基于线粒体COI 序列的矛尾复虾虎鱼5 个野生群体的遗传多样性分析
2021-02-16祝斐应博凯贾超峰孟乾高波孙瑞健宋双双徐大凤卢瑶瑶张志伟陈淑吟张志勇陈心
祝斐,应博凯,贾超峰,孟乾,高波,孙瑞健,宋双双,徐大凤,卢瑶瑶,张志伟,陈淑吟,张志勇,陈心
(1. 江苏省海洋水产研究所,江苏 南通 226000;2. 南通大学 生命科学学院实验室,江苏 南通 226000)
矛尾复虾虎鱼(Synechogobius hasta) 隶属于鲈形目(Perciformes)、虾虎鱼科(Gobiidae)、复虾虎鱼属(Synechogobius),其具有广温广盐的特点,在中国、韩国、日本乃至印度尼西亚等沿海地区的浅海海区均有分布(陈大刚,1975)。矛尾复虾虎鱼不仅个体大(最大可长至500g 以上),且无肌间刺、味道极其鲜美(杨海峰等,2013)。随着近海开发和过度捕捞的加剧,矛尾复虾虎鱼栖息产卵场受到破坏,野生资源量急剧减少。为掌握野生矛尾复虾虎鱼种质资源现状,亟须开展沿海矛尾复虾虎鱼群体的遗传多样性评估。
线粒体DNA 因其母系遗传、进化速率快且不易发生重组的特点,已在水产动物种质资源评价及群体遗传结构研究中发挥了重要作用(陈星等,2012)。对矛尾复虾虎鱼的研究主要集中在基础生物学(张家旭等,2021;孟宽宽等,2017)和苗种繁育(黄金田等,2018;李莉等,2012)等方面,种质资源分析及遗传多样性评价方面的内容仅在早些年有过报道。Song 等(2010)研究表明东营、赣榆等地的核苷酸多样性较低,无高水平遗传多样性,矛尾复虾虎鱼的资源状况不容乐观。宋娜等(2011)对矛尾复虾虎鱼群体遗传多样性的研究结果显示:其各群体相互混杂,丹东与天津群体没有各自特有的单倍型序列分支;此外,浙江沿海的4个地区中只有舟山有少量群体特有的单倍型序列,该6 个群体单倍型多样性处于中等水平,无法维持高水平遗传多样性。本研究采用线粒体COI 分子标记对长江口、宁波、南通如东、连云港和大连5 个矛尾复虾虎鱼群体进行遗传多样性分析并探讨其产生不同地理分布格局的原因,为保证矛尾复虾虎鱼资源的可持续利用和开发提供理论依据。
1 材料与方法
1.1 材料
矛尾复虾虎鱼于2019 年11 月分别购自宁波(采样海区:121.70毅E,30.06毅N)、长江入海口(采样海区:121.73毅E,31.33毅N)、如东(采样海区:121.45毅E,32.54毅N)、连云港(连云港海鲜批发市场:119.14毅E,34.84毅N) 和大连(大连海鲜市场:121.26毅E,38.81毅N),共5 个野生群体,每个群体各30 尾样品(图1)。

图1 矛尾复虾虎鱼群体的采样地点
1.2 方法
1.2.1 样品的制备 将采集到的样品剪取适量尾鳍装入盛有无水乙醇的塑料离心管中并编号,在-20 益冰箱中保存备用。
1.2.2 样品DNA 的提取 使用细胞/组织基因DNA 提取试剂盒(GK0122 上海捷瑞生物工程有限公司) 提取总gDNA,每个样品50 滋L。样品在1.0%的琼脂糖凝胶电泳上进行检测,验证高分子量DNA 提取的质量。用紫外分光光度计测定DNA浓度后,于4 益下保存备用。
1.2.3 PCR 扩增与测序 用Premier 5.0 设计一对引物,上游引物序列ZF1COIF(aat acc aga cgc ccc tat),下游引物序列ZF1/2COIR(gga aga tga acc cca gtg)。PCR 的反应体系为25 滋L:2.5 滋L 10 伊PCR buffer,0.5 滋L dNTP Mix,1 滋L Primer ZF2COIF,1 滋L PrimerZF1/2COIR,0.5 滋L Pfu DNA polymerase,2 滋L DNA 模板,用ddH2O 补足至25 滋L(即加入17.5滋LddH2O)。PCR 反应程序如下:在95 益下预变性5 min;在95 益下变性45 s,在50益下退火45 s,在72 益下延伸1 min,共进行35 个循环;最后在72 益下延伸5 min,PCR 产物于4 益保存。PCR 产物经1%琼脂糖凝胶电泳检测后,送至上海生工生物工程有限公司进行测序。
1.3 数据处理
测序完成后,利用ClustalW 对测序结果进行校对与校正。用DnaSP 5(Librado et al,2009)计算各群体单倍型数、单倍型多样性、核苷酸多样性、平均核苷酸差异、Tajima忆s D 值和Fu忆s Fs 值。利用Arlequin 3.5 软件计算两两群体间的遗传分化指数(FST) (Excoffier et al,2005),遗传变异和地理遗传结构分析采用分子方差分析(Analysis of molecular variance,AMOVA);利用MEGA 5.1 软件(Tamura et al,2011)分析序列的碱基组成,计算群体内及群体间的遗传距离;采用邻接法(Neighbor-Joining,NJ) (Saitou et al,1987),并基于Kimura 2-parameter model 构建矛尾复虾虎鱼单倍型邻接关系进化树(即绘制NJ 邻接树),采用Bootstrap(重复次数1000)检验各分支置信度;利用Network 10.2 软件(Bandelt et al,1999)构建单倍型网络及进化关系图。
2 结果
2.1 序列变异分析
利用NCBI 数据库的Blast 比对,确定COI 的有效片段为427bp。这5 个群体的427bp COI 片段,共检测到15 个变异位点,占总位点数的3.51%,其中简约信息位点11 个,单独位点4 个,变异均为转换或颠换,转换明显多于颠换,没有检测到插入或缺失现象(表1)。COI 序列中碱基A、T、G、C 的平均含量分别为24.6%、29.6%、20.1%、25.7%,其中T 碱基含量最高,G 碱基含量最低。A+T 的平均含量(54.2%)>C+G 的平均含量(45.8%),表明COI 序列具有AT 偏好性。

表1 矛尾复虾虎鱼线粒体COI 基因序列变异位点
2.2 单倍型在群体中的分布及遗传关系
在150 尾矛尾复虾虎鱼中共检测出17 个单倍型,其中5 个为共享单倍型,其余12 个为独有单倍型。单倍型Hap2 为5 个群体的共享单倍型,出现频次最高,分布于86 个样本中,占总样本数的57.33%,这可能是矛尾复虾虎鱼的原始单倍型。5 个群体的单倍型数量:连云港群体有8 个(Hap2、 Hap7、 Hap8、 Hap9、 Hap10、 Hap11、Hap12、Hap13),数量最多,大连群体有6 个(Hap1、Hap2、Hap3、Hap4、Hap5、Hap6),长江口 群 体 有6 个(Hap2、Hap7、Hap8、Hap9、Hap11、Hap14),宁波群体有5 个(Hap2、Hap8、Hap15、Hap16、Hap17),南通如东群体有3 个(Hap2、Hap7、Hap8),相对最少。利用Kimura 2-parameter model 构建17 个单倍型的NJ 系统发育树(图2),发现有一明显分支(由Hap1、Hap15 构成)且节点支持率不低。利用Network 10.2 软件分析单倍型之间的进化关系:5 个群体的单倍型网络分布图呈现一种以5 个群体的共享单倍型(Hap2)为中心的放射状结构;5 个群体中宁波部分群体单倍型和其他四个群体的单倍型基本处于分割状态,表明宁波的部分群体与其他群体存在较大的遗传距离,这与NJ 系统发育树的结果相一致(图3)。
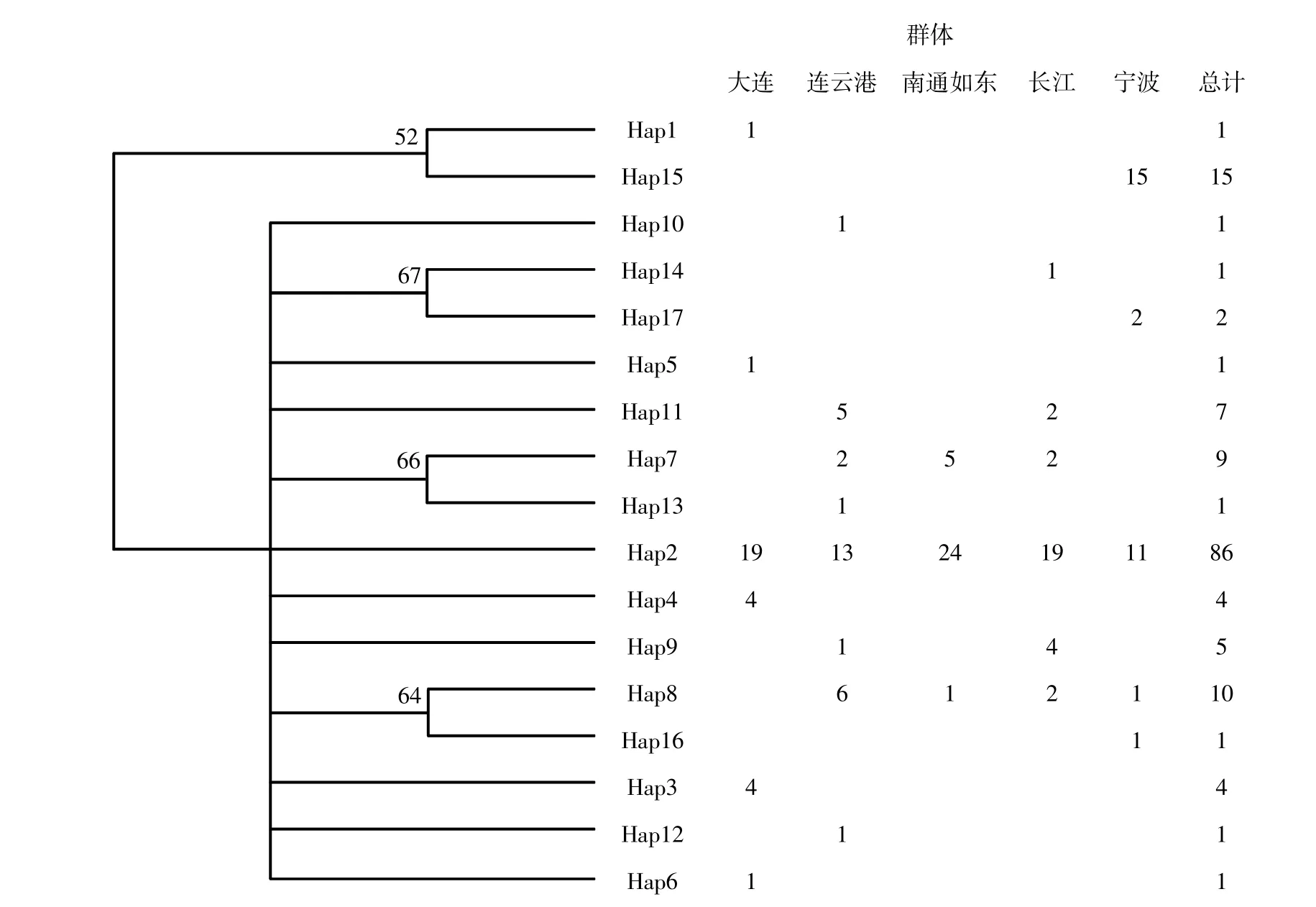
图2 矛尾复虾虎鱼17 种单倍型在5 个群体中的分布及其分子系统树
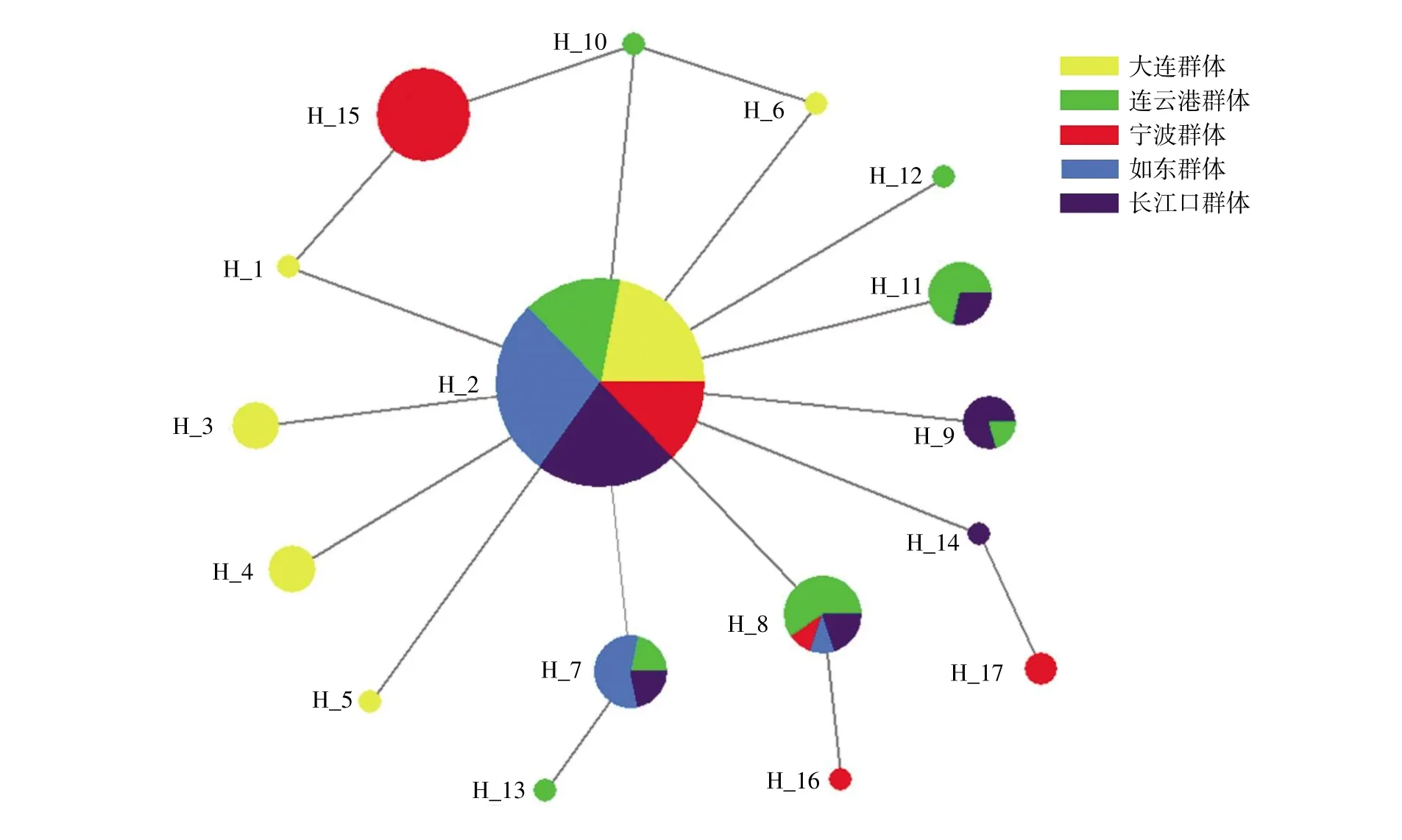
图3 矛尾复虾虎鱼单倍型网络关系图
2.3 群体多样性和遗传结构
各群体的单倍型多样性指数、核苷酸多样性指数等遗传参数指标如表2 所示。由表可知,各群体的单倍型多样性为0.343~0.761,各群体核苷酸多样性为0.000 83~0.003 48,连云港和宁波群体的单倍型多样性较高(0.761 和0.630),长江和大连群体其次(0.586 和0.579),如东群体的单倍型多样性较低(0.343);宁波群体的核苷酸多样性指数较高,连云港、大连和长江群体其次,如东群体的核苷酸多样性指数较低。以上结果表明:连云港和宁波群体具有较高的遗传多样性,其次是大连和长江群体,如东群体的遗传多样性较低。
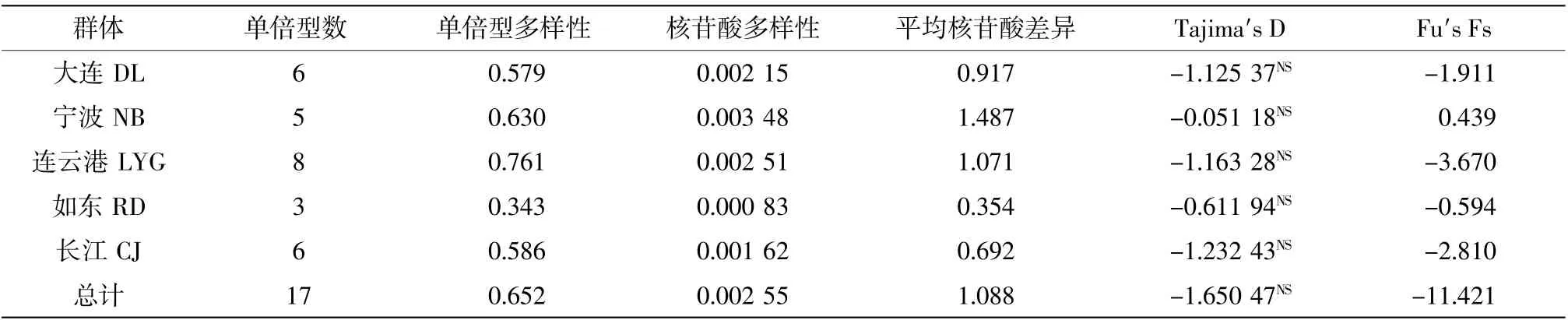
表2 各群体的单倍型多样性、核苷酸多样性和平均核苷酸差异
群体间FST 分析结果显示,两两群体间的FST值为0.015 57~0.355 66(表3),宁波群体与其他群体间的遗传分化系数FST 相对较大,为0.278 58~0.355 66,且存在显著差异(P<0.05)。5 个群体的遗传距离分析显示:群体内遗传距离为0.000 83~0.003 50,群体内遗传距离最大的为宁波群体(0.003 50),遗传距离最小的为如东群体(0.000 83);两两群体间的遗传距离为0.001 27~0.004 17,宁波群体与其他群体间遗传距离均相对较远(0.003 36~0.004 17),这与FST 分析结果相一致。AMOVA 分析结果表明来自群体内的遗传变异显著高于来自群体间的遗传变异百分比(表4)。
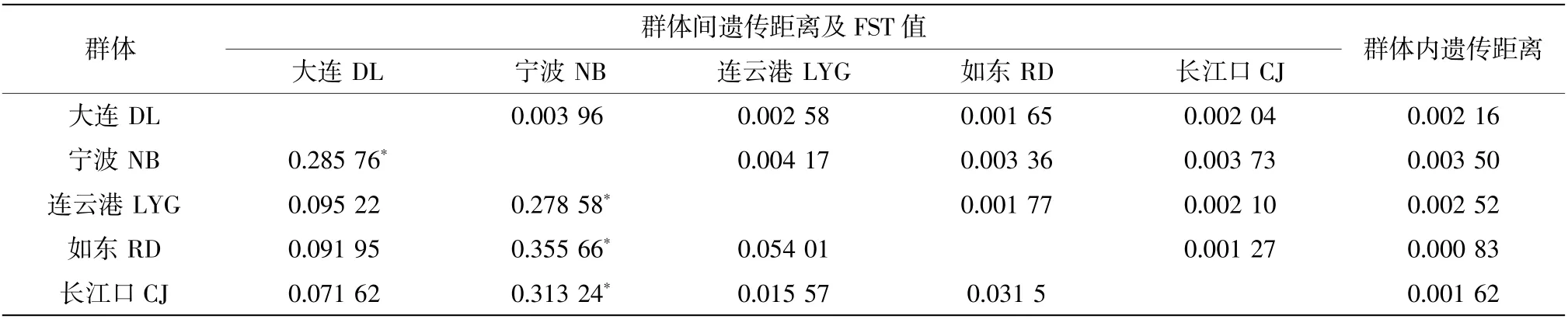
表3 矛尾复虾虎鱼群体内遗传距离及两两群体间遗传距离和遗传分化系数(FST)
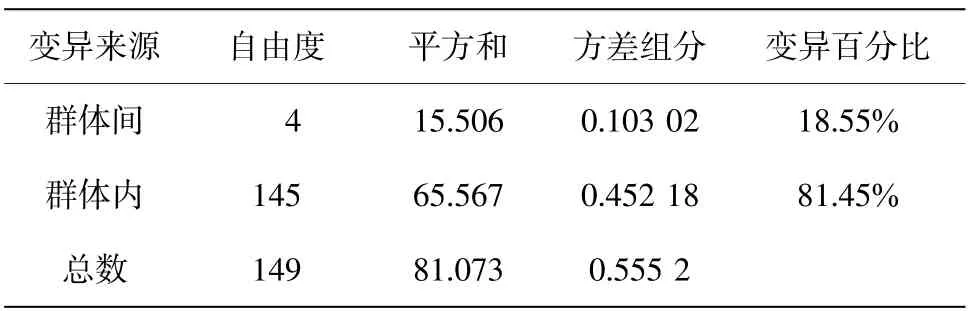
表4 矛尾复虾虎鱼群体遗传差异的AMOVA 分析
2.4 群体历史动态分析
表2 的中性检验结果显示,除宁波群体,其他群体的D 值和Fs 值均为负,表明检验的结果偏离了中性模式但不显著(P>0.1)。总体上说,矛尾复虾虎鱼群体历史上比较稳定,结合核苷酸不配对分析结果与中性检验结果大致吻合,即除了长江和连云港群体可能有过较小的群体扩张,其他群体均比较稳定。

图5 虾虎鱼核苷酸不配对分布
3 讨论
3.1 群体遗传多样性分析
单倍型多样性和核苷酸多样性作为线粒体标记,是用以衡量遗传多样性的重要指标,单倍型多样性和核苷酸多样性数值与物种的遗传多样性正相关(Jean-Pierre,2002)。Grant 等(1998)认为单倍型多样性数值高于0.5、核苷酸多样性大于0.005则表明物种的多样性越高,反之越低。本研究中,5 个矛尾复虾虎鱼群体单倍型多样性为0.343~0.761,各群体核苷酸多样性为0.000 83~0.003 48,将矛尾复虾虎鱼5 个群体作为整体,分析结果显示,5 个群体的矛尾复虾虎鱼单倍型多样性平均为0.652,核苷酸多样性平均为0.002 55,表明这5 个群体的遗传多样性处于中等水平,其中宁波群体的遗传多样性略高于其他4 个群体。宁波沿海属于东海海区,大连海域属于渤海海区,连云港、长江口和如东海域属于黄海海区,Song 等(2010)认为东海矛尾复虾虎鱼遗传多样性和遗传分化指数要高于渤海和黄海样本,本研究支持该观点。5 个群体中除了如东群体为低单倍型多样性和低核苷酸多样性类型,其他群体都是高单倍型多样性和低核苷酸多样性类型,核苷酸多样性由于累积时间远大于单倍型多样性,更能反映群体的遗传多样性(Lan et al,1993)。这5 个群体均呈现出核苷酸多样性偏低的情况,其主要原因可能是人类活动使群体扩散以及群体间基因交流受阻导致。如东群体核苷酸多样性结果显示:如东群体近期经历了瓶颈效应,而其他群体的低核苷酸多样性则可能是瓶颈效应后群体迅速扩张的结果。矛尾复虾虎鱼群体这种高单倍型多样性和低核苷酸多样性的现象,在东营、烟台(孙志成 等,2020) 和浙江沿海地区(顾翠平,2014)等地虾虎鱼研究中也出现过,且在其他鱼类中也有发现(李大命等,2020;程清清等,2017;曹洋铭等,2020)。研究者们一致认为群体近期可能经历了瓶颈效应,又经历了一个由较小的有效群体快速增长为大群体的过程,也表明了国家对渔业资源的保护政策初见成效。
3.2 虾虎鱼群体遗传分化分析
矛尾复虾虎鱼5 个群体中,宁波群体和其他4 个群体的遗传距离相对较大,但均小于0.01。根据Shaklee 等(1982)提出的鱼类在属、种和群体三级水平上遗传距离分别是0.9、0.30 和0.05 的分类依据,虾虎鱼的所有群体均未达到群体的分化标准。宁波群体和其他群体的遗传距离可能和海水中的环境屏障有关。随着鱼类分子系统地理学研究的深入,发现除了海洋中的物理障碍可以阻止鱼类交流,温度、海沟等环境屏障也有同样作用(Liu et al,2010;Han et al,2010)。宋娜等(2011)对丹东群体和天津群体之间的斑尾复虾虎鱼研究也得出了类似结果,即某种屏障阻碍了群体间的基因交流,可以导致群体遗传距离的差异。
遗传分化系数FST 是反映群体间遗传分化程度的重要指标(Allendorf,1983),FST 值越高,遗传分化程度越高。当FST 为0~0.05 时为无分化,0.05~0.15 为中度分化,0.15~0.25 为高度分化,超过0.25 时则表明存在极大的遗传分化。由表3 可知宁波群体与其他4 个群体之间的FST 均超过0.25,且P<0.05,为显著差异,说明宁波群体与其他4 个群体间有明显的遗传分化;其他群体间多为无分化或中度分化且P>0.05,差异不显著,这种现象主要原因与遗传距离差异类似。因宁波与其以北的长江口之间存在某种环境屏障,而长江口及其以北地区间不存在这种屏障且有洋流的输送作用,导致长江口群体和宁波群体距离上很近但遗传分化系数偏大、除宁波外的群体距离上不近但遗传分化系数偏小。除如东群体,其余4 个群体均存在特有的单倍型,这表明群体间存在一定的遗传分化,但总体而言矛尾复虾虎鱼群体间分化程度较低。根据丹东和天津斑尾刺虎鱼遗传多样性研究,结果显示丹东与天津群体相互混杂,没有群体特有的单倍型序列分支,群体分化程度也较低,与本研究结果相符(宋娜等,2011)。
3.3 系统进化分析
在单倍型的NJ 进化树中,17 个单倍型被分为2 个大分支,宁波的部分群体构成的Hap15 与只有单个个体的大连的Hap1 构成一个大分支,另一个大分支由5 个群体构成。根据遗传分化系数FST,宁波群体与其他群体间的遗传结构存在高度分化;宁波群体与其他4 个群体的遗传距离相对较远,群体遗传高度分化且主要的遗传变异来源于群体内,具有一定的地理差异。
4 结论
本研究通过对矛尾复虾虎鱼的5 个群体进行遗传多样性分析,发现只有宁波群体的遗传多样性相对较高,大连、长江口、连云港这3 个群体的遗传多样性较低,如东群体的遗传多样性最低,矛尾复虾虎鱼的群体资源现状并不乐观,过度捕捞可能是其遗传多样性偏低的主要原因。整体而言,我国矛尾复虾虎鱼野生种质资源现状堪忧,亟须开展相关的保护管理,鉴于长江口以北沿岸地区的群体未有显著遗传分化,可在江苏、山东等地开展人工育苗和增殖放流增加虾虎鱼资源量,修复和改善矛尾复虾虎鱼的栖息环境、严格控制捕捞强度,从而在不会产生负面遗传效应的前提下提高矛尾复虾虎鱼群体的遗传多样性。
