是当地牧民目前对草地的放牧利用情况[18]。
1.3 样品采集与处理
野外试验于2018年4月 − 9月中旬进行。每一试验区内分别设置3 个1 m × 1 m 的样方,将样方内的短花针茅植株全部齐地面刈割,带回实验室,105 ℃杀青30 min 后,65 ℃烘24 h 至恒重,用天平秤其干物质重。将烘干的短花针茅叶片挑出并用球磨仪粉碎研磨,用于测定短花针茅叶片的热值和还原糖含量。
短花针茅叶片热值的测定:采用C1 Package 1/10(IKA,Germany) 全自动量热仪,测定时,称取约1 g干样品,放置于弹氧装置中,将点火线拴在点火丝上,然后点火使样品充分燃烧,测定植物叶片完全燃烧时释放的热量。
叶片热值(kJ·g−1) = 充分燃烧释放的热量/称取的干样品重量。
叶片还原糖含量用3,5-二硝基水杨酸法测定[19]。
1.4 数据处理与分析
使用Excel 2013 整理数据,使用SPSS 23.0 进行双因素方差分析、Duncan 多重比较和回归分析,使用Sigmaplot 14.0 制图。
2 结果与分析
2.1 短花针茅叶片热值的变化
载畜率和月份分别对短花针茅叶片热值的影响极显著(P< 0.001),但载畜率与月份对短花针茅叶片热值影响的交互作用不显著(P> 0.05) (表1)。
湖南浏阳的雨季出现在5月下旬,为避免降雨高峰时段出现在施肥后前期,故水样的采集时间为施基肥后的第1、2、3、5、7 天,追肥后的第1、2、3、5天的8:30—9:30,以50 mL医用注射器采集田面水(李瀚等,2015),在不扰动土层的前提下,按S型5点取样,混合后注入250 mL集水瓶,每个处理分取3份,将其pH调为1~2后于-20 ℃冷冻保存,待测(HJ 636—2012)。
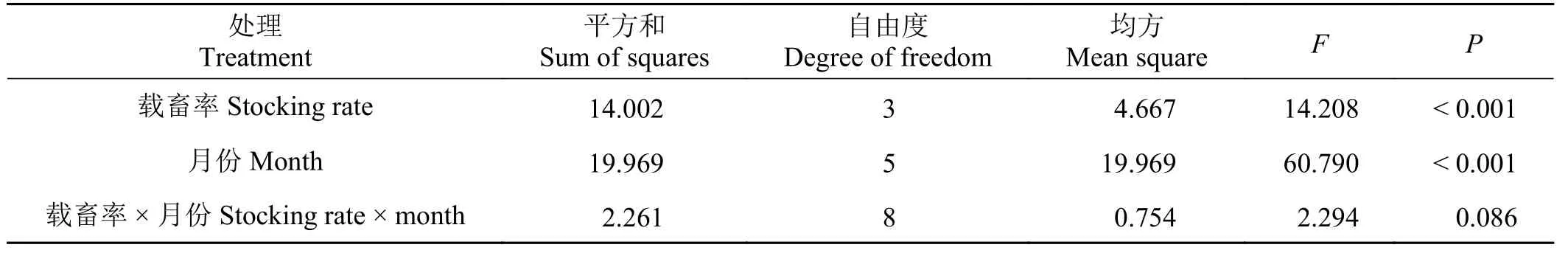
表1 载畜率对短花针茅叶片热值的方差分析Table 1 F-value of RMANOVA for the effects of stocking rate on gross calorific values of Stipa breviflora leaf
随着载畜率的增加,短花针茅叶片热值逐渐下降,对照与轻度放牧、中度与重度放牧处理之间差异不显著(P> 0.05),但对照和轻度放牧显著高于中度和重度放牧处理下的短花针茅叶片热值(P<0.001)。从4月 − 9月,短花针茅叶片热值表现为逐渐上升的趋势,4月的短花针茅叶片热值为16.52 kJ·g−1,9月的为19.07 kJ·g−1,9月的短花针茅叶片热值显著高于4月−7月的叶片热值(P< 0.05),与8月的叶片热值差异不显著(P> 0.05) (图1)。

图1 载畜率和月份对短花针茅叶片热值的影响Figure 1 The effects of stocking rate and month on gross calorific values of Stipa breviflora leaf
2.2 短花针茅干物质量的变化
短花针茅的干物质量极显著受到载畜率和月份的影响(P< 0.01),且受载畜率与月份的交互作用影响显著(P< 0.05) (表2)。
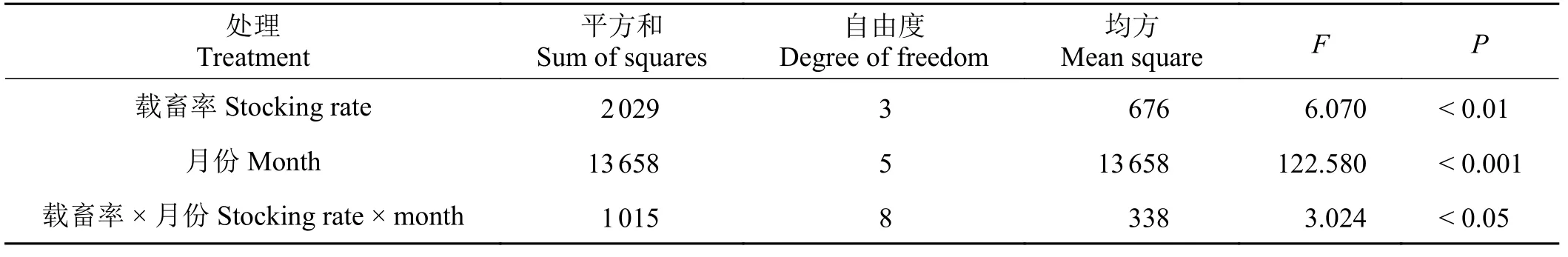
表2 载畜率对短花针茅干物质的方差分析Table 2 F-value of RMANOVA for the effects of stocking rate on dry matter values of Stipa breviflora
在短花针茅返青初期(4月),不同载畜率水平下的干物质重量差异不显著(P> 0.05)。5月 − 8月,短花针茅干物质变化趋势相同,随着载畜率增加,短花针茅干物重量先增加后减少,表现为LG > CK >MG > HG,且差异显著(P< 0.05)。9月份随着载畜率增加短花针茅干物质逐渐减少,CK > LG > MG >HG (P< 0.05)。从4月 – 9月,短花针茅干物质重量均表现为逐渐增加,8月和9月显著高于4月 − 7月的干物质量(P< 0.05)。在围封和中度放牧处理下,9月短花针茅干物质重达到最大值,分别为55.0和37.6 g·m−2。轻度放牧和重度放牧样地在8月短花针茅干物质最重,分别为64.25 和30.91 g·m−2(图2)。
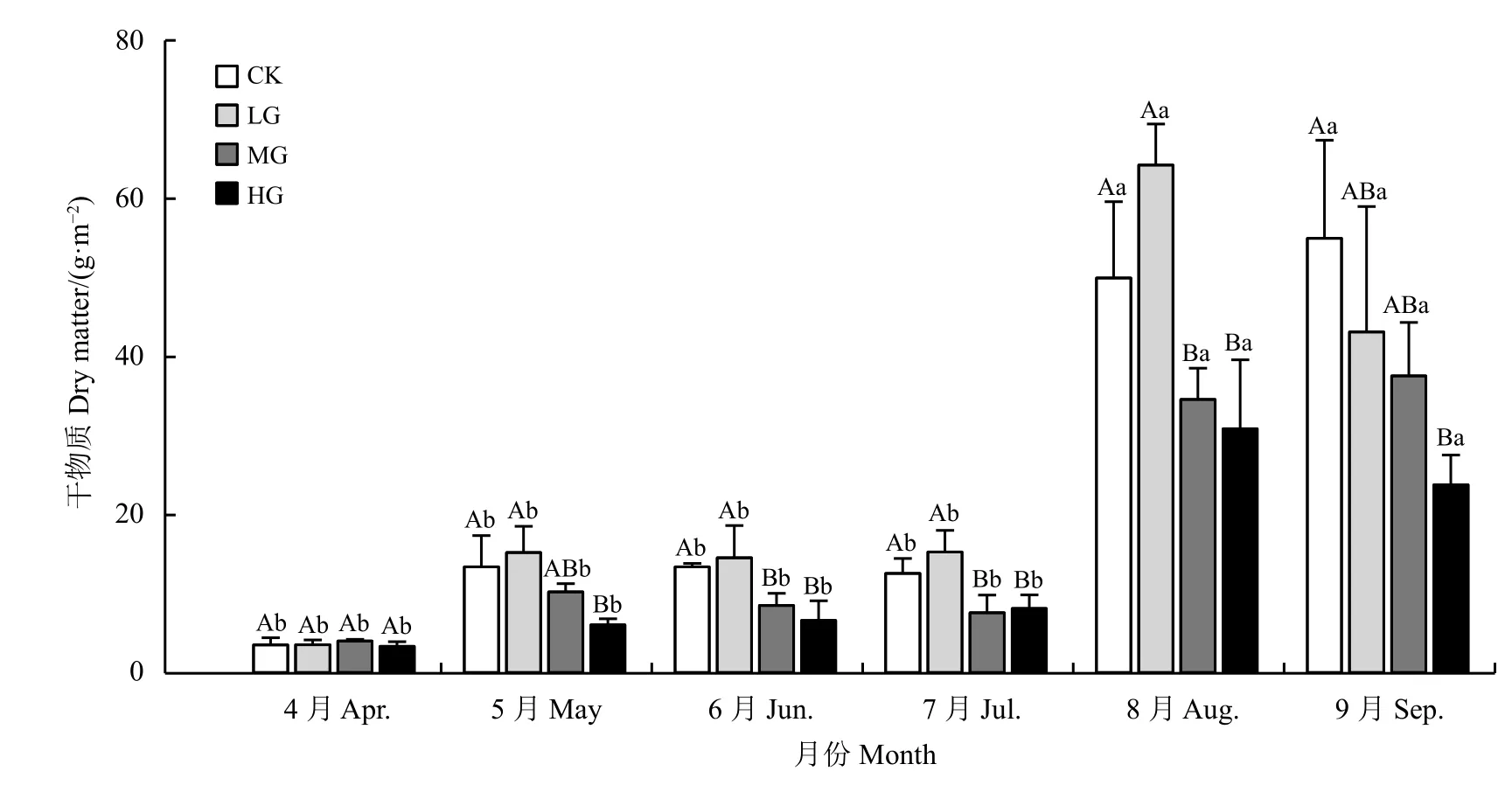
图2 载畜率和月份对短花针茅干物质的影响Figure 2 The effects of stocking rate and month on dry matter values of Stipa breviflora
2.3 短花针茅叶还原糖的变化
载畜率和月份对短花针茅叶还原糖的影响极显著(P< 0.01),且受载畜率和月份的交互作用影响显著(P< 0.05) (表3)。在短花针茅返青初期(4月),不同载畜率水平下叶还原糖含量差异显著(P< 0.05),随着载畜率增加,短花针茅叶还原糖含量逐渐减少,表现为CK > LG > MG > HG。5月、7月和9月短花针茅叶还原糖含量变化趋势相似,放牧显著降低了短花针茅叶还原糖含量(P< 0.05)。6月与8月短花针茅叶还原糖含量差异不显著(P> 0.05) (图3)。
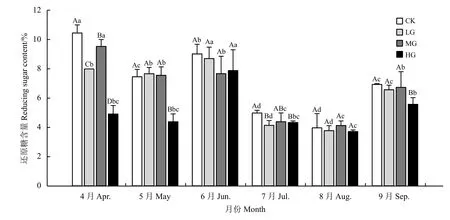
图3 载畜率和月份对短花针茅叶还原糖含量的影响Figure 3 The effects of stocking rate and month on reducing sugar content of Stipa breviflora leaf
表3 载畜率对短花针茅叶还原糖含量的方差分析Table 3 F-value of RMANOVA for the effects of stocking rate on reducing sugar values of Stipa breviflora leaf
4月 − 9月,短花针茅叶还原糖含量均先减少后增加,7月和8月显著低于其他月份的叶还原糖含量(P< 0.05)。在围封和中度放牧处理下,4月短花针茅叶还原糖含量达到最大,分别为10.45%和9.53%。轻度放牧和重度放牧样地在6月时短花针茅叶还原糖含量达到最大值,分别为8.70%和7.88%。
2.4 短花针茅叶热值、干物质和叶还原糖之间的关系
短花针茅干物质与叶片热值呈显著正相关关系(P< 0.001,R2= 0.695),随着干物质量增加,短花针茅叶片热值先逐渐增加,达到一个阈值后逐渐稳定。短花针茅干物质与叶还原糖之间呈显著负相关关系(P< 0.05,R2= 0.128),随着干物质量增加,短花针茅叶片还原糖逐渐减少。短花针茅叶还原糖与叶热值之间不存在显著的相关关系(P> 0.05) (图4)。
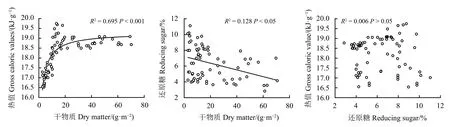
图4 短花针茅叶热值与干物质、叶还原糖与干物质、叶还原糖之间的关系Figure 4 The relationship between leaf gross calorific value and dry matter, reducing sugar and dry matter,and reducing sugar and calorific value of Stipa breviflora
3 讨论与结论
3.1 热值对载畜率的响应
本研究表明,除4月短花针茅返青期以外,5月−9月短花针茅的干物质含量均随载畜率的增加显著降低(P< 0.05)。张爽等[19]发现放牧显著降低了短花针茅地上生物量,可能是导致荒漠草原退化的原因之一。曲艳等[20]的研究也表明与围栏禁牧相比,自由放牧显著降低了短花针茅的地上生物量,其原因可能是放牧家畜的践踏、采食和排泄行为对植物群落和土壤的影响,随着载畜率增加,草地负担随之加重,对短花针茅的采食增加,导致生物量下降、营养物质减少、种子库劣变,同时土壤环境的恶化也影响了短花针茅的生长发育。
热值反映绿色植物通过光合作用积累有机物的能力,将太阳能转化成化学能储存在植物体内,并固定CO2的无机碳到植物体中。本研究中,载畜量显著影响短花针茅叶片热值,且随载畜率的增加热值显著降低(P< 0.05),这与卫智军等[21]的研究结果一致。温军等[22]也发现高寒草甸禾本科植物的热值也随退化程度的增加而降低。短花针茅作为荒漠草原建群种,是家畜的主要饲草来源,随着放牧强度增加,家畜践踏和采食作用加剧,短花针茅生物量减少的同时也会导致土壤养分流失,破坏土壤结构[23],使短花针茅再生能力下降,植株矮小化、破碎化,整体热值也随之降低。王玺等[24]的研究发现,短花针茅叶面积随载畜率的增加呈下降趋势,叶面积的减小导致短花针茅光和作用和固定能量的能力的下降,进而导致热值降低。植物热值的差异可能是植物体内碳含量的不同造成的。任何有机体内的有机质骨架都是由碳元素构成的,热值是植物体碳素完全燃烧所释放的能量[2]。因此,植物体内碳素含量越高,植物体完全燃烧的热值就可能越高。王亚婷等[25]发现短花针茅叶片碳含量随载畜率的增加而降低,叶片中碳含量的降低导致光和能力以及叶片干物质含量的下降,导致热值降低,本研究的结果与之一致。
3.2 还原糖含量对载畜率的响应
还原糖含量对维持植物生长发育具有重要作用,是植物生长的灵敏指示剂,可以通过氧化还原反应直接给植物供能[3]。与围封禁牧相比,重度放牧条件下短花针茅叶片还原糖含量显著降低(P<0.05),朱桂林等[26]也发现与禁牧相比,自由放牧区内短花针茅还原糖含量降低。在长期重度放牧条件下,家畜的采食作用导致草地负担加剧,地上植被啃食严重,同时短花针茅会产生大量分蘖,消耗了植物贮藏的以还原糖为基础的营养物质,导致短花针茅叶片还原糖含量显著降低,同时也使得植株破碎化、矮小化。7月份和8月份的短花针茅叶还原糖含量显著低于其他月份的叶还原糖含量(P< 0.05),虽然这2 个月是短花针茅果后营养期,是生物量高峰季,但也因此大量消耗叶片中贮藏的还原糖,而在9月份的生长季结束时,出现的一个积累期为短花针茅的越冬和返青储存能量和营养[27]。
围栏禁牧作为草地生产力的恢复措施被广泛应用于内蒙古草地管理[28],围封导致草地生产力上升,物种多样性显著下降[29],其原因是围封有利于植物生长,草地群落中的优势种对光、热资源的竞争中处于优势地位,而其他物种在竞争中处于不利地位,逐渐消失。从能量功能群的角度分析,围封有利于草地群落中高能量物种的生长发育,而短花针茅是荒漠草原群落中的高能量物种,长期围封导致草地中的能量向短花针茅中移动,热值升高[30]。围封也显著增加了高寒荒漠草原优势植物叶片营养含量,从而促进了植物生长效率[31]。因此,在荒漠草原生态系统中,围封在植物养分响应方面表现出积极的生态保护作用。
