长白山不同林型土壤跳虫多样性特征及其影响因子1)
2021-03-09吴新毅冯立超李燕徐炜超孟庆繁
吴新毅 冯立超 李燕 徐炜超 孟庆繁
(北华大学,吉林省·吉林市,132013) (吉林农业科技学院) (北华大学)
在陆地生态系统中,跳虫群落组成繁杂、数量庞大,物种多样性高,是土壤内重要的动物类群之一[1]。土壤跳虫在土壤养分循环中起着重要的作用,土壤内土壤动物活动直接改变土壤内有机物质矿化过程,影响植物群落对土壤养分的利用[2-3],而植物群落改变亦能影响土壤理化性质变化[4-5],从而改变植物群落自身生产力、物种分布范围、种群组成及演替[6]。在跳虫与植被、生物因子及环境因子互作影响的研究中,土壤跳虫通过选择性地抑制特定植物从而间接地促进其他植物种群发展,进而影响植物群落多样性组成[7-9]。通过食物网的调节作用改变土壤内其他生物的群落,间接影响土壤内物质循环,对植物生长产生促进或抑制作用[10-11],并进一步改变地上植物群落结构。由地上植被类型、植物凋落物组成所主导的土壤有机物(有机碳、氮等),土壤微生物组成直接决定以土壤微生物、动物残体及植物凋落物为食的土壤跳虫组成结构[12-14]。土壤理化性质变化如pH、土壤含水量等可显著改变跳虫群落结构[15-16],林分类型变化对土壤跳虫群落的直接影响相对较小[17]。
我国有关土壤跳虫生态学的研究起步于20世纪80年代末[18],虽然起步相对较晚,但发展迅速。天然林生态系统系内,土壤跳虫群落结构与多样性受生境和季节变化影响显著,生境的植被类型与土壤理化性质共同决定土壤跳虫分布[19],而在农田生态系统中,土壤跳虫的物种数在各生境差异较小,土壤跳虫个体密度和物种数存在季节差异,这种差异因生境而异[20]。长白山不同山地森林类型下土壤跳虫的分布存在差异,在退化程度不同的森林中呈现不同群落的组成[21]。为弄清长白山植被如何通过影响土壤理化性质进而影响跳虫群落结构,本研究以长白山自然保护区不同林型下土壤跳虫群落为研究对象,从植物群落组成及土壤理化性质等方面研究土壤跳虫的分布规律,探讨林分类型、土壤理化性质对土壤跳虫物种组成及多样性格局的影响,揭示长白山土壤跳虫群落的分布特征,从而了解土壤跳虫对环境的适应机制以及土壤跳虫的生态功能,为自然保护区生态资源多样性保护提供理论依据。
1 研究区概况
长白山自然保护区位于吉林省东南部,是我国建立较早的以森林生态系统为主要保护对象的自然综合体。地理坐标为:41°41′49″~42°25′18″N,127°42′55″~128°16′48″E。保护区总面积19.6万 hm2。该区为中国生物多样性热点地区,也是世界上同纬度物种最丰富的地区。长白山山体高大,是东北亚地区海拔最高的山脉,中国境内最高峰白云峰,海拔2 691 m。该区属温带大陆性气候,冬季漫长严寒,夏季短暂炎热。年降水量变幅为800~1 800 mm,年均温度3.6 ℃,年相对湿度为65 %~74 %[22]。长白山植被垂直分布明显,从低海拔到高海拔依次为阔叶红松林带、暗针叶林带、亚高山岳桦林带、高山冻原带。主要组成树种有红松(Pinuskoraiensis)、白桦(Betulaplatyphylla)、岳桦(B.ermanii)、紫椴(Tiliaamurensis)、槺椴(T.mandschurica)、槭树(Acersaccharum)、榆树(Ulmuspumila)、山杨(Populusdavidiana)、鱼鳞云杉(Piceajezoensis)、臭冷杉(Abiesnephrolepis),其中红松为建群种。
2 研究方法
2.1 样地设置
2018年7—8月份,选取长白山自然保护区北坡杨桦林、阔叶红松林和云冷杉林3个林型[22],每个林型内各选择6块代表性林分设置样地,作为重复,样地面积为20 m×30 m。
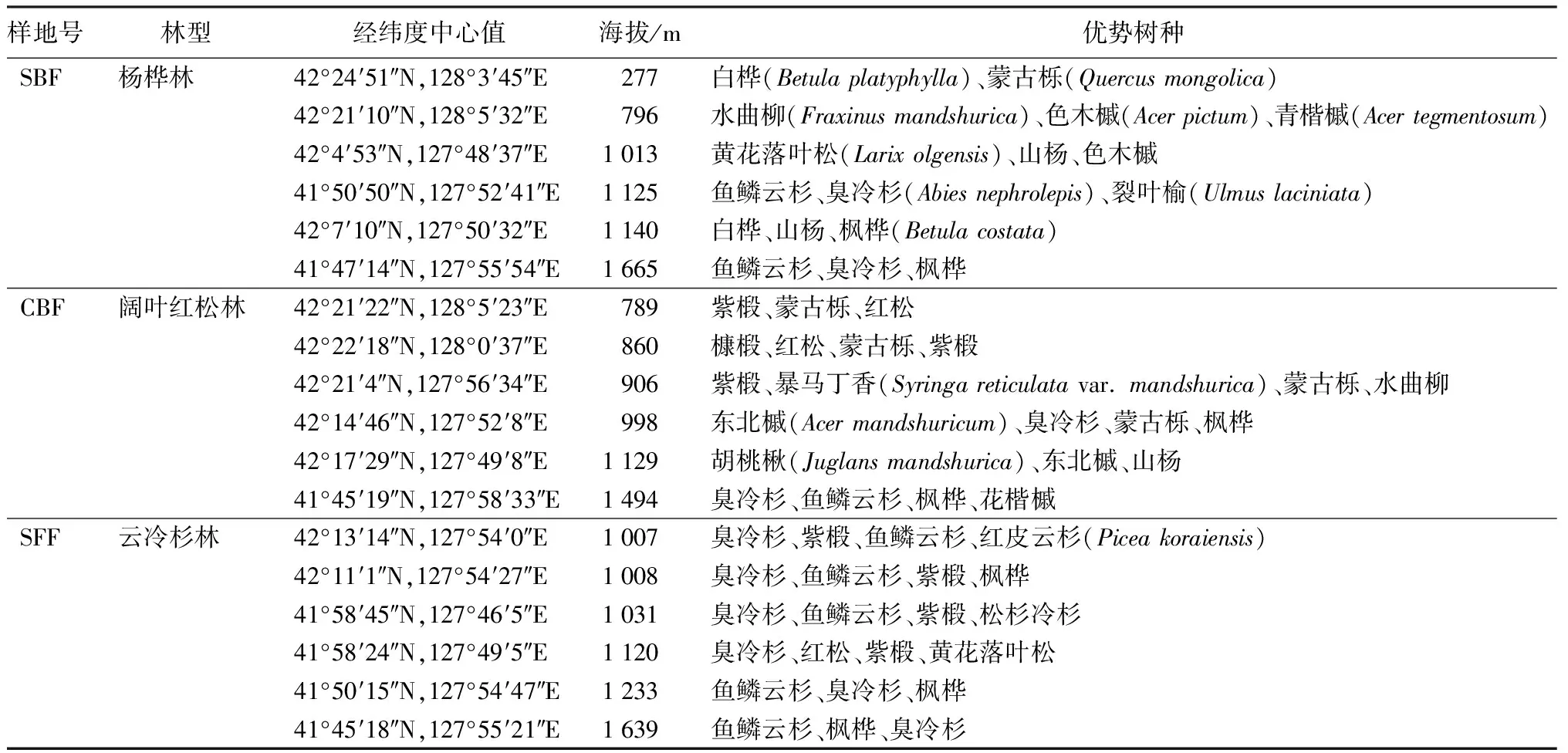
表1 长白山自然保护区不同林型样地概况
2.2 土壤样本采集
分别在样地四角和对角线中心点各设一个1 m×1 m样方,在样方内进行3次采样,将采集点上枯枝落叶层样本装入自封袋内,利用直径为5 cm的土钻在3个采样点采集土样,采集深度15 cm,每个样品分别装入自封袋内,编号,带回实验室分离。
2.3 跳虫分离与鉴定
采用干漏斗(Berlese-Tullgren)分离法分离土壤跳虫。将野外采集的样品放置于装有漏斗的筛子上,漏斗下部接装含有质量分数75 %酒精的装置,用以收集和保存分离后的土壤跳虫,分离时间为7~12 d。将装有跳虫样品的装置倒入另一玻璃瓶内进行水浴加热,待瓶内酒精煮沸,悬浮的跳虫标本下沉后取出,进行跳虫分拣工作。跳虫分拣按照形态学进行划分,记录个体数,保存于装有质量分数75%酒精离心管内[23]。分离后的跳虫标本按照褪色、洗涤和制片等玻片标本制作流程将跳虫样本制成玻片标本并鉴定。
2.4 土壤理化性质分析
将野外采集的土壤样品进行自然风干,手动挑拣出大石块、植物根系,再用研钵将土壤样品磨细,过20目、100目筛,筛选出2份土样,装于标号的封口袋中。过20目筛的土样用于测土壤pH,过100目筛的土样用于测全氮、全磷、有机质、有机碳质量分数。采用V(水)∶m(土)=2.5∶1.0电极法测定pH值。采用烘干称质量法测量土壤含水量。采用电位法测量土壤还原电位值。采用油浴法测定土壤有机质质量分数。采用重铬酸钾氧化—外加热法测定土壤有机碳质量分数。采用凯氏定氮法测量土壤全氮质量分数。采用碱熔-钼锑抗比色法测量土壤全磷质量分数[24]。
2.5 数据分析
采用Shannon-Wiener多样性指数(H′)、Pielou均匀度指数、Berger-Parker优势度指数计算各林型下土壤跳虫群落多样性。采用Jaccard相似性系数计算各不同林型间的相似性[25-26]。
统计方法采用Canoco5.0多变量排序分析软件冗余分析RDA分析方法,对比不同林型下弹尾纲(Collembola)种群多样性;采用SPSS分析软件,运用非参数检验统计分析方法Kruskal-Wallis检验、单因素方差分析方法比较不同林型下弹尾纲种类及数量组成差异性。
3 结果与分析
3.1 不同林型下土壤跳虫群落组成及α多样性
共采集土壤跳虫534头,隶属8科,20属,30种。不同林型下土壤跳虫的物种组成见表2。
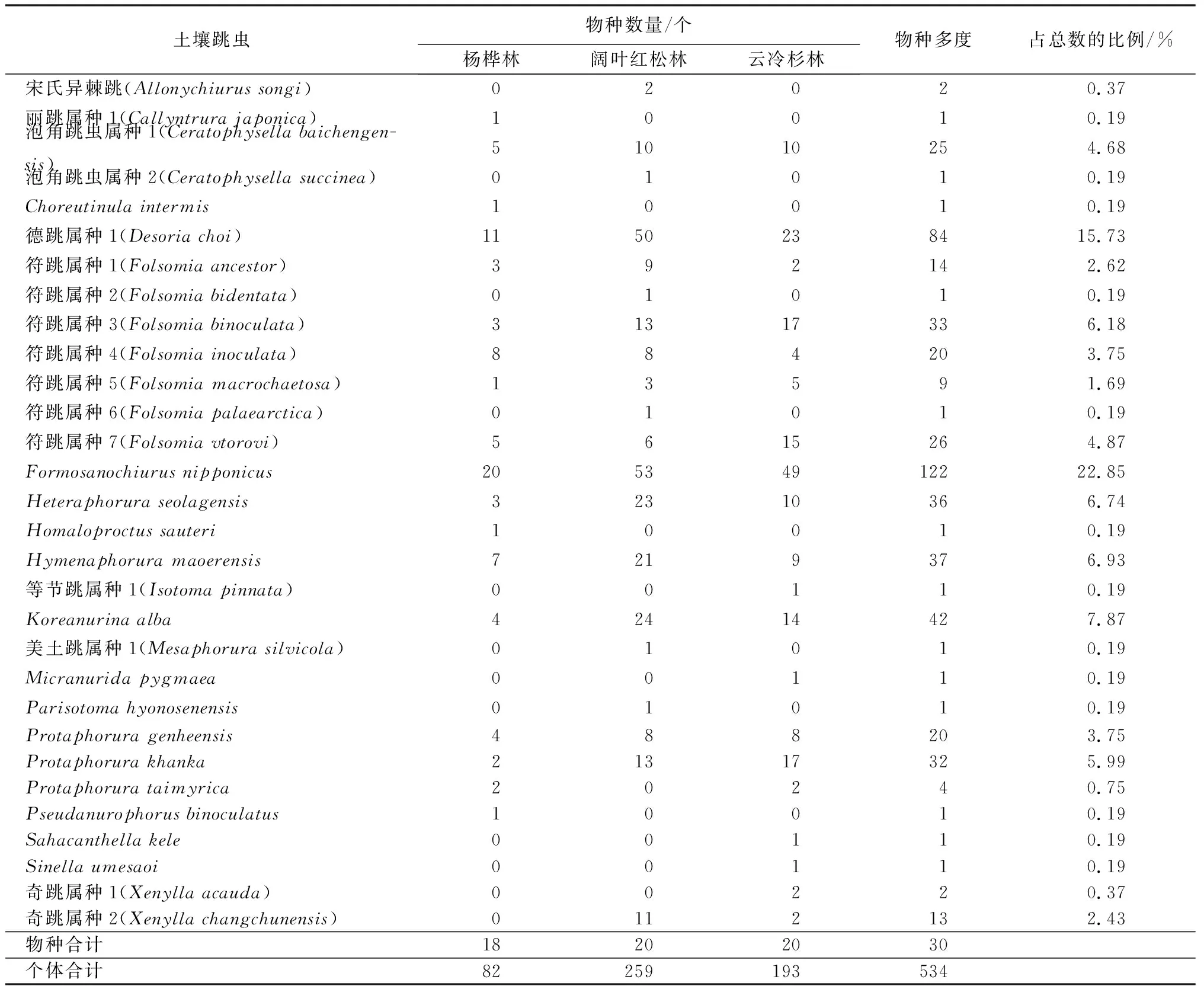
表2 长白山自然保护区不同林型土壤跳虫物种组成
由表2可见,保护区3个林分类型下土壤跳虫优势种为Formosanochiurusnipponicus和德跳属种1,分别占跳虫总个体数的22.85%和15.73%;Koreanurinaalba、Hymenaphoruramaoerensis和Heteraphoruraseolagensis为亚优势种,共占总个体数量的21.54%;等节跳属种1、Parisotomahyonosenensis、符跳属种6、泡角跳虫属种2、Sahacanthellakele、丽跳属种1、Sinellaumesaoi、Choreutinulaintermis、Micranuridapygmaea、Homaloproctussauteri、Pseudanurophorusbinoculatus、美土跳属种1和符跳属种2,共13种稀有种,共占总个体数量的2.43%。不同林型间,跳虫的物种组成结构不同,杨桦林和云冷杉林跳虫优势种为德跳属种1、Formosanochiurusnipponicus,而阔叶红松林跳虫优势种为德跳属种1、Formosanochiurusnipponicus和Koreanurinaalba。
杨桦林下土壤跳虫物种数最少,低于阔叶红松林与云冷杉林,但3种林型间跳虫物种数差异不显著(P=0.251)。在杨桦林下,跳虫个体数最少,低于阔叶红松林与云冷杉林(P<0.05)(表3)。

表3 长白山自然保护区不同林型下土壤跳虫个体与物种数
由表4可知,3个林型间土壤跳虫物种多样性指数(H′)差异不显著(P=0.796)。杨桦林跳虫物种多样性指数最高,针阔混交林跳虫物种多样性指数(H′)最低;3个林型间跳虫均匀度指数(J)(P=0.07)和优势度指数(W)(P=0.218)差异也均不显著,但多重比较结果表明,杨桦林与阔叶红松林的均匀度指数差异显著。

表4 长白山自然保护区不同林型下土壤跳虫α多样性
3.2 不同林型下土壤跳虫β多样性
由图1可见,不同林型间跳虫群落物种组成差异较大,其中,3个林型共有跳虫物种为13种,分别是Formosanochiurusnipponicus、德跳属种1、Koreanurinaalba、Hymenaphoruramaoerensis、Heteraphoruraseolagensis、符跳属种4、Protaphorurakhanka、符跳属种7、泡角跳虫属种1、Folsomiainoculata、Protaphoruragenheensis、符跳属种1、Folsomiamacrochaetosa;杨桦林与云冷杉林共有种1种,为Protaphorurataimyrica;阔叶红松林与云冷杉林共有种1种,为奇跳属种2;杨桦林与阔叶红松林间无共有种;杨桦林独有4种,分别为Callyntrurajaponica、Choreutinulaintermis、Homaloproctussauteri、Pseudanurophorusbinoculatus;阔叶红松林独有6种,分别为宋氏异棘跳、Parisotomahyonosenensis、符跳属种6、泡角跳虫属种2、美土跳属种1、符跳属种2。云冷杉林独有5种,分别为奇跳属种1、等节跳属种1、Sahacanthellakele、Sinellaumesaoi、Micranuridapygmaea。长白山自然保护区内不同林型间,跳虫群落组成具有中等相似性。其中阔叶红松林与云冷杉林的群落相似性最高,为0.58,杨桦林与阔叶红松林的群落组成相似性最低,为0.52。
3.3 林分类型及土壤理化性质对土壤跳虫种类影响
由图2可见,泡角跳虫属种2、Protaphorurataimyrica、Parisotomahyonosenensis、符跳属种3等主要跳虫物种与云冷杉林呈显著正相关关系;德跳属种1、Protaphorurakhanka、Hymenaphoruramaoerensis更倾向于分布在阔叶红松林;符跳属种7、符跳属种5、Heteraphoruraseolagensis与杨桦林呈显著负相关关系;在土壤理化性质中,随着pH下降,土壤内跳虫物种丰富度逐渐升高,呈显著负相关关系;土壤碳氮比、土壤温度对跳虫生存影响具有促进作用,呈显著正相关关系;土壤还原电位越高,符跳属种1、Formosanochiurusnipponicus、Ceratophysellabaichengensis、符跳属种7物种个体数量越少,为显著负相关关系;土壤含水量与符跳属种2呈显著正相关关系,而土壤有机质与符跳属种2呈显著负相关关系。土壤养分(N、P)对大多数跳虫组成无明显影响。

图1 不同林型间跳虫物种分布差异

SBF.杨桦林;CBF.阔叶红松林;SFF.云冷杉林;Tem.温度;SD.土壤结构;WC.含水量;SRP.还原电位;SOM.有机质;SOC.有机碳;C/N.碳氮比;tN.全氮;tP.全磷;sp1.德跳属种1;sp2.等节跳属种1;sp3.符跳属种1; sp4.符跳属种3; sp5.符跳属种7;sp6.符跳属种5;sp7. Parisotoma hyonosenensis;sp8.符跳属种6;sp9.符跳属种4;sp10.Protaphorura genheensis;sp11.Protaphorura khanka;sp12.Hymenaphorura maoerensis;sp13.Formosanochiurus nipponicus;sp14.Protaphorura taimyrica;sp15.Koreanurina alba;sp16.宋氏异棘跳;sp17.奇跳属种2;sp18.泡角跳虫属种2;sp19.泡角跳虫属种1;sp20.Heteraphorura seolagensis;sp21.奇跳属种1;sp22.Sahacanthella kele;sp23.丽跳属种1;sp24.Sinella umesaoi;sp25.Choreutinula intermis;sp26.Micranurida pygmaea;sp27.Homaloproctus sauteri;sp28.Pseudanurophorus binoculatus;sp29.美土跳属种1;sp30.符跳属种2。
4 结论与讨论
不同林型间土壤跳虫群落结构差异较大,土壤理化性质对土壤跳虫群落分布影响显著。植被类型与土壤理化性质共同决定土壤跳虫群落组成结构特征。
4.1 不同林型间土壤跳虫群落组成结构比较
本研究比较分析了长白山自然保护区杨桦林、阔叶红松林和云冷杉林3个典型林分类型间土壤跳虫多样性差异。其中,在杨桦林中,跳虫种类和个体数量最低,多样性指数和均匀度指数最高;在阔叶红松林中,跳虫种类和个体数量最多,多样性指数与均匀度指数最低。在3个林型间,跳虫群落组成具有中等相似性,优势种均为Formosanochiurusnipponicus和德跳属种1。从杨桦林到云冷杉林,跳虫个体数量逐渐达到较高水平。在阔叶红松林中,跳虫种群丰富度达到最高峰。植物多样性的变化可能会改变凋落物的数量和质量,从而影响土壤动物群落的组成和功能发挥[27-28]。本研究中,对比几种不同林型,阔叶红松林内树种组成丰富,不同林分组成显著增强了环境及空间异质性[29],改变地表凋落物组成,为跳虫群落的生存与发展提供了丰富的食物资源及生存环境条件[30],通过进一步对土壤环境条件的改变,促使土壤内能够容纳更多土壤动物[31-33],群落结构组成更为丰富。
4.2 土壤理化性质对跳虫群落组成结构的影响
不同林型间,大多数跳虫物种与云冷杉林呈显著正相关关系,大部分物种与阔叶红松林呈显著正相关关系,大部分物种与杨桦林呈显著负相关关系。在土壤理化性质中,大多数跳虫物种与土壤pH、土壤还原电位呈显著负相关关系,大多数跳虫物种与土壤碳氮比、土壤温度呈显著正相关关系,大多数跳虫物种与土壤养分无明显关系。由此看出,弹尾目受林型与土壤理化性质影响。针叶林比阔叶林更易形成腐殖质,使凋落物减少,进而对土壤跳虫群落产生影响[34]。在云冷杉林枯落物层中,水分、阳离子浓度(K+、Ca2+、Mg2+)和土壤pH显著降低,使土壤酸化[35],土壤肥力降低[36],进而导致跳虫物种多样性降低。植物通过矿化作用,从深层土壤吸收钙和其他阳离子[37],并通过凋落物沉积将这些阳离子沉积在土壤表面,影响土壤酸度,从而影响了土壤动物的分布[38-40]。土壤温度影响根系生长、水分利用率和微生物活性,进而影响跳虫群落[41]。土壤碳氮比影响有机质的分解难易程度,有些物种受土壤碳氮比影响较大的原因在于低碳氮比引起的土壤有机质的快速转化[42]。植被、土壤和土壤动物之间的变化具有密切的时间同步性,表明这些成分之间的相互作用可能在生态系统的演替变化中发挥重要作用[43]。
