不同浓度镉对烟草幼苗生长发育及生长素相关基因表达的影响
2021-03-12罗勇焦桂珍刘胜波魏跃伟邵惠芳贾宏昉
罗勇, 焦桂珍, 刘胜波, 魏跃伟, 邵惠芳, 贾宏昉*
(1.河南农业大学烟草学院, 郑州 450002; 2.史丹利化肥宁陵有限公司, 河南 商丘 476000)
重金属镉(Cd)的生物毒性很强,是植物生长发育的非必需元素[1]。近年来,工业和农业生产不同程度地将镉排放到土壤环境中,成为危害最大、分布最广的农业污染物之一[2]。2014年,我国土壤镉的点位超标率为7.0% ,位居8种无机污染物之首[3]。在土壤-植物系统中,镉较其他重金属更易迁移,可通过植物根系的吸收在植株体内积累[4],造成生态环境污染和农产品品质下降[5],并通过食物链危害人体健康[6-7]。镉是人体最易积累的毒性物质,半衰期长达6.2~18年[8]。镉积累到一定程度时,就会对植物产生毒害[9-11],包括影响细胞分裂,降低生长速率[12],破坏光合作用[13-14],影响植物酶活性与膜完整性,促进脂质过氧化[15]等。通常植株会出现发育缓慢、矮小、叶片失绿等症状,进而作物产量和质量降低。
烟草作为我国乃至世界上一种非常重要的经济作物,有着较强的镉富集能力。镉积累过量降低烟草的品质和产量,并在卷烟抽吸过程中通过烟气进入人体,影响人体健康。目前镉对烟草生长发育的作用机制研究已有一定的进展,但大多数研究集中在抗氧化性方面,镉对烟草幼苗生长素及相关基因表达的影响报道较少。生长素是调节植物发育、参与胁迫反应中根系构型变化的一种关键激素。
因此,本研究以能标记生长素的DR5∷GUS云烟87转基因烟草为试验材料,探究不同浓度镉处理对烟草生物学特性、MDA含量、生长素水平及NtYUCCA、NtPINs基因家族表达的影响,揭示不同浓度镉对烟草生长发育的作用机理,以期为控制烟草重金属镉含量,提高烟叶安全性提供科学依据。
1 材料与方法
1.1 试验材料
试验所使用的烟草材料为DR5∷GUS云烟87转基因纯合株系。转基因烟草材料的获得以及pDR5∷GUS的构建参照Jia等[16]方法。
1.2 试验设计
将饱满均匀的烟草种子消毒后播种到培养皿中,将点好种子的培养皿置于28 ℃暗室中催芽3 d。将发芽后的烟草幼苗置于人工气候培养箱培养2周,第1周使用1/4霍格兰氏(Hoagland’s)营养液培养,第2周使用1/2营养液培养,每3 d更换一次营养液。选择长势一致的烟苗移至装有石英砂的PVC塑料碗中,1/2 营养液缓苗3 d后,使用全营养液进行镉处理21 d后,取样检测。人工培养箱培养条件为光强280 μmol·m-2·s-1,28 ℃/24 ℃(16 h昼/8 h夜),相对湿度72%。以氯化镉水合物(CdCl2·2.5H2O)作为镉源。将CdCl2·2.5H2O用蒸馏水溶解,加入到Hoagland’s营养液中。镉浓度分别设定为0(Cd0)、5(Cd5)、10 (Cd10)、20 (Cd20)、40 μmol·L-1(Cd40)。每个处理3次重复。
1.3 试验方法
1.3.1烟草表型的记录和侧根数的测定 使用尼康D7000相机拍照,记录不同镉浓度下的烟草表型,人工测定二级根的数量。
1.3.2生物量测定 对烟草植株进行地上部和地下部采样,用JA2003天平(上海仪分科学仪器有限公司)称重,记录样品鲜重。
1.3.3GUS染色和NBT染色 将新鲜洗净的烟苗整株浸没于β-葡萄糖苷酸酶(GUS)染液中,37 ℃恒温培养箱中避光染色24 h后,用95%乙醇浸泡脱色。将新鲜的清洗干净的烟苗浸没于氮蓝四唑(NBT)染液中, 30 ℃恒温培养箱中避光染色5 h,使用95%乙醇脱色。用德国蔡司Stemi 508体式显微镜观察,并拍照记录。
1.3.4叶绿素含量和MDA含量的测定 叶绿素含量测定参照吴德慧[17]方法,丙二醛(malondialdehyde,MDA)含量测定参照Feng等[18]方法。Cd含量测定参照张晓等[19]方法,使用电感耦合等离子体质谱仪(ICP-MS 7500A,美国Agilent)测定镉含量。将烟苗的地上部和地下部取样,于-70 ℃冷冻保存。参照Jia等[20]方法测定烟株中生长素(IAA)含量。IAA标准品购自Sigma-Aldrich公司。
对新鲜烟苗整株取样,根据GenBank发布的生长素相关基因的序列设计扩增引物,以NtL25为内参基因,由北京擎科新业生物技术有限公司合成,引物信息见表1。参照Jia等[20]方法进行qRT-PCR分析。每个样品3次重复,实时定量的结果采用2-ΔΔCt方法[21]进行分析。
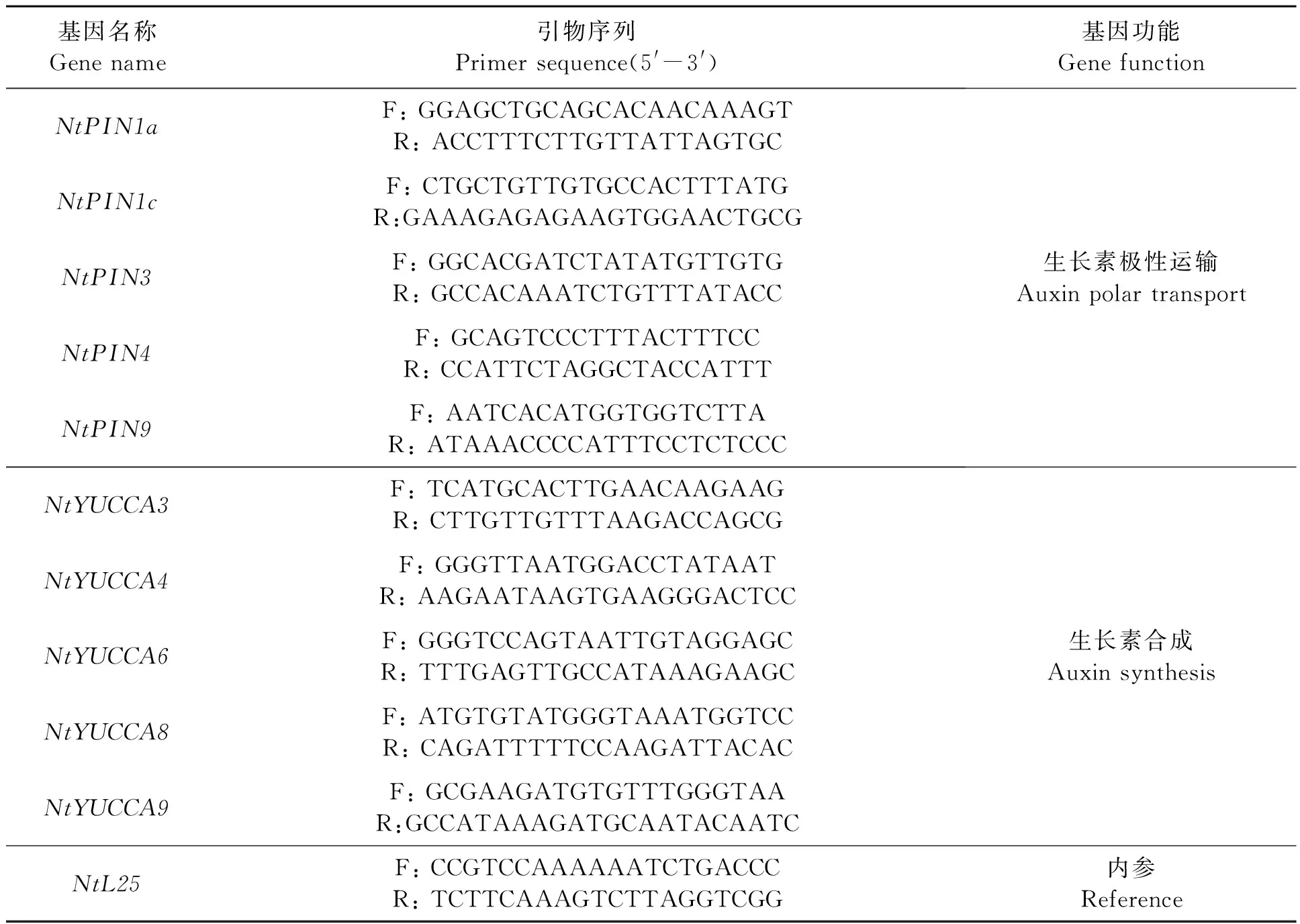
表1 引物信息Table 1 Primer sequences
1.4 数据处理
采用Sigmplot 2018和DPS进行数据分析和整理。
2 结果分析
2.1 不同浓度镉处理对烟苗表型和生物量的影响
5个不同浓度镉处理下的烟苗表型见图1。与对照Cd0相比,随着供镉水平的不断提高,烟草幼苗逐渐矮化,主根变短,侧根数减少,叶子发黄,叶片短小。烟苗鲜重和侧根数结果(图1)显示, Cd5、Cd10、Cd20、Cd40处理的生物量(鲜重)分别较Cd0显著减少11.1%、38.0%、45.8%、73.1%,而侧根数分别较Cd0显著减少26.7%、40%、53.3%、73.3%。结果表明,镉浓度的不断增加对烟苗产生了毒害作用,抑制了其正常的生长发育。
2.2 不同浓度镉处理对烟苗镉含量的影响
烟苗中镉含量的测定结果(图2)显示,随着供镉水平不断提高,烟苗中的镉含量呈上升趋势。与Cd5相比,Cd10、Cd20、Cd40处理中的镉含量分别显著增加84.8%、353.4%、681.7%。烟草对镉的富集作用导致烟苗中镉含量增加,40 μmol·L-1处理下镉积累最多,表明该处理下镉对烟苗的毒害作用更强,这与表型及生物量结果一致。
2.3 不同浓度镉处理对烟苗叶绿素含量的影响
叶绿素是植物进行光合作用的重要物质,植物在逆境条件下叶绿体结构遭到破坏, 从而导致叶绿素含量下降,光合作用相关的酶失活或变性。由图3可以看出,叶绿素含量随供镉水平的增加而降低,且均达到显著差异水平。Cd20处理的叶绿素含量下降至Cd0的39.9%, Cd40处理下降为Cd0的24.5%,叶绿素大量降解。
2.4 不同浓度镉处理对烟苗抗氧化能力的影响
2.5 不同浓度镉处理对烟苗生长素分布和含量的影响
为了探究镉处理是否对烟苗生长素合成或运输产生影响,利用DR5∷GUS云烟87能够标记生长素分布的特性[22],对根尖部位进行染色,GUS染色的颜色深浅可以反应植物组织中生长素分布情况。染色结果(图5)显示,随着镉胁迫程度的加深,蓝色逐渐变浅,在根尖聚集的生长素越来越少,维管束中的生长素也逐渐减少。不同处理的地上部和地下部生长素含量均显著下降,Cd40处理的烟苗地上部和地下部生长素含量仅为Cd0的40.4%和15.5%。镉处理可能抑制了烟苗中生长素的合成,以及生长素从形态学上端运输到形态学下端的极性运输。
2.6 不同浓度镉处理对烟苗生长素相关基因表达的影响
烟苗中生长素合成限速酶NtYUCCA基因家族和生长素极性运输蛋白基因家族NtPINs的基因表达结果(图6)显示,除NtYUCCA3、NtPIN3的表达水平变化不显著外,生长素相关基因的相对表达量均随着供镉水平的上升而降低。与Cd0相比,Cd5、Cd10、Cd20、Cd40处理的生长素合成基因NtYUCCA4和NtYUCCA8的相对表达量均显著降低,其中Cd40处理分别显著下降60.1%和66.4%;NtYUCCA6和NtYUCCA9的相对表达量在Cd5处理有所下降,但差异不显著,Cd40处理下分别显著降低54.8%和70.2%;生长素极性运输蛋白基因NtPIN1a、NtPIN1c、NtPIN4的相对表达量均显著降低,Cd40处理分别下降70.8%、74.5%、58.5%;NtPIN9的相对表达量在Cd5和Cd10处理中分别下降1.5%和3.4%,但差异不显著,在Cd20、Cd40处理中分别显著降低33.1%和54.2%。结果表明,在镉胁迫下生长素合成和极性运输相关基因表达均受到抑制,且抑制作用随着镉浓度的增加而增强。
3 讨论
本研究发现,随着供镉水平的增加,镉会严重影响烟草幼苗的生长发育。在40 μmol·L-1的镉浓度下,烟草幼苗生物量比对照减少73.1%,根系发育不正常,侧根数下降73.3%。这与前人对烟草[23-24]及冬小麦[25]的研究结果一致。而贾月慧等[26]研究指出,50、100 mg·L-1的镉对生菜生物量干质量和鲜质量均有显著促进作用。这可能是因为不同作物对镉的吸收与品种、pH、离子环境等有关,因而其有效浓度存在较大差异,但总体趋势是高浓度镉对作物生长表现出抑制作用。烟草属于易富集镉的作物[27],本研究发现在40 μmol·L-1的镉浓度下,烟苗镉含量增加681.7%。

