不同林分类型叶片稳定碳、氮同位素的变化特征
2022-03-16马锐豪樊伟王斐夏开温正宇徐小牛
马锐豪 樊伟 王斐 夏开 温正宇 徐小牛


摘要: 为了解各种生物及非生物因素对植物叶片稳定碳、氮同位素含量的影响,研究了蔡家桥林场内马尾松次生林(PM-SF)、湿地松人工林(PE-P)及杉木人工林(CL-P),不同叶龄叶片的养分(C、N、P、K、Ca、Mg)含量及其化学计量比与稳定碳同位素(δ13C)、稳定氮同位素(δ15N)含量的关系,探讨植物叶片δ13C、δ15N含量变异特征。结果表明,各林型叶片δ13C含量随叶龄变化不显著,叶片δ15N含量随叶龄的增大而减小。总体上,马尾松次生林叶片δ13C、δ15N含量均高于湿地松人工林及杉木人工林。多元回归分析结果表明,葉片养分含量、叶龄和林型的共同影响分别解释叶片δ13C、δ15N含量变化的83.94%和83.00%。相关性分析结果表明,叶片δ13C含量与N含量呈显著正相关关系,与C含量/N含量呈显著负相关关系;δ15N含量与N、P、K含量呈显著正相关关系,与Ca含量、C含量/N含量呈显著负相关关系。总体来看,植物叶片对碳、氮同位素的分馏受林分类型、叶龄、营分元素含量等多种因素的影响,不同林型条件下,环境和养分因子的变化在一定程度上反映并影响了植物叶片δ13C、δ15N含量的变化特征。
关键词: 稳定碳、氮同位素;林型;叶龄;营养元素
中图分类号: S718.3 文献标识码: A 文章编号: 1000-4440(2022)01-0102-09
Abstract: In order to understand the effects of various biotic and abiotic factors on contents of stable carbon isotope and stable nitrogen isotope in plant leaves, the relationship between nutrients (C, N, P, K, Ca, Mg) contents and their stoichiometric ratios and the contents of stable carbon isotope (δ13C) and stable nitrogen isotope (δ15N) in leaves at different foliar ages of Pinus massoniana secondary forest (PM-SF), Pinus elliottii artificial forest (PE-P) and Cunninghamia lanceolata artificial forest (CL-P) in Caijiaqiao Forest Farm were analyzed, and the variation characteristics of δ13C and δ15N contents in plant leaves were discussed. The results showed that, with the change of foliar ages, the change of foliar δ13C contents of different forest types was not significant, but the foliar δ15N content of different forest types decreased with the increase of foliar age. The foliar δ13C and δ15N contents of Pinus massoniana secondary forest were higher than those of the two artificial forests on the whole. Multiple regression analysis suggested that, the combined effects of foliar nutrient contents, foliar age and forest type could explain 83.94% and 83.00% of the foliar δ13C and δ15N contents, respectively. Results of the correlation analysis showed that, the foliar δ13C content was in significant positive correlation with N content, and was in significant negative correlation with the ratio of C content and N content; the foliar δ15N content was in significant positive correlation with N content, P content and K content, and was in significant negative correlation with Ca content, C content and the ratio of C content and N content. In conclusion, the fractionation of carbon and nitrogen isotopes in plant leaves is affected by multiple factors including forest type, foliar age and contents of nutrient elements, and the variation of environmental and nutrient factors can reflect and affect the change properties of foliar δ13C and δ15N contents to some extent under different forest types.
Key words: stable carbon and stable nitrogen isotopes;forest type;foliar age;nutrient element
近年来,稳定碳氮同位素作为研究植物生理生态学最有力的工具之一,广泛应用于研究森林演替及植物生长发育不同阶段对陆地生态系统养分循环的潜在影响[1],为研究植物生理过程提供了可靠的手段。碳同位素的分馏广泛存在于陆地生态系统的碳循环过程中。植物体内碳同位素的分馏主要是叶片通过光合作用,对13CO2进行识别并排除的结果。稳定碳同位素(δ13C)作為植物长期水分利用效率的指标,能够表现其一生中气孔导度、CO2同化和蒸腾作用的累积效应,进而表征植物的生长规律及其生理特性[2]。对常绿树种而言,叶龄是控制植物养分积累和吸收的关键因素[3]。在叶片生长发育的过程中,δ13C含量很大程度上会受到养分状况的影响。其中,C、N、P、K、Ca、Mg等叶片功能元素参与蛋白质、脱氧核糖核酸、核糖核酸的合成及叶片器官的形成,间接影响光合作用能力,在叶片δ13C的分馏过程中起着关键作用[4]。
稳定氮同位素(δ15N)技术经常被用于评估植物群落的氮素利用效率、指示生态系统氮循环的模式[5],描述植物种间竞争以及氮素的吸收特征[6],表现植被类型变化后植物体内的氮循环效率。δ15N通常作为反映不同过程中氮素组成的重要示踪剂,用以表现生态系统中植物氮素的利用率、有效性、氮素流失途径及其损失量等[7]。
过去的几十年里,中国经历了巨大的土地利用变化,森林资源不断丰富,包括原始林、次生林及更为符合经营目的的人工林[8]。不同的森林群落不仅会影响养分元素的生物地球化学循环[9],也会影响稳定同位素特征,进而导致植物在生理生态特性、养分需求和生长发育特征等方面的变化[10]。在全球或区域尺度上,已有不少研究关注了自然或人工控制条件下叶片δ13C和δ15N的特征[11-12],而对于不同林分类型(林型),δ13C和δ15N的含量随植物叶片习性、物候特征尤其是随叶龄等中等尺度上变化的研究尚不完整[13]。为此,本研究就中国亚热带马尾松(Pinus massoniana)次生林(PM-SF)、湿地松(Pinus elliottii)人工林(PE-P)、杉木(Cunninghamia lanceolata)人工林(CL-P)3种林型不同叶龄的叶片养分元素及稳定碳、氮同位素的变化特征进行分析,探讨不同林型在中等尺度(即叶龄)上叶片营养状况的改变,进而加深对植物生长发育特点的理解,并通过评估人工林的建立对生态系统中植物营养状况的影响,为森林可持续经营提供科学依据。
1 材料与方法
1.1 试验林场概况
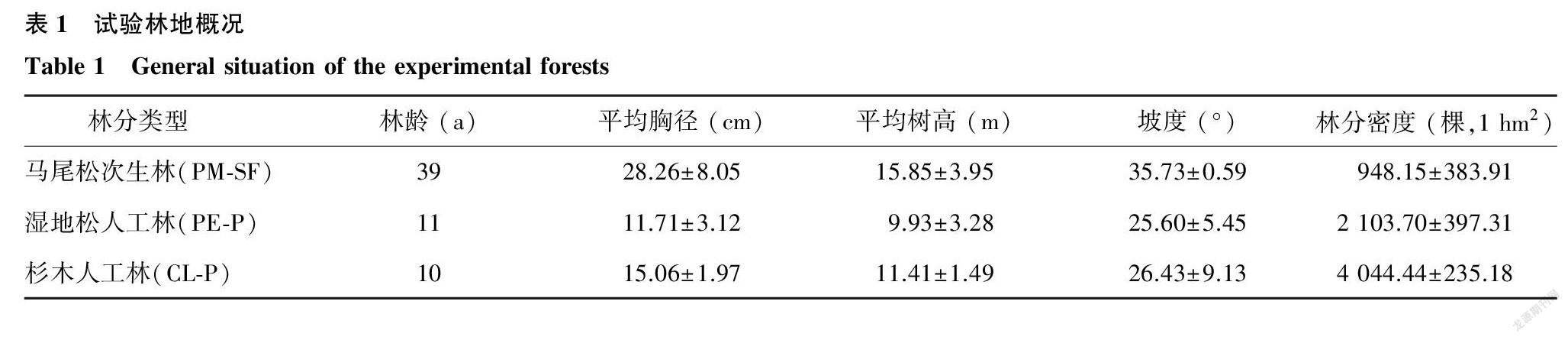
试验地位于安徽省旌德县国营蔡家桥林场,地处黄山北麓(118°30′E,30°20′N),经营总面积为3 576.7 hm2。试验地区属亚热带湿润季风气候,光照充足,四季分明。年平均气温16 ℃,近10年的年平均降水量为1 522.7 mm,年均蒸发量约1 257.0 mm,年均相对湿度70%。土壤以山地黄壤为主,植被丰富,森林覆盖率达85%。地带性植被为亚热带常绿针叶林、阔叶林,经长期森林经营,大面积的马尾松次生林被杉木、湿地松、檫木(Sassafras tzumu)、枫香(Liquidambar formosana)、毛竹(Phyllostachys heterocycla)等人工植被所替代。
2020年8月,在蔡家桥林场内选取马尾松次生林及其采伐后营造的杉木、湿地松人工林为研究对象,选择立地条件相似的林型,每个林型设置3个20 m×20 m独立的重复样地,共9个样地,用于植物样本的采集。各林型样地概况如表1所示。
1.2 植物鲜叶的采集与处理
在每个样地内进行每木测量,在此基础上,根据平均标准木法在各样地内随机选取4~5棵健康的标准木,用高枝剪从树冠的上部、中部和下部沿4个方向收集马尾松、杉木、湿地松的活枝,运用“节点法”按照不同叶龄(当年生、一年生、二年生)采集鲜叶[14]。分别将每个样地内相同叶龄的新鲜叶片混合装袋并带回实验室,将鲜叶放入65 ℃的烘箱中烘干至恒质量(>48 h),然后使用微型植物粉碎机磨碎后过100目筛,放入自封袋中保存后待测。
利用同位素质谱仪(Thermo Scientific Delta V Advantage,Germany)联用的元素分析仪(EA-IRMS)系统测定植物叶片样品的稳定碳同位素(δ13C)含量和稳定氮同位素(δ15N)含量:ω(δ13C)=(RSP/Rsd-1)×100%,ω(δ15N)=(RSP/Rsd-1)×100%,式中,R为13C/12C或15N/14N,Rsp为样品的R值,Rsd为标准物质的R值;碳同位素以国际标准物质PDB(Pee dee belemnite)为标准,氮同位素以标准大气中的N2为标准。稳定碳、氮同位素组成的内标精密度(标准差)分别优于0.08%和0.020%。
样品碳(C)和氮(N)含量使用元素分析仪(EA3000 Vector, Italy)测定。叶片磷(P)、钾(K)、钙(Ca)和镁(Mg)含量用HNO3∶HClO4=3∶1(体积比)法消煮后,分别用流动注射分析仪(FIA Star 5000, FOSS, Denmark)和原子吸收分光光度计(AA6300, Shimadzu, Japan)进行测定。
1.3 数据处理
根据叶龄(当年生叶龄、1年生叶龄、2年生叶龄)和林分类型(马尾松次生林、湿地松人工林、杉木人工林)分组,运用Excel 2013对数据进行整理,计算所有参数的平均值和标准差。采用双因素方差分析和Tukey检验,比较叶片δ13C、δ15N含量及各营分元素在不同叶龄和林型中的差异。通过单因素方差分析和LSD检验分析林型和叶龄对植物叶片营分元素及δ13C、δ15N的影响,随后采用逐步多元回归法评估各因子的相对重要性并建立回归模型。运用Pearson相关系数来检验各因子之间的相关性。使用软件R 3.6.1和SPSS 22.0进行统计分析,并用软件Origin 2018绘制图表。
2 结果与分析
2.1 林型和叶龄对叶片δ13C、δ15N含量的影响
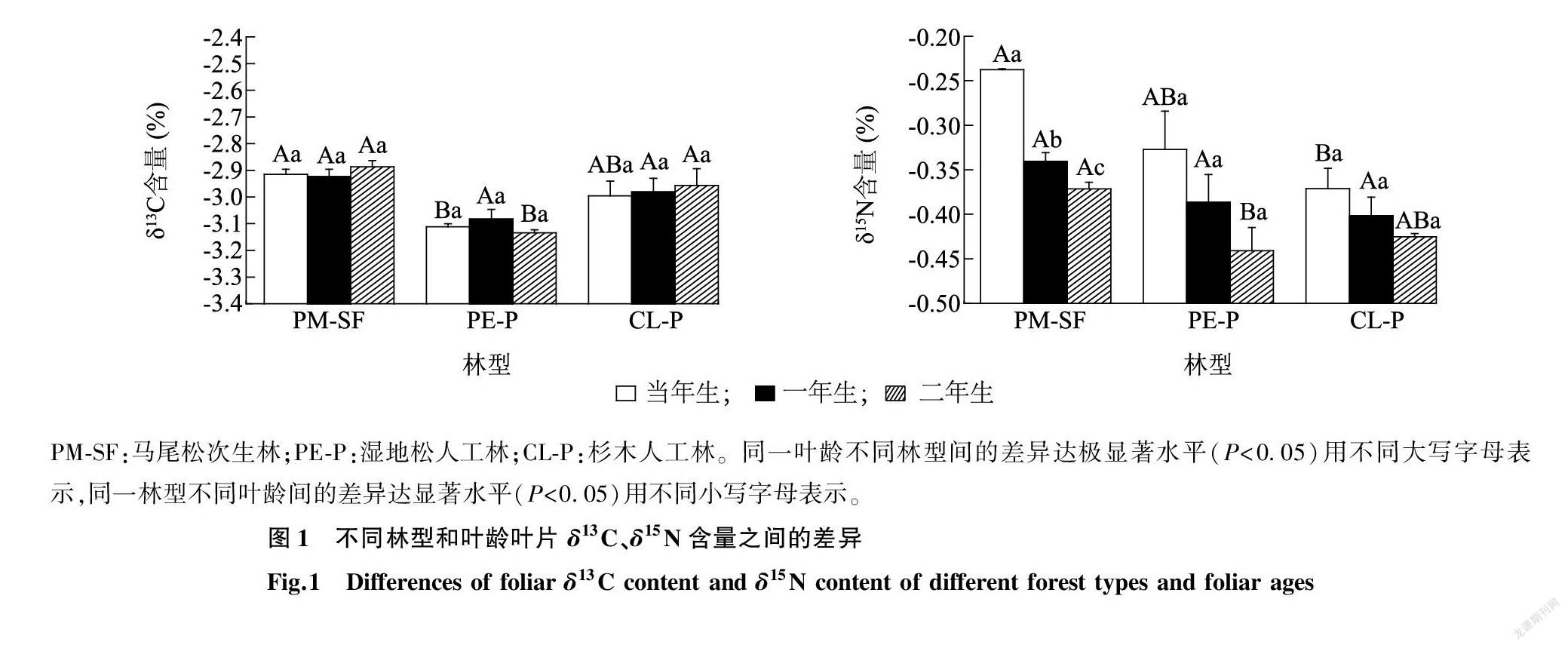
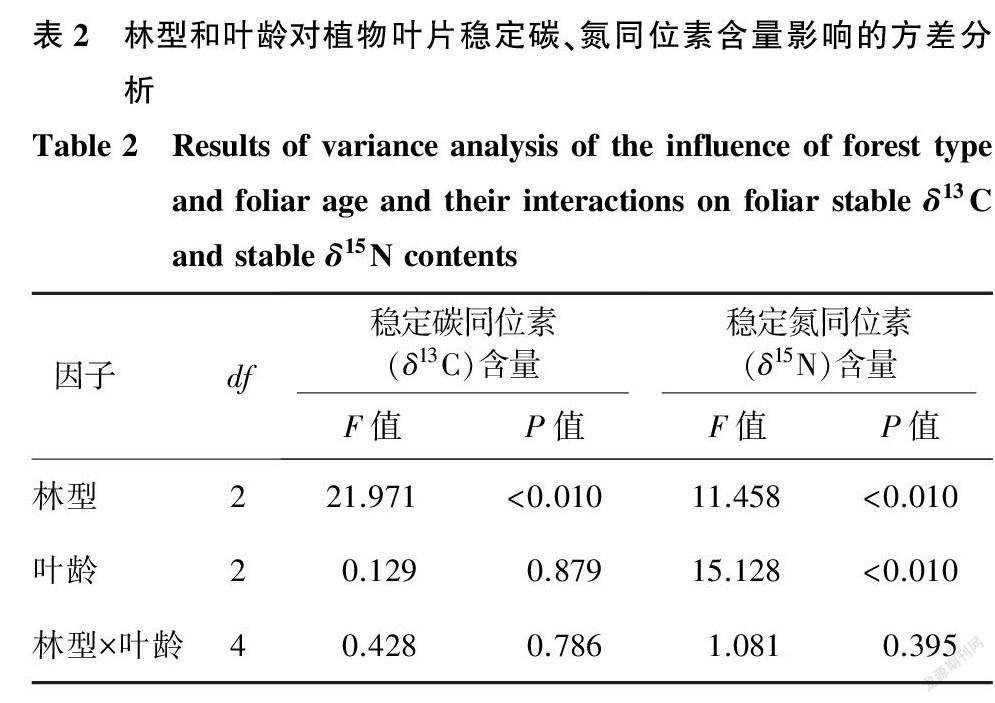
林型对叶片δ13C含量呈极显著的影响(P<0.01)。当年生叶中,马尾松δ13C含量(-29.15‰)显著高于湿地松(-3.112%);一年生叶中,各林型的叶片δ13C含量无显著差异;二年生叶中,湿地松叶片δ13C含量(-3.134%)显著低于马尾松(-2.887%)和杉木(-2.957%)(图1)。总体上,叶片δ13C含量呈马尾松(-2.909%)>杉木(-2.978%)>湿地松(-3.109%)的趋势。叶龄对叶片δ13C含量无显著影响(P=0.879)(表2),随着叶龄的增加各林型植物叶片δ13C含量变化都不相同。
林型和叶龄对叶片δ15N含量都呈极显著的影响(P<0.01)(表2)。当年生叶中,马尾松叶片δ15N含量(-0.237%)和湿地松叶片δ15N含量(-0.327%)显著高于杉木(-0.371%);而一年生叶中,各林型植物叶片δ15N含量并无显著差异;在二年生叶中,马尾松叶片δ15N含量(-0.371%)和杉木叶片δ15N含量(-0.425%)显著高于湿地松叶片δ15N含量(-0.441%)(图1)。各林型叶片δ15N含量都随叶龄的增大而减小,除马尾松次生林各叶龄叶片δ15N含量差异显著外,湿地松人工林和杉木人工林各叶龄叶片δ15N含量差异均不显著。林型与叶龄的交互作用对植物叶片稳定碳、氮同位素含量均无显著影响(P>0.05)(表2)。
2.2 林型和叶龄对叶片营养元素含量的影响
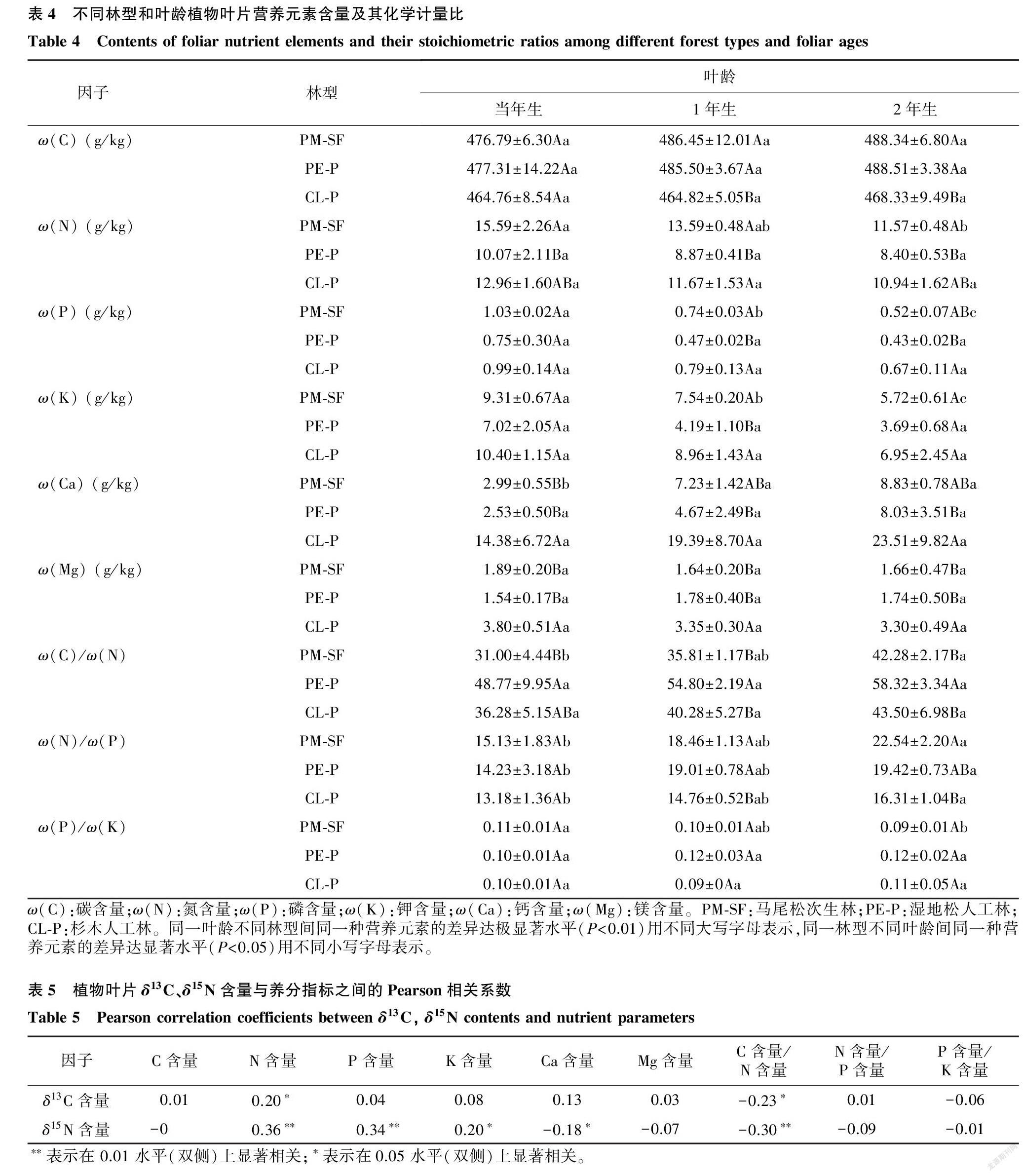
林型对叶片营养元素C、N、P、K、Ca、Mg含量均有极显著的影响(P<0.01)。叶龄对叶片C含量(P=0.108)、Mg含量(P=0.584)无显著影响(表3)。除当年生叶片外,杉木叶片C含量显著低于马尾松及湿地松叶片C含量,而杉木各叶龄叶片Mg含量均显著高于马尾松及湿地松叶片(表4)。林型和叶龄对叶片N、P、K、Ca含量均有显著影响(P<0.01)(表3),且在同一林型中随叶龄增大而减小。林型和叶龄的交互作用对叶片C含量(P=0.852)、N含量(P=0.651)、P含量(P=0.581)、K含量(P=0.848)、Ca含量(P=0.961)和Mg含量(P=0.504)均无显著影响(表4)。
林型和叶龄对叶片ω(C)/ω(N)和ω(N)/ω(P)比值均有极显著影响(P<0.01),在同一林型下随叶龄的增加而增加。而同一叶龄下,湿地松叶片的ω(C)/ω(N)显著高于马尾松和杉木,总体上呈湿地松(53.96)>杉木(40.02)>马尾松(36.36)的趋势;除当年生叶片外,同一叶龄不同林型的叶片ω(N)/ω(P)均有显著差异(表4)。林型和叶龄对叶片ω(P)/ω(K)无显著影响(P=0.276,P=0.884)(表3)。林型和叶龄的交互作用对叶片ω(C)/ω(N)(P=0.958)、ω(N)/ω(P)(P=0.135)和ω(P)/ω(K)(P=0.549)均无显著影响(表3)。
2.3 叶片δ13C、δ15N含量与养分因子之间的关系
对叶片δ13C、δ15N含量与养分因子进行相关性分析,结果如表5所示。叶片δ13C含量与ω(C)/ω(N)呈显著负相关关系,与N含量呈显著正相关关系。叶片δ15N含量与N、P含量呈极显著正相关关系,与K含量呈显著正相关关系;叶片δ15N含量与Ca含量呈显著负相关关系,与ω(C)/ω(N)呈极显著负相关关系。
叶片C、N、P、K含量间在各林型及叶龄下都有极显著的相关性,而叶片中Mg含量只与C含量(负相关)和Ca含量(正相关)有极显著的相关性,叶片Ca含量只与Mg含量有极显著的相关性(图2)。表明叶片中的Mg和Ca含量与其他养分含量没有较强关联。
2.4 林型、叶龄和叶片养分含量对δ13C、δ15N含量的相对重要性
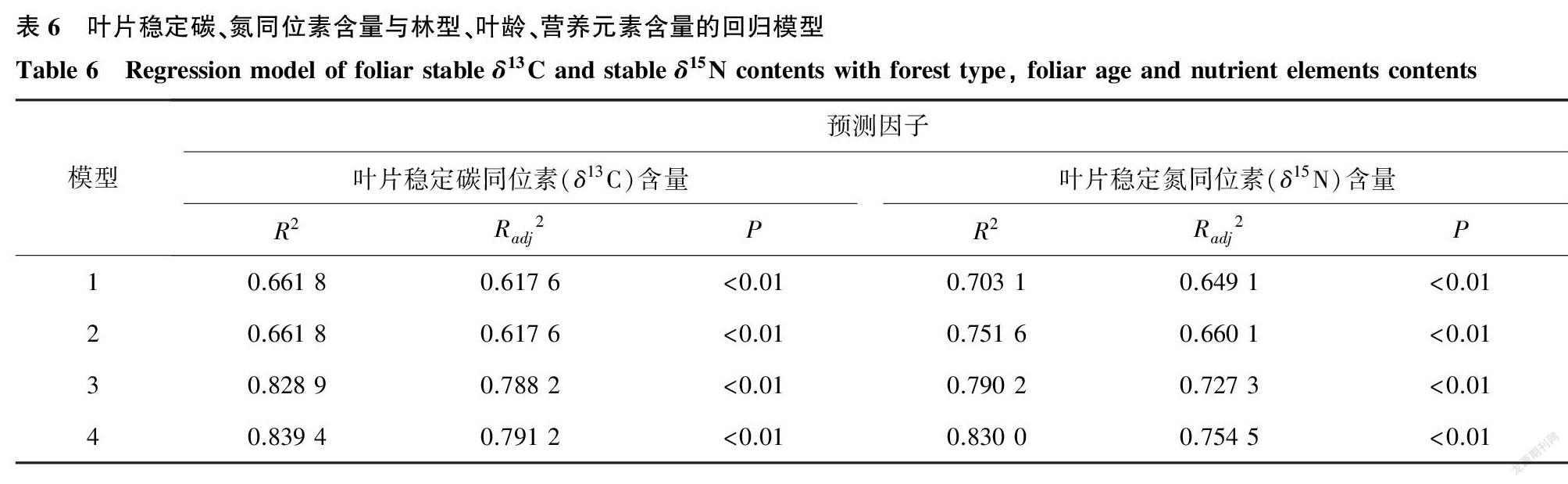
模型1至模型4分别以养分因子、养分因子+叶龄、养分因子+林型、养分因子+叶龄+林型为自变量,以叶片δ13C、δ15N含量为因变量建立多元回归模型,采用逐步回归法尽可能消除其共线性的潜在影响(模型自动纳入P<0.05的变量,排除P>0.10的变量)。总体上,4个多元回归模型对叶片δ13C含量的解释度为模型4(83.94%)>模型3(82.89%)>模型2(66.18%)=模型1(66.18%)。模型2与模型1对δ13C含量的解释度相同,表明当叶龄与养分因子共同作为自变量时,仅有养分因子对δ13C含量具有解释作用。4个模型对叶片δ15N的解释度为模型4(83.00%)>模型3(79.02%)>模型2(75.16%)>模型1(70.31%)(表6)。上述结果中,模型4即以养分因子+叶龄+林型为因变量时,对叶片δ13C、δ15N含量的解释度最大,表明了林型与叶龄对于解释叶片δ13C、δ15N含量的变化具有一定的重要性。
3 讨论
3.1 林型和叶龄对叶片δ13C含量的影响
本研究中,3种林分叶片δ13C含量为-3.157%~-2.840%,且林型对植物叶片δ13C含量的影响极显著。以往研究结果表明,C3植物光合作用过程中碳同位素的分馏导致叶片δ13C含量介于-3.2%~-2.2%[15],与本研究所测得的叶片δ13C含量范围基本一致。植物在光合作用过程中分馏较重的同位素13C,尤其是叶片细胞质优先吸收12C,导致植物叶片胞间13C贫化,使叶片中的碳同位素比率(13C/12C)低于大气中CO2的碳同位素比率[16]。植物对碳同位素的分馏作用使叶片δ13C含量与植物细胞内外CO2浓度之比(Ci/Ca)呈线性相关,并受叶片电导率、RuBP羧化效率和其他影响植物生理生化的环境因素的影响[17]。因此,不同植物的光合能力以及它们对环境的耦合反应不同,其叶片δ13C含量会有所差异[18]。此外,叶片δ13C含量在很大程度上受到植物的生活型和系統发育类型的影响[19]。本研究中,林型对叶片δ13C含量的影响极显著(P<0.01)。主要是由于湿地松人工林及杉木人工林均为马尾松天然林间伐后重建的,与马尾松次生林相比,2种人工林不同的管理措施以及其本身具有高度的立地特异性,导致3个林型间林分结构、林龄、树高、林分密度等均有较大差异,从而对叶片δ13C含量产生了较大的影响。
以往研究结果已经证实,叶片δ13C含量随叶龄的增大而下降[20]。新生叶片生长相对较快,需要合成大量有机物以满足其生长发育的需求,导致叶片细胞间CO2浓度处于饥饿状态,对13CO2的分馏减少;而在叶片的外部形态和生理代谢功能趋于成熟后,植物逐渐能够有效辨别并排除13CO2[21]。因此,新生叶片的13C更加富集,成熟甚至衰老叶片中的13C相对贫化。然而,植物对13CO2的分馏不仅受其自身新陈代谢影响,降水、光照等环境条件,也能够影响植物固定CO2过程中对同位素的辨别[22]。本研究取样时间为2020年8月底,当年5-6月,研究地刚受到洪涝灾害(987.2 mm)。洪水通过降低植物根系导水率,降低了叶水势从而导致叶片气孔导度下降,当植物受到短时间持续的洪涝影响时,气孔导度的降低对叶片δ13C含量的影响较大,导致植物叶片对13C的辨别排除减少,使叶片δ13C含量上升[23]。因此,由于非生物和生物因子之间复杂的相互作用,导致本研究中叶龄对叶片δ13C含量无显著影响,且随叶龄的增大叶片δ13C含量并无明显的变化趋势。
3.2 林型和叶龄对叶片δ15N含量的影响
本研究中,林型和叶龄对叶片δ15N含量均有极显著的影响。总体上,马尾松次生林各叶龄叶片δ15N含量高于2个人工林。植物叶片δ15N的变化特征能够表现出生态系统短期内的氮素有效性及其运转轨迹[24]。当土壤的氮素供应相对于植物需求更高时,氮会以无机氮的形式在土壤氮库中积累,较大的土壤NH+4库增加了以NH3形式的挥发、反硝化产生的N2挥发及不完全硝化产生的NO-3淋失,从而加大土壤氮同位素的分馏,增加土壤无机氮库中δ15N含量,植物δ15N含量也会随着土壤中δ15N含量的增加而增加[25]。Bai等[26]的研究結果表明,间伐林分的叶片δ15N含量高于未间伐林分。此外,间伐后的林分冠层密度下降,林地内留存下来的植物对氮素的吸收增多,导致叶片δ15N富集[27]。本研究中经砍伐后(马尾松次生林)的林分密度降低,土壤氮素供应相对于植物需求更高,较高的氮素吸收使得马尾松叶片内δ15N含量升高,因此马尾松次生林叶片δ15N含量显著高于湿地松及杉木人工林。另一方面,植物通过减少从菌根真菌中吸取养分所消耗的氮来降低其对菌根真菌的依赖性,植物的氮素利用率越高对菌根真菌的依赖性就越弱[28]。杉木为具有丛枝菌根的浅根系树种,对氮素的吸收能力弱于马尾松及湿地松这类具有外生菌根且深根系树种,进一步证实了叶片δ15N含量与植物的氮素吸收能力密切相关[29]。
植物对碳同位素的分馏是通过对空气中CO2的固定或内部碳水化合物的转移进行,而氮同位素的分馏则是植物通过对氮素的同化及再分配引起的[30]。植物在对无机氮的同化过程中,较重的同位素15N被“识别”并结合为化合物在内部进行转移。本研究中,各林分叶片δ15N含量随叶龄的增加而减少。常绿针叶树种的叶片木质素含量通常随叶龄的增加而增加,木质化细胞壁厚度的增加导致植物组织内部运输养分的能力降低[31]。因此,对于常绿树种而言,叶龄是控制植物养分积累和吸收的关键因素。此外,本研究中,叶片δ15N与N含量呈极显著正相关关系,而氮作为植物进行光合作用的关键营养元素,通常从成熟叶片转移到更年轻的叶片中[32]。有研究结果表明,植物叶片在生长过程中其主要的含氮化合物会发生显著变化,如蛋白质、叶绿素等随叶片衰老逐渐被降解[33]。在这样的降解过程中,较轻的氮同位素(14N)更易参与反应而贫化,同时,随着基质不断降解(如叶绿体酶的高级降解),化合物中δ15N丰富的部分将被输送至更年轻的叶片中,导致15N富集。
3.3 叶片δ13C、δ15N含量与养分含量的关系
近年来,许多研究者开始关注叶片δ13C含量与养分之间的关系,并发现叶片营养元素含量与气孔导度或光合能力有关,因此它们可能与δ13C含量也存在某种关联。本研究中,叶片N含量与δ13C含量呈显著正相关关系,表明植物的光合作用能力随叶片氮浓度的升高而增强[34]。叶片ω(C)/ω(N)比值能够表现植物对C、N的吸收与利用,反映植物的生长及健康状况[35]。本研究中叶片δ13C含量与ω(C)/ω(N)呈显著负相关关系,与以往研究结果一致,表明植物通过降低氮素利用效率(NUE)获取更高的水分利用效率(WUE)[36]。
植物中氮素的有效性以及植物吸收氮素后的分馏作用被认为是影响叶片δ15N含量的重要因素[37]。N和P同是植物的基本元素,也是各类蛋白质和遗传物质的重要组成元素,具有相似的生化途径[38],且在植物叶片中共同参与如光合作用、蒸腾作用、呼吸作用等生理生化过程[39]。Verboom等[40]的研究中,叶片P和K含量及其比值表现出密切联系,植物对P和K的吸收影响了叶片N含量,并能够通过植物叶片中的养分含量关系表现出来。Ca元素参与合成植物的细胞壁,且在韧皮部中的流动性较差,更易在细胞中积累,能够稳定表现植物叶片生长发育过程中养分的变化[41]。本研究中,叶片δ15N含量与N、P、K含量之间呈显著正相关关系,与Ca含量之间呈显著负相关关系,表明了不同林型间叶片δ15N含量的差异能够反映植物营养元素含量的变化。
由多元线性回归方程模型可知,影响δ13C含量的营养元素及其化学计量比为C、Ca、ω(C)/ω(N),影响δ15N含量的为P、K、Mg、Ca、ω(P)/ω(K),与相关性分析结果并不完全相同。这可能是由于林分类型与营养元素对叶片δ13C、δ15N含量的影响是交互的。本研究中林型对C、N、P、K、Ca、Mg含量及化学计量比ω(C)/ω(N)、ω(N)/ω(P)的影响均极显著,且营养元素之间同样存在较强的相互影响。表明林型与营养元素之间的相互作用可能直接影响叶片δ13C、δ15N含量,也可能分别通过影响植物的水分利用效率或叶片N含量,间接影响叶片δ13C、δ15N含量[42-43]。
4 结论
通过对蔡家桥林场马尾松次生林、湿地松人工林及杉木人工林的植物叶片营养元素含量、稳定碳、氮同位素含量及各种影响因子进行分析,得出以下结论:(1)不同林分类型及自然或人工控制条件下的植物δ13C分馏情况不同,且林型对叶片δ13C含量有极显著的影响;(2)各林分叶片δ15N含量总体呈马尾松次生林>湿地松人工林>杉木人工林的趋势,各林分叶片δ15N含量均随叶龄的增大而显著减小;(3)叶片养分含量对δ13C、δ15N含量的影响较为显著,且养分间的相互作用能够通过碳、氮同位素含量反映;(4)根据多元回归模型及相关性进行的相对重要性分析,林型和叶片ω(C)/ω(N)、N含量对叶片δ13C含量的变异起主导作用,叶龄、林型、ω(C)/ω(N)及N、P、K、Ca含量等因子对叶片δ15N含量有显著或极显著影响。综合而言,叶片δ13C含量与环境因素关系密切,植物应对环境产生的变化或与环境因素产生的耦合作用能够通过叶片δ13C含量反映,而叶片δ15N含量则反映出了植物体内的养分含量变化规律。
参考文献:
[1] GATICA M G, ARANIBAR J N, PUCHETA E. Environmental and species-specific controls on δ13C and δ15N in dominant woody plants from central-western Argentinian drylands[J]. Austral Ecology, 2017, 42(5): 533-543.
[2] 葛露露,孟庆权,林 宇,等. 滨海沙地不同树种人工林叶片和土壤表层稳定碳氮同位素及水分利用效率研究[J]. 西北植物學报, 2018, 38(3): 544-552.
[3] PERAKIS S S, TEPLEY A J, COMPTON J E. Disturbance and topography shape nitrogen availability and δ15N over long-term forest succession[J]. Ecosystems, 2015, 18(4): 573-588.
[4] LI C, WU C, DUAN B, et al. Age-related nutrient content and carbon isotope composition in the leaves and branches of Quercus aquifolioides along an altitudinal gradient[J]. Trees, 2009, 23(5): 1109-1121.
[5] 赵维俊,刘贤德,金 铭,等. 祁连山青海云杉林叶片-枯落物-土壤的碳氮磷生态化学计量特征[J]. 土壤学报, 2016, 53(2): 477-489.
[6] 张秋芳,谢锦升,陈奶寿,等. 生态恢复对马尾松叶片化学计量及氮磷转移的影响[J]. 生态学报, 2017, 37(1): 267-276.
[7] BURNHAM M B, ADAMS M B, PETERJOHN W T. Assessing tree ring delta δ15N of four temperate deciduous species as an indicator of N availability using independent long-term records at the Fernow Experimental Forest, WV[J]. Oecologia, 2019, 191(4): 971-981.
[8] 王宝荣,曾全超,安韶山,等. 黄土高原子午岭林区两种天然次生林植物叶片-凋落叶-土壤生态化学计量特征[J]. 生态学报, 2017, 37(16): 5461-5473.
[9] CAO Y, CHEN Y. Coupling of plant and soil C∶N∶P stoichiometry in black locust (Robinia pseudoacacia) plantations on the Loess Plateau, China[J]. Trees, 2017, 31(5): 1559-1570.
[10]WANG Q, LI F, RONG X, et al. Plant-soil properties associated with nitrogen mineralization: Effect of conversion of natural secondary forests to larch plantations in a headwater catchment in Northeast China[J]. Forests, 2018, 9(7): 386.
[11]KEELING R F, GRAVEN H D, WELP L R, et al. Atmospheric evidence for a global secular increase in carbon isotopic discrimination of land photosynthesis[J]. Proceedings of The National Academy of Sciences of The United States of America, 2017, 114(39): 10361-10366.
[12]哈丽古丽·艾尼,伊丽米努尔,管文轲,等. 不同生境胡杨叶片δ13C和δ15N及其对环境因子的响应[J]. 西北植物学报, 2020, 40(6): 1031-1042.
[13]熊 鑫,张慧玲,吴建平,等. 鼎湖山森林演替序列植物-土壤碳氮同位素特征[J]. 植物生态学报, 2016, 40(6): 533-542.
[14]KIKUZAWA K, LECHOWICZ M J. Quantifying leaf longevity[M]. Tokyo: Springer, 2011:23-29.
[15]BARREZUETA UNDA S, PAZ GONZLEZ A, LUNA ROMERO E, et al. Variability of δ13C and δ15N in cocoa cultivars, in the province of El Oro, Ecuador [J]. Terra Latinoamericana, 2019, 37(2): 131-140.
[16]葛體达,王东东,祝贞科,等. 碳同位素示踪技术及其在陆地生态系统碳循环研究中的应用与展望[J]. 植物生态学报, 2020, 44(4): 360-372.
[17]郜士垒,何宗明,黄志群,等. 杉木宿存叶片的分解及稳定性碳氮同位素和化学组成[J]. 生态学杂志, 2015, 34(9): 2457-2463.
[18]JIA Y, WANG G, TAN Q, et al. Temperature exerts no influence on organic matter δ13C of surface soil along the 400 mm isopleth of mean annual precipitation in China[J]. Biogeosciences, 2016, 13(17): 5057-5064.
[19]刘建锋,张玉婷,倪妍妍,等. 栓皮栎叶片δ13C和δ15N的纬向趋势及其影响因子[J]. 应用生态学报, 2018, 29(5): 1373-1380.
[20]CERNUSAK L A, TCHERKEZ G, KEITEL C, et al. Viewpoint: Why are non-photosynthetic tissues generally 13C enriched compared with leaves in C3 plants? Review and synthesis of current hypotheses[J]. Functional Plant Biology, 2009, 36(3): 199-213.
[21]LI C, WANG B, CHEN T, et al. Leaf age compared to tree age plays a dominant role in leaf δ13C and δ15N of Qinghai Spruce (Picea crassifolia Kom.)[J]. Forests, 2019, 10(4): 310.
[22]GERSCHLAUER F, SAIZ G, SCHELLENBERGER COSTA D, et al. Stable carbon and nitrogen isotopic composition of leaves, litter, and soils of various ecosystems along an elevational and land-use gradient at Mount Kilimanjaro, Tanzania[J]. Biogeosciences, 2019, 16(2): 409-424.
[23]FAN R, MOROZUMI T, MAXIMOV T C, et al. Effect of floods on the δ13C values in plant leaves: a study of willows in Northeastern Siberia[J]. Peer J, 2018, 6: e5374.
[24]张金美,张 萌,匡武名,等. 水华条件下鄱阳湖区植物叶片碳氮同位素特性[J]. 环境科学研究, 2016, 29(5): 708-715.
[25]董 雪,李永华,辛智鸣,等. 唐古特白刺叶性状及叶片δ13C、δ15N沿降水梯度的变化特征[J]. 生态学报, 2019, 39(10): 3700-3709.
[26]BAI S H, DEMPSEY R, REVERCHON F, et al. Effects of forest thinning on soil-plant carbon and nitrogen dynamics[J]. Plant and Soil, 2016, 411(1/2): 437-449.
[27]QIU S, BELL R W, HOBBS R J, et al. Overstorey and juvenile response to thinning and drought in a jarrah (Eucalyptus marginata Donn ex Sm.) forest of southwestern Australia[J]. Plant and Soil, 2012, 365(1/2): 291-305.
[28]HOGBERG P, JOHANNISSON C, YARWOOD S, et al. Recovery of ectomycorrhiza after ′nitrogen saturation of a conifer forest[J]. New Phytol, 2011, 189(2): 515-525.
[29]CRAINE J M, BROOKSHIRE E N J, CRAMER M D, et al. Ecological interpretations of nitrogen isotope ratios of terrestrial plants and soils[J]. Plant and Soil, 2015, 396(1/2): 1-26.
[30]方运霆,刘冬伟,朱飞飞,等. 氮稳定同位素技术在陆地生态系统氮循环研究中的应用[J]. 植物生态学报, 2020, 44(4): 373-383.
[31]YAN T, L X T, ZHU J J, et al. Changes in nitrogen and phosphorus cycling suggest a transition to phosphorus limitation with the stand development of larch plantations[J]. Plant and Soil, 2017, 422(1/2): 385-396.
[32]TIAN D, YAN Z, NIKLAS K J, et al. Global leaf nitrogen and phosphorus stoichiometry and their scaling exponent[J]. National Science Review, 2018, 5(5): 728-739.
[33]蔣 龙,徐振锋,吴福忠,等. 亚热带3种典型常绿森林土壤和植物叶片碳氮磷化学计量特征[J]. 应用与环境生物学报, 2019, 25(4): 759-767.
[34]WALIA A, GUY R D, WHITE B. Carbon isotope discrimination in western hemlock and its relationship to mineral nutrition and growth[J]. Tree Physiol, 2010, 30(6): 728-740.
[35]GE J, XIE Z. Leaf litter carbon, nitrogen, and phosphorus stoichiometric patterns as related to climatic factors and leaf habits across Chinese broad-leaved tree species[J]. Plant Ecology, 2017, 218(9): 1063-1076.
[36]司高月,李晓玉,程淑兰,等. 长白山垂直带森林叶片-凋落物-土壤连续体有机碳动态——基于稳定性碳同位素分析[J]. 生态学报, 2017, 37(16): 5285-5293.
[37]GAUTAM M K, LEE K S, SONG B Y, et al. Early-stage changes in natural 13C and 15N abundance and nutrient dynamics during different litter decomposition[J]. Journal of Plant Research, 2016, 129(3): 463-476.
[38]PANG Y, TIAN J, ZHAO X, et al. The linkages of plant, litter and soil C∶N∶P stoichiometry and nutrient stock in different secondary mixed forest types in the Qinling Mountains, China[J]. Peer J, 2020, 8: e9274.
[39]TONG R, ZHOU B, JIANG L, et al. Leaf nitrogen and phosphorus stoichiometry of Chinese fir plantations across China: A meta-analysis[J]. Forests, 2019, 10(11): 945.
[40]VERBOOM G A, STOCK W D, CRAMER M D. Specialization to extremely low-nutrient soils limits the nutritional adaptability of plant lineages[J]. The American Naturalist, 2017, 189(6): 684-699.
[41]ACHAT D L, POUSSE N, NICOLAS M, et al. Nutrient remobilization in tree foliage as affected by soil nutrients and leaf life span[J]. Ecological Monographs, 2018, 88(3): 408-428.
[42]CHEN C, WU Y, WANG S, et al. Relationships between leaf δ15N and leaf metallic nutrients[J]. Rapid Communications in Mass Spectrometry, 2021, 35(2): e8970.
[43]李善家,张有福,陈 拓. 西北油松叶片δ13C特征与环境因子和叶片矿质元素的关系[J]. 植物生态学报, 2011, 35(6): 596-604.
(责任编辑:张震林)
1544500783237
