放牧强度对短花针茅植物种群空间分布的影响
2021-07-28李治国吕世杰闫宝龙韩国栋王忠武屈志强
李治国,吕世杰,闫宝龙,韩国栋,王忠武,屈志强,王 静
(内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010011)
短花针茅(Stipa breviflora)荒漠草原具有地带过渡性、生态严酷性和脆弱性特征[1-3],对干扰的响应十分敏感[4-5]。该草地环境中生化可演替成为克氏针茅(S. krylovii)典型草原;草地环境旱生化则演替为小针茅(S. klemenzii)草原;短花针茅荒漠草原生境向暖干条件转变,则演替为短花针茅 + 无芒隐子草(Cleistogenes songorica) + 碱韭(Allium polyrhizum)植物群落(伴随土壤沙化过程),相反生境的暖湿化,则演替为短花针茅 + 无芒隐子草 + 达乌里胡枝子(Lespedeza daurica)植物群落[6]。短花针茅植物种群作为建群种,其控制着植物群落的结构和功能[2-3],掌握短花针茅变化情况有助于了解荒漠草原植物群落的发展动态。
经长期研究显示,以放牧利用为主的短花针茅荒漠草原,其建群种在植物群落中仍占主导地位和作用[7-14];尽管受放牧强度或放牧制度影响,其空间分布状态发生改变,但结构性因素仍是其空间分布的主控因素[15-16];空间多重分形特征存在,且空间自相关程度较高[17-18]。这些空间分布特征的呈现与建群种短花针茅对放牧的响应特征直接相关。有研究表明,短花针茅属密丛型禾草,放牧导致短花针茅株丛发生破碎化,进而使得单位面积密度增加[7,9,13]。以密度指标为基础的短花针茅空间分布研究,缺少放牧强度导致株丛破碎化的考虑,基于高度、盖度和密度综合指标的全面分析,能够整体揭示短花针茅在不同放牧强度下的空间分布状态以及变化规律。
综上,本研究以荒漠草原短花针茅为对象,采用机械取样法测定其高度、盖度和密度,并计算重要值指数,结合变异分析和多重分形分析,明确以下几个问题:首先,短花针茅植物种群空间分布在不同放牧强度下是否存在多重分形特征;其次,如果多重分形特征存在,短花针茅植物种群空间分布的复杂程度或变化程度如何;最后,导致不同放牧强度下短花针茅空间分布发生转变的原因是什么。相关问题的解决可以为草地植物群落稳定性以及家畜对建群种影响的相互关系提供数据支持和理论支撑。
1 试验地概况与研究方法
1.1 试验地概况
试验地位于内蒙古四子王旗短花针茅荒漠草原,并于2004 年建立了长期放牧强度试验基地,地处111°88′00″ E、41°78′00″ N,平均海拔1 440 m。草地建群种为短花针茅,优势种为冷蒿(Artemisia frigida)、无芒隐子草,构成了短花针茅 + 冷蒿 + 无芒隐子草荒漠草原类型。气候属于中温带半干旱大陆性季风气候。土壤为淡栗钙土。
1.2 试验设计
试验区面积为51.9 hm2,自西向东划分成相等的12 个样地,即每个样地面积4.33 hm2。采用随机区组试验设计,设3 个区组,每一区组包含4 个载畜率,分别是不放牧的对照区(CK)、轻度放牧区(LG)、中度放牧区(MG)和重度放牧区(HG)。
LG、MG、HG 放牧区载畜率分别为每公顷每半年0.93、1.82、2.71 个羊单位,对照区载畜率为0。自2004年开始放牧试验,夏、秋季为放牧季节,冬、春季为休牧季节。每天将家畜赶入放牧区,让其自由采食(赶入放牧区之前提供饮水),傍晚赶回畜圈(饮水,补盐),放牧时间自05:30 至17:30,放牧区无饮水设施。
1.3 取样设计
于2016 年8 月在放牧区内选择40 m × 40 m 代表性样地(样地植物群落物种组成相近、地形相近,距围栏门口距离相等),以正西南方为坐标原点建立平面直角坐标系,采用50 m 的测绳两根(精确刻度10 cm),垂直于相对坐标系的横纵坐标,逐一确定取样点的相对坐标(xi,yi),然后将记录下来的相对坐标,按照5 cm 间隔尺度设置取样点并记录相对坐标,图中每一小方格为2.5 m × 2.5 m (图1)。取样点设置样方50 cm × 50 cm (样点位置位于样方中心),分种记录植物种群的高度(cm)、盖度(%)和密度(每0.25 m2株数或丛数)。
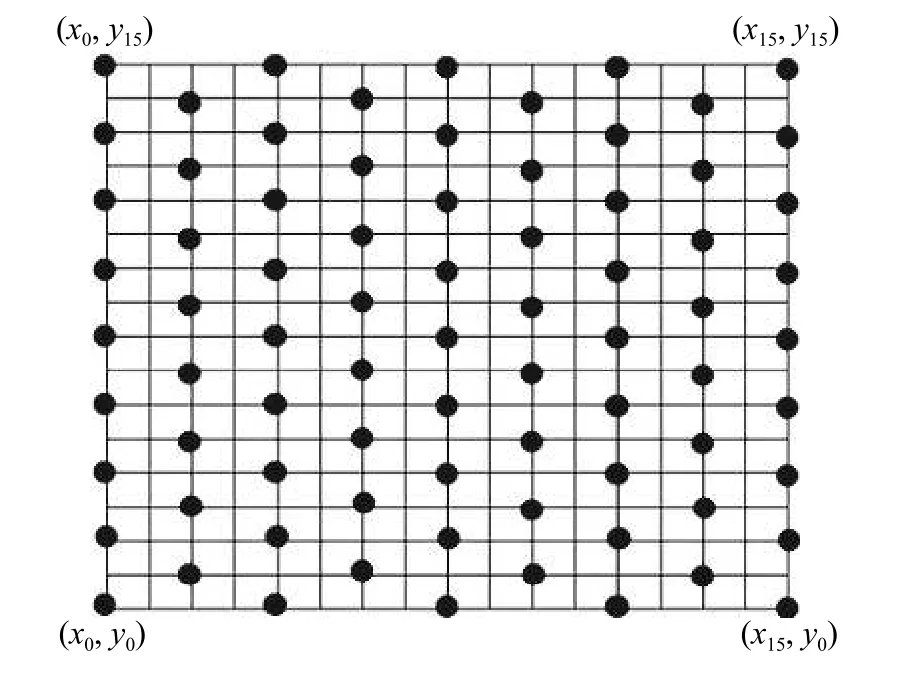
图1 空间取样的样点分布图Figure 1 Spatial sampling point distribution
1.4 数据整理与分析
将得到的坐标点数据进行补全,首先寻找与待补数据点最近的3 个点进行均值插补,如与(x1,y1)距离最近的3 个点分别为(x0,y0)、(x0,y2)和(x2,y1),然后再依次计算相应的待补坐标点数据,分别利用最近的3 个点和最近的4 个点进行均值插补(高度、盖度和密度分别进行插补)。然后计算短花针茅植物种群相对高度(H)、相对盖度(C)和相对密度(D),将三者平均再乘以100%得到短花针茅植物种群在各个样点的相对重要指数(important value index, IVI),每一处理区得到数据个数为289 个的数据集。计算公式如下:
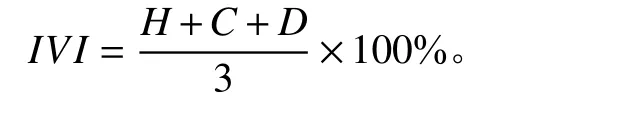
根据多重分形理论及其对数据集的要求(面积取样样点满足22n个),将坐标点含有x17和y17的数据去掉,形成16 × 16 的空间数据点集,然后在Excel 2010 内进行VBA 编程,计算多重分形的相关参数[19-22]。
变异分析:在Excel 2010 中绘制箱式图(也称箱线图),其中线的左右两端分别代表最小值和最大值,即数据的变化范围,箱体由下四分位数和上四分位数构成,表征的是位于上下四分位数之间的数据集中情况,箱体中间的分界线是中位数。


式中:d代表求导,q代表统计阶距,Mi代表取样尺度下第i个样点短花针茅重要指数占比,n为取样样点数,τ(q)为Legendre 变换曲线。
2 结果与分析
2.1 短花针茅在不同放牧强度下的变异分析
短花针茅植物种群在CK 和LG 处理区内,其重要指数最小值均为0 (箱线左端);随着放牧强度的增加,短花针茅重要指数最大值呈逐渐增加的变化趋势,分别为67.02%、75.50%、87.80%和100% (箱线右端) (图2)。箱线最左、最右端的距离,代表重要指数的变动范围,随放牧强度增加,其变动范围没有表现出一定的规律性。箱体反映的是数据集中情况,除CK 处理区外,随着放牧强度的增加,短花针茅重要指数的集中程度随之增加。相对于CK 处理区,LG 处理区的箱体左端较小,而右端较大,其中位数(箱体中线)靠左且接近,说明CK 和LG 处理区的平均重要指数比较接近,经计算其重要指数分别为32.71%和32.55%。由此可见,伴随放牧强度的增加,短花针茅在群落中的重要指数有增大的变化趋势。
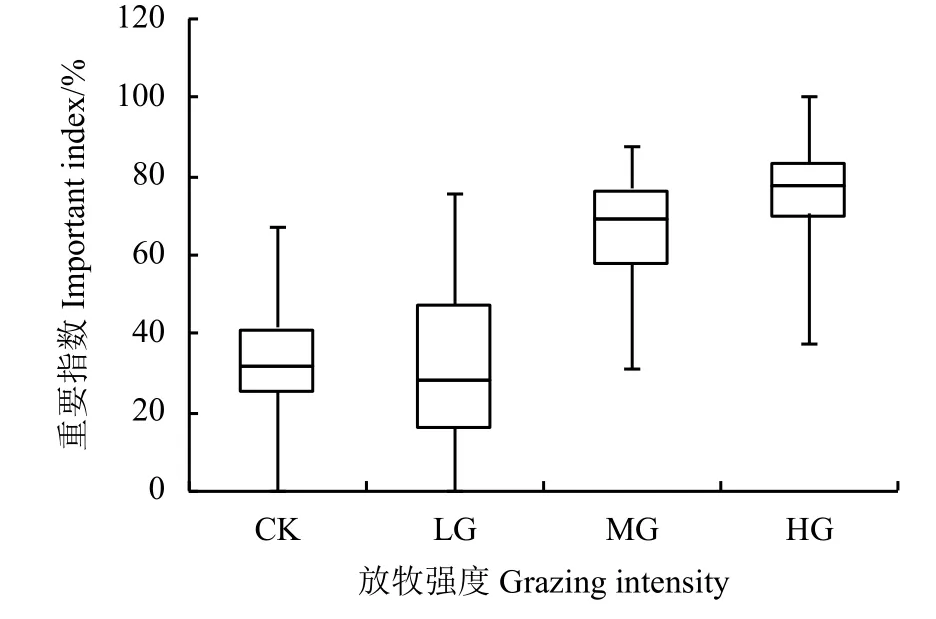
图2 不同放牧强度下短花针茅的重要指数Figure 2 Important indexes of Stipa breviflora under different grazing intensities
2.2 短花针茅植物种群在不同放牧强度下的多重分形判断
在不同放牧强度下,分别计算短花针茅取样尺度及其重要指数密度配分函数的双对数曲线关系,即ε 和χq(ε)的 log(ε)−log[χq(ε)]关系,结果显示(图3),伴随分隔距离对数的增大,重要指数密度配分函数值趋近于0,且在统计阶距-10 ≤q≤ 10 的范围内,双对数曲线均趋于直线且斜率由负值转为正值,这说明短花针茅植物种群在不同放牧强度下存在空间多重分形特征,可以进行多重分形分析。
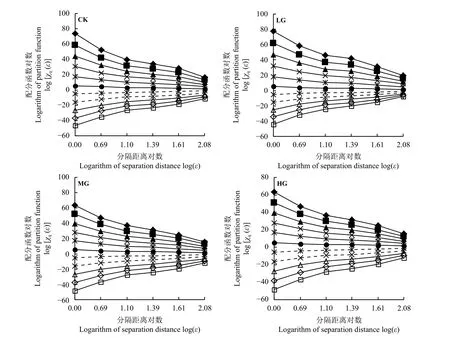
图3 短花针茅重要指数在不同放牧强度下的log(ε)−log[χq(ε)]关系Figure 3 log(ε)−log[χq(ε)] relationship of important indexes of Stipa breviflora under different grazing intensities
2.3 短花针茅植物种群的广义维数谱分析
短花针茅植物种群广义维数谱分析结果(图4)显示,在统计阶距为0 时,CK、LG、MG 和HG 处理区的广义维数谱具有交点,此点代表的是短花针茅植物种群重要指数的计盒分形维数,表征了短花针茅植物种群重要指数空间分布的复杂性和不均匀性,说明在CK、LG、MG 和HG 处理区短花针茅植物种群重要指数空间分布的复杂性和不均匀性比较接近;其计盒维数接近2 (表1),所以短花针茅植物种群空间分布主要由结构性因素控制,且空间自相关程度较高,空间分布比较均匀。
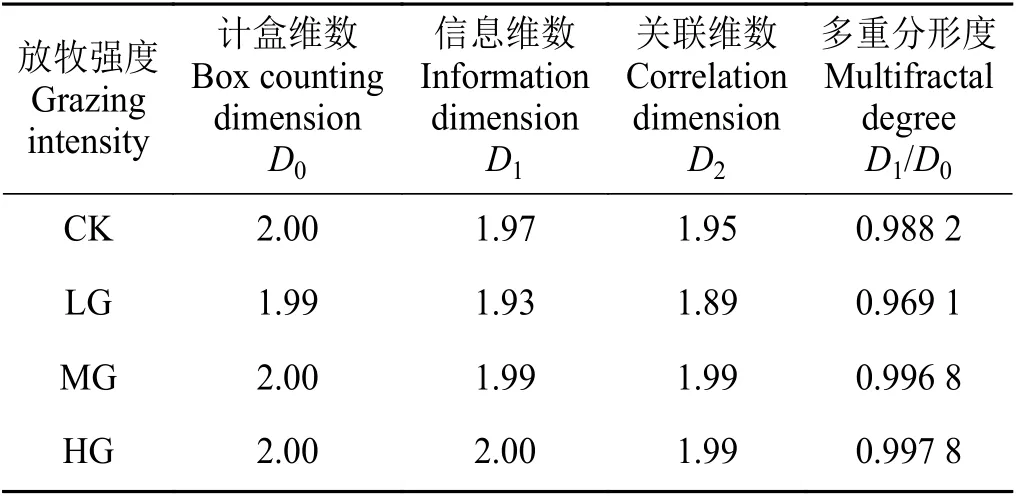
表1 短花针茅在不同放牧强度下的分形维数和多重分形度Table 1 Fractal dimension and multifractal degree of Stipa breviflora under different grazing intensities
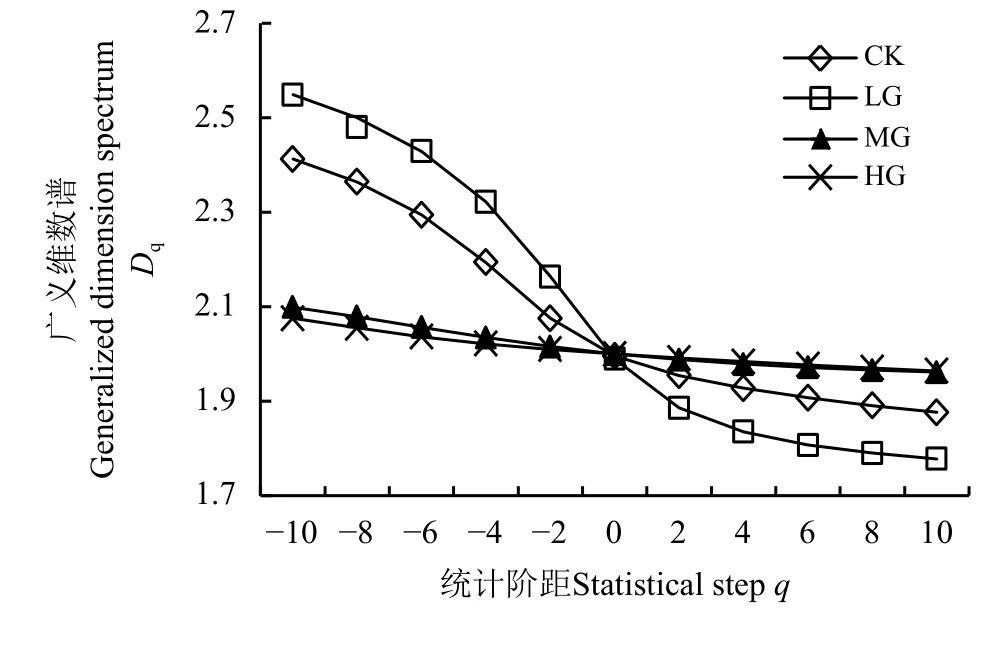
图4 短花针茅植物种群重要指数在不同放牧强度下的广义维数谱曲线Figure 4 Generalized dimensional spectrum curve of the important index of Stipa breviflora population under different grazing intensities
在统计阶距-10 ≤q≤ 10 内,CK 和LG 处理区的短花针茅植物种群广义维数谱变化比较强烈,且变动范围在CK 处理区小于LG 处理区(图4),说明轻度放牧下短花针茅植物种群多重分形特征更为明显,而CK 处理区的短花针茅植物种群多重分形特征较弱。因此,相对来讲,LG 处理区具有更为复杂的空间分布形式。在MG 和HG 处理区短花针茅植物种群重要指数空间分布的复杂性和不均匀性十分接近,尽管统计阶距不同,但二者的广义维数谱比较相似,当统计阶距q< 0 时二者广义维数均小于CK 处理区。综合来看,在CK 和LG 处理区,短花针茅植物种群在群落中的重要性空间变化复杂;反之,在MG 和HG 处理区,短花针茅植物种群在群落中的重要性空间变化简单。
在广义维数谱中,当统计阶距q分别为0、1 和2 时,对应的分形维数分别为计盒维数、信息维数和关联维数。尽管放牧强度不同,但是3 个维数在放牧强度间的大小差异规律比较一致,即在LG 处理区小于CK、MG 和HG 处理区(表1),表明轻度放牧下短花针茅植物种群空间分布的复杂性和不均匀性较高,这主要是因为存在相对随机因素的影响,这一随机因素就是放牧家畜在LG 处理区的高度选择性牧食行为。
尽管不同放牧强度下短花针茅植物种群重要指数的各分形维数存在差异,但是D1/D0值接近于1(表1)。说明尽管放牧强度不同,但是短花针茅植物种群重要指数在群落中均以高值出现,即作为建群种的短花针茅在群落中的主导和控制作用没有发生改变。
2.4 短花针茅植物种群在不同放牧强度下的多重分形奇异谱
由奇异性指数α(q)和多重分形谱f[α(q)]曲线特征(图5)可知,无论放牧强度如何,短花针茅重要指数的多重分形奇异谱均呈“左钩状”,表明短花针茅重要指数在群落中均比较高,其与计盒维数及广义分形谱表征结果一致。尽管短花针茅植物种群在群落中的地位和作用比较重要,且起到主导和控制作用,但是这种作用的大小单从计盒维数等分形维数角度难以细致的刻画,有必要进行进一步的数量分析。
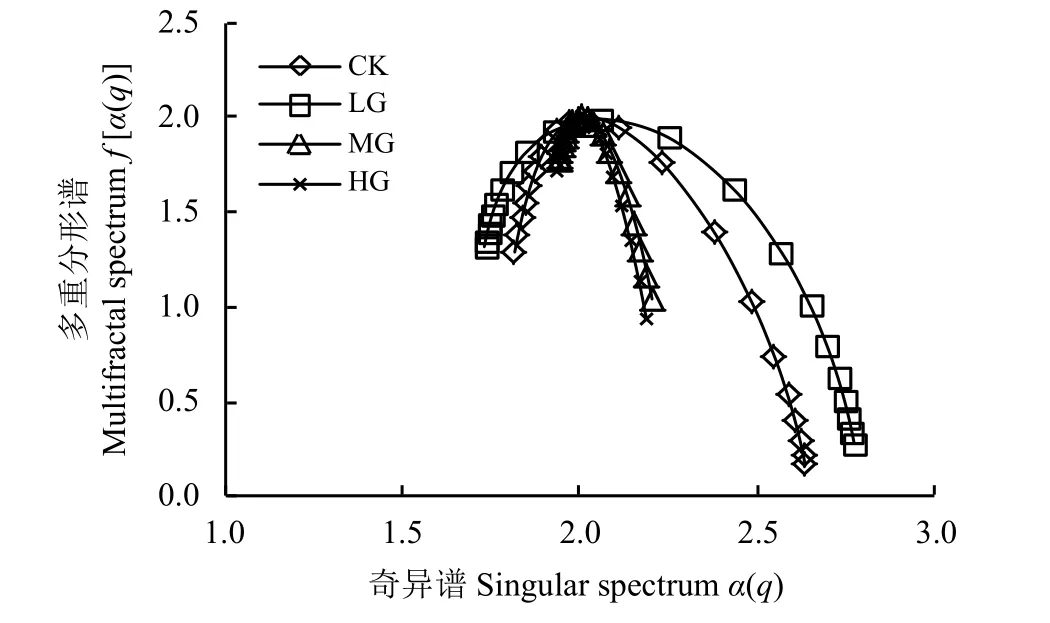
图5 短花针茅重要指数在不同放牧强度下的多重分形奇异谱曲线Figure 5 Multifractal singular spectrum curves of important indexes of Stipa breviflora under different grazing intensities
奇异性指数极差Δα反映的是多重分形谱的宽度,CK、LG、MG 和HG 处理短花针茅重要指数的多重分形谱宽度分别为0.819、1.043、0.263 和0.251,说明轻度放牧下短花针茅植物种群重要指数的空间分布不均匀且复杂程度最高,其次是CK 处理,而MG 和HG 处理短花针茅重要指数的空间分布比较均匀,二者相差不大,但均小于CK 处理(表2)。
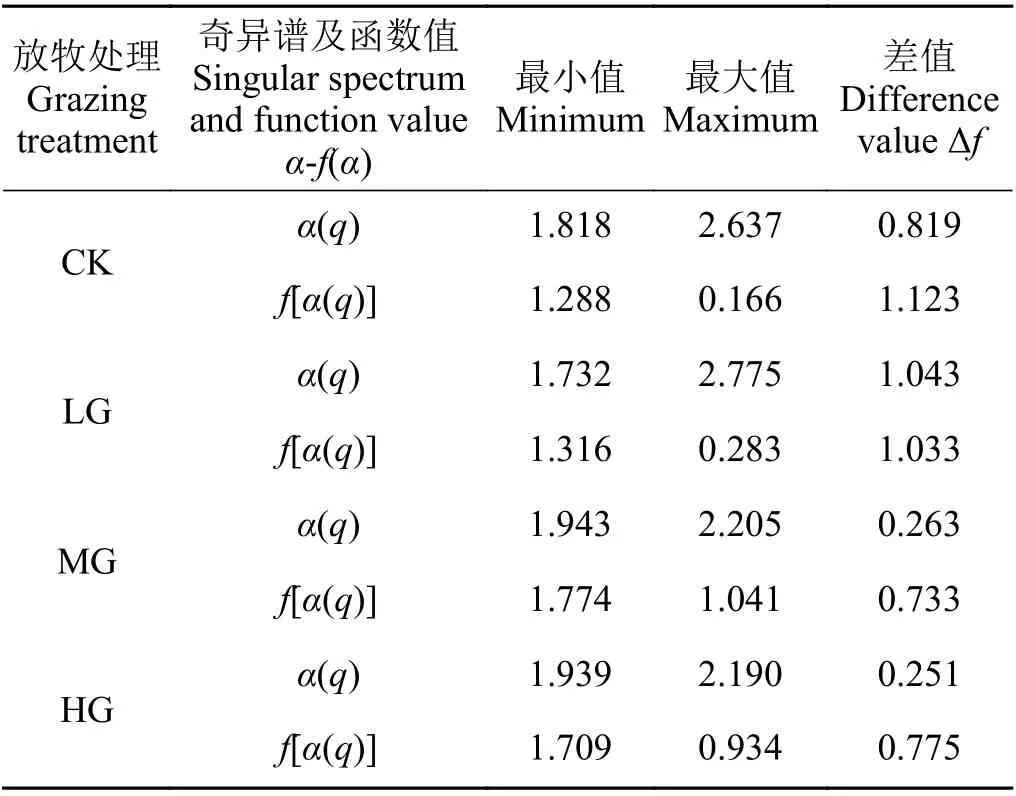
表2 短花针茅植物种群重要指数在不同放牧强度下的多重奇异谱参数Table 2 Multiple singular spectrum parameters of important index of Stipa breviflora population under different grazing intensities
多重分形谱函数值的极差Δf反映的是短花针茅重要指数的概率子集分布情况,若Δf大于0,表征短花针茅重要指数空间分布以大概率子集占主导地位,相反,则表征短花针茅重要指数空间分布以小概率子集占主导地位;结果显示(表2),Δf均大于0,说明大概率子集是短花针茅重要指数空间分布的主要特征,即短花针茅植物种群在群落中占主导地位和作用。
3 讨论
3.1 短花针茅植物种群的多重分形特征
关于短花针茅植物种群的多重分形特征,卫智军等[17]和吕世杰等[18]均进行过相关研究,但是二者得出的结论是短花针茅的空间分布主要以低密度形式分布,这可能是因为短花针茅属于丛生性禾草,其株丛基径变化范围较大,以密度进行空间分形分析,尽管也得到了结构性因素占主导地位和空间自相关程度较高的结论,但却得到了低密度分布属于短花针茅的普遍分布形式这一结论。本研究采用重要值指数,不仅反映了短花针茅的空间占有能力(高度和盖度),也反映了其在单位面积上的有效繁殖能力(密度)。但也有研究者认为,放牧导致短花针茅植物种群株丛发生破碎化(且越大的株丛越容易发生破碎化),从而使得到的密度值偏高,不能够真实反映其繁殖能力[7]。本研究认为,短花针茅本身是多年生丛生禾草,其繁殖包含有性繁殖和无性繁殖,其株丛破碎化是对放牧的积极响应,其有可能在后续发展成相对独立的植株,作为密度统计会表现较高,但也反映其适应策略和未来株丛密度的变化趋势。所以采用重要指数表征短花针茅在群落中的地位和作用比较合理,也在一定程度上反映其受放牧影响的适应和繁殖策略。因此,采用重要指数进行空间分布的探讨,不仅依然证明短花针茅植物种群空间分布存在多重分形特征,且更能够反映短花针茅植物种群在群落中的地位和作用。
在统计阶距q分别为0、1 和2 时,广义维数谱取值分别为计盒维数(容量维数,D0)、信息维数(D1)和关联维数(D2)[23],分别表征植物种群的空间占有能力、空间分布格局强度以及个体空间关联的尺度变化[24-25]。因此,从空间占有能力来看,轻度放牧可使短花针茅植物种群的空间占有能力小幅下降,但中度放牧和重度放牧不会改变短花针茅植物种群的空间占有能力。从格局强度来看,轻度放牧使得短花针茅植物种群空间分布聚集性增加(D1= 1.93),伴随放牧强度的增大,短花针茅植物种群空间分布由聚集性向均匀性转变(D1值逐渐增大,轻度放牧除外)。从关联维数来看,短花针茅植物种群空间分布的自相关强度在轻度放牧处理区最低,表征其个体间的竞争压力最小,伴随放牧强度的增加,短花针茅植物种群个体空间的竞争强度随之增加(轻度放牧除外)。因D1/D0越接近于1,表征短花针茅植物种群空间分布越集中于密集区域[18],而本研究结果显示各放牧强度下短花针茅植物种群的D1/D0值均大于0.95,接近于1,所以尽管放牧强度不同,短花针茅植物种群空间分布整体上均集中于密集区域。这与图5 和表2 的研究结果一致。
3.2 短花针茅植物种群空间分形与其在群落中地位和作用的关系
由于短花针茅是试验区荒漠草原的建群种,其在群落中的地位和作用及其对群落结构和功能的影响得到许多研究者的一致认可[8-10]。本研究结果显示,多重分形可以明确短花针茅植物种群空间分布的不均匀性,能够阐释究竟是大概率子集还是小概率子集在空间分布上占主导地位,但是难以反映不同放牧强度之间短花针茅植物种群在群落中的作用大小及其与复杂度和多重分形谱的关系。经过比对发现(除CK 外),随着放牧强度的增加,短花针茅在群落中的地位和作用增强,同时其空间分布复杂程度降低,但与其空间分布的高概率子集并没有明显的相关关系。由于无论放牧强度如何,短花针茅空间分布均以高概率子集分布占主导地位,同时短花针茅是建群种,对群落结构和功能起主导作用,且伴随放牧强度的增加,其在群落中的地位和作用进一步加强。因此,放牧导致建群种短花针茅植物种群在群落中的主导作用增强,其在高强度放牧下维持群落稳定及决定群落结构和功能方面具有决定性作用,这一点可以从前人研究的生物量方面得到佐证[2,26]。
由于计盒维数、信息维数和关联维数的生态学意义不同[18,24],在不同放牧强度下,尽管短花针茅植物种群在植物群落中的主导作用依然存在,但是均有自己的变化特点。首先,轻度放牧导致短花针茅植物种群计盒维数降低,表征其空间占有能力下降,也意味着其生态位宽度在下降;此时短花针茅空间分布的聚集性增加,个体间的竞争强度减弱。其次,伴随放牧强度的增大(轻度放牧除外),短花针茅植物种群的生态位宽度几乎没有变化,空间分布由聚集性向均匀性转变,个体间的竞争强度随之增大。这一变化规律显示重度放牧区需要调整生态适应策略,以维持其种群延续及其在植物群落中的主导地位;通过对短花针茅植物种群的数量特征分析,株丛破碎化与其重牧条件下的适应策略直接相关[27-28]。
4 结论
在不同放牧强度下,短花针茅植物种群空间分布存在多重分形特征;伴随放牧强度的增大(轻度放牧除外),短花针茅植物种群空间分布的多重分形复杂性降低:生态位宽度几乎没有变化,空间分布由聚集状态转为均匀状态,个体间的竞争强度增大;轻度放牧下短花针茅植物种群空间分布多重分形特征最为复杂:生态位宽度降低,空间分布聚集特征最明显,个体间的竞争强度最小。无论放牧强度如何变化,短花针茅植物种群在植物群落中的主导地位没有改变,空间分布的整体特征是以大概率子集空间分布为主。
