黄曲霉毒素的产生机制及污染防控策略
2021-02-02邢福国张晨曦
邢福国,李 旭,张晨曦
(中国农业科学院 农业农村部农产品质量安全收贮运管控重点实验室/农产品加工研究所,北京 100193)
黄曲霉(Aspergillusflavus)是一种广泛分布的腐生好氧真菌,属于子囊菌亚门(Ascomycotina)、曲霉属(Aspergillus),是仅次于烟曲霉(A.fumigatus)的曲霉病诱发菌[1]。黄曲霉可以寄生于花生、玉米、大米等主要粮食作物及坚果、香料、中药材等经济作物。根据联合国粮农组织统计,黄曲霉是全球谷物最主要的污染真菌之一[2]。我国是黄曲霉污染的重灾区,抽样调查显示我国多个省份的储藏玉米和花生中都检出了黄曲霉菌[3]。更重要的是黄曲霉侵染过程产生多种有毒次生代谢产物,如黄曲霉毒素(aflatoxins,AFs)、环匹阿尼酸(cyclopiazonic acid)、黄曲霉震颤素(aflaterm)等,其中黄曲霉毒素是毒性最高、产量最大、污染范围最广泛的真菌毒素[4]。真菌毒素的发生严重损害作物品质和经济价值,危及人畜健康和食品安全。
黄曲霉毒素是一类二氢呋喃香豆素衍生物,根据在紫外光照射下产生的荧光不同主要分为B族(蓝色,AFB1和AFB2)和G族(绿色,AFG1和AFG2)[5]。黄曲霉毒素因为其高毒性和高致癌性而广受关注,研究表明:长期暴露于黄曲霉毒素会导致人和牲畜的免疫抑制、营养代谢不良、不孕、先天畸形、内分泌紊乱、以及严重的肝细胞癌变[6]。同时,黄曲霉毒素也是迄今为止发现的理化性质最稳定的真菌毒素,一旦污染发生,很难通过物理或化学的手段去除,并会通过代谢和加工过程在食物或饲料中富集,严重威胁人畜健康[5-6]。
黄曲霉在农作物生长、收获、运输、贮藏、加工等不同阶段都可能侵染,而这些过程的不同光照、温度、水活度、宿主等环境差异均会影响黄曲霉毒素产量。研究不同环境因素对黄曲霉毒素合成的影响对黄曲霉毒素污染防控具有重要意义。本文概述了光照、温度、水活度、碳氮源、pH值、氧化胁迫对黄曲霉毒素合成的影响及调控途径,并总结相应的黄曲霉毒素污染防控措施,旨在为进一步解析黄曲霉毒素产生机制和更好的防控真菌毒素污染提供一定帮助。
1 黄曲霉毒素的生物合成
1.1 生物合成过程
黄曲霉毒素的生物合成过程非常复杂,但经过众多研究人员的长期研究,已基本解析清楚。黄曲霉毒素合成过程包括27个酶促反应,以乙酰辅酶A和丙二酰辅酶A为合成底物,在聚酮合酶AflC(polyketide synthase,PKS)的催化下形成聚酮骨架,再经过多步的加氧、脱氢、甲基转移等修饰过程,最终形成了高度氧化的黄曲霉毒素[7]。已知的黄曲霉的生物合成过程见图1。
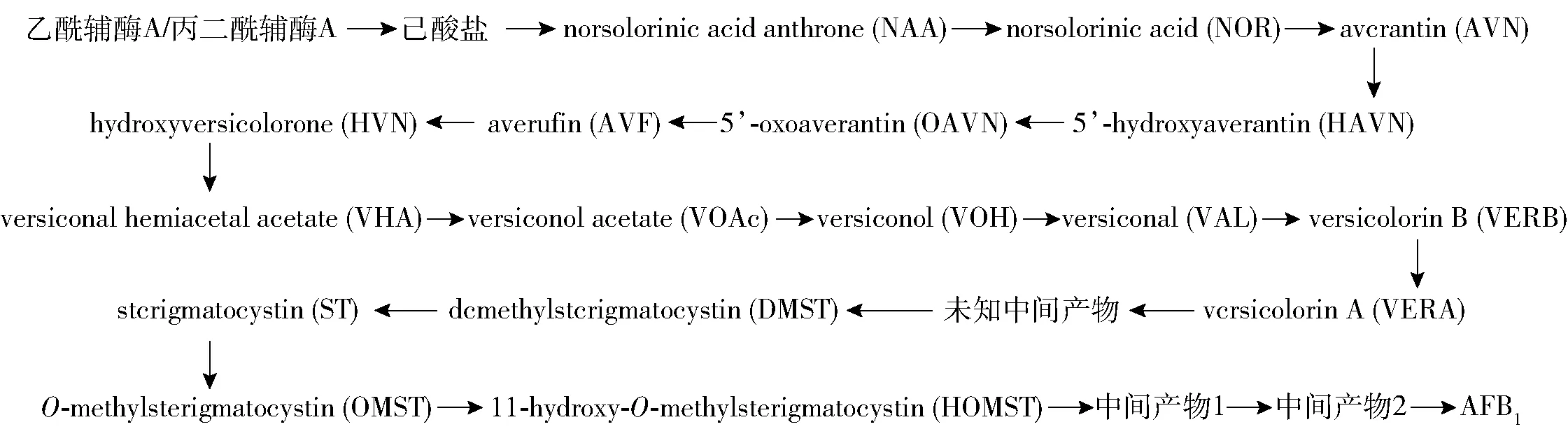
图1 黄曲霉毒素的生物合成过程
最新的研究发现,从HOMST到最终的黄曲霉毒素B1之间还有两个中间产物[8]。而根据基因簇功能推测,在VERA和DMST之间还有一个中间产物未被确认[7-8]。
1.2 合成基因簇
黄曲霉合成基因簇约80 kbp,位于3号染色体端粒上,为54号基因簇,包含30个基因,在黄曲霉和寄生曲霉中相对保守,同源性在90%~99%[7]。整个合成基因簇的绝大多数基因功能已基本解析清楚,hypC是最后被确认的基因簇结构基因,位于aflC和aflD之间,负责催化NAA到NOR的酶促反应[9]。
黄曲霉毒素合成结构基因的最直接调控元件是簇内转录调控因子AflR和转录激活子AflS。AflR对黄曲霉毒素合成过程是必须的,AflR至少能够与17个黄曲霉毒素基因簇结构基因的启动子区结合,回文序列5′-TCG(N5)CGA-3′是AflR最主要的识别序列,同时也能结合5′-TTAGGCCTAA-3′及5′-TCGCAGCCCGG-3′[10-11]。此外,AflR还能识别其自身启动子的5′-TTAGGCCTAA-3′区域,说明AflR可能激活自身转录表达,而黄曲霉毒素的合成可能存在着自反馈调控机制[12]。aflS是毒素基因簇内的转录激活子,aflS敲除突变体导致多个毒素合成基因的转录下调,而过量表达aflS促进了毒素合成基因的表达及AFB1合成[13-14]。aflS的转录水平同样被AflR调控。AflR和AflS以蛋白复合物发挥对合成基因簇的转录调控作用,4分子AflS搭配1分子AflR。当AflS表达量不足以形成复合物时,毒素合成基因簇结构基因的转录也会下调[13,15]。
此外,黄曲霉合成基因簇上还有两个基因功能并不完全清楚,aflT编码一个MFS家族真菌转运蛋白,蛋白定位在黄曲霉毒素小体内,因此推测AflT与毒素的外泌和运输相关,但aflT突变体并不影响黄曲霉毒素产量[16]。hypD编码一个膜结合蛋白,hypD敲除突变体的AFB1和孢子产量均显著下降,表明HypD参与真菌发育和毒素合成过程[17]。黄曲霉毒素合成过程和合成基因簇的研究为后续的调控机制解析及毒素抑制和降解工作奠定了坚实的理论基础。
2 环境因素对产黄曲霉毒素的影响
2.1 光照的影响
光照是最重要的环境信号之一,不同的波长、强度、光照时间都显著影响丝状真菌的生长、发育及次级代谢产物合成过程。早期光照相关的研究主要集中于光照对于真菌产孢和形态发育的影响,特别是对真菌有性生殖和无性生殖的调节。近几年,关于光照调控真菌次生代谢产物的相关研究也越来越多[18]。红光促进红曲霉中多种次生代谢产物的产量,而蓝光只能促进γ-米诺丁酸的产生[19];UV-A能够减少炭黑曲霉中赭曲霉毒素的产生,而625 nm红光和740 nm远红光抑制韦氏曲霉中赭曲霉毒素合成[20-21]。最新研究表明:455 nm和470 nm的蓝光能够显著抑制黄曲霉和寄生曲霉生长,并抑制黄曲霉毒素基因簇表达和黄曲霉毒素合成,而绿光、黄光和红光对黄曲霉毒素合成过程没有显著影响[22]。
在黄曲霉上,真菌可以感知近紫外光、蓝光、绿光、红光和远红光。曲霉属真菌中,已报道有11个光受体和多个信号级联系统参与到光信号调控过程[18]。这些光信号调控系统中,Velvet复合体是最核心的调控元件。VeA-LaeA-VelB形成三聚体复合物,与不同光受体蛋白,如红光受体FphA发生互作,感知不同光信号,进而进入核内调控相关发育和次生代谢相关基因的表达[23-25]。复合体核心调控因子是全局性调控因子VeA,影响黄曲霉生长、发育、多个次生代谢过程。对于黄曲霉毒素生物合成过程,VeA是必须的,黄曲霉和寄生曲霉的veA敲除突变体不产毒素,几乎完全抑制了aflR和aflS的表达,还影响了大多数的黄曲霉毒素合成基因簇基因表达[24-25]。除VeA之外,LaeA和VelB也正调控毒素合成和毒素基因簇转录[25]。这些研究均有力地证明了Velvet复合体是光信号调控黄曲霉毒素合成的关键。另外,红光和蓝光信号受体调控可能通过了HOG途径(high osmolarity glycerol pathway),多个MAPK激酶的表达受到不同光信号的影响,同时也表明了光信号和水活度、氧化应激等不同环境因子之间也可能存在着交叉的影响和调控。
2.2 温度的影响
温度是调控真菌生长和次生代谢产物形成的重要因素之一,也是调控黄曲霉毒素形成的关键环境条件,直接影响黄曲霉毒素产量。对大多数丝状真菌来说,最适的次生代谢产物产生温度略低于其最适生长温度[7,18]。黄曲霉和寄生曲霉在12~34 ℃产毒,而在36 ℃时基本停止产毒,最适产毒温度为20~30 ℃[5,18]。最适产毒温度也会根据水活度和培养基质等其他因素的差异而发生改变,Arrus等[26]以巴西坚果为培养基,水活度为0.97时,25 ℃达到最大的黄曲霉毒素产量;而在粳米上,33 ℃时AFB1的产量最高[27]。多个转录组分析和RT-qPCR实验结果表明,温度对黄曲霉合成基因簇基因的转录表达有着显著影响,黄曲霉毒素合成基因簇调控基因和大多数结构基因在相对低温的条件下被诱导表达。在30 ℃时,aflR和aflS表达水平分别比37 ℃高5倍和24倍,可见温度可能通过影响合成基因簇特异性调控因子AflR和AflS的转录水平进而影响结构基因的表达以及毒素产量[28]。此外,AflS是AflR的激活子,低温导致了更高的AflS/AflR比例,从而进一步促进了毒素合成结构基因的转录[15,28]。到目前为止,还没有发现其他转录因子或调控蛋白响应温度变化,也没有发现温度感应的相关受体,温度变化的信号如何传递到AflR和AflS也是未知的,温度如何调控黄曲霉毒素合成还需要进一步解析。
2.3 水活度的影响
水活度是食品或农作物中自由水的含量,微生物只能利用自由水进行代谢,水活度是影响食品微生物生长和次生代谢的关键环境因素。同样,不同水活度条件也影响黄曲霉毒素的生物合成。一般来说,较低的水活度抑制黄曲霉毒素产生,高水活度条件促进黄曲霉毒素产生[5,18,27]。大部分黄曲霉和寄生曲霉在水活度低于0.90时,生长就会受到抑制,黄曲霉毒素产量显著降低[29]。Lahouar等[30]从高粱上分离的一株黄曲霉在水活度低于0.94时就停止产毒,表明水活度对黄曲霉毒素的调控有着菌株特异性。此外,水活度也常常和温度、光照、基质、培养时间等环境条件相互作用,共同调节真菌毒素的产生[5,27]。玉米贮藏过程发现,贮藏5~7 d,在水活度0.98时适宜黄曲霉毒素产生;而贮藏10~12 d,水活度0.93~0.95时,黄曲霉毒素污染最严重[31]。花生上最适的产毒条件为0.98,而粳米在水活度0.96时黄曲霉毒素产量最高[27]。Gallo等[32]发现在杏仁培养基上,水活度0.96和28 ℃条件AFB1产量最高。进一步对相关基因的表达检测发现,在最适水活度条件下,黄曲霉合成基因簇基因表达水平最高,尤其是aflS和aflR的转录水平受水活度影响明显。Zhang等[33]发现在水活度0.99条件下黄曲霉毒素和孢子产量均明显高于水活度0.93,转录组数据显示随着水活度的提高,16个黄曲霉合成基因簇基因表达显著增强,参与渗透势调节的HOG途径关键调控因子SakA转录明显提高。笔者团队近期也发现,最适的水活度条件能够提高激酶SakA和转录因子AtfB的转录表达,推测水活度条件可能通过“激酶SakA-转录因子AtfB-AFs合成基因簇”的调控途径影响黄曲霉毒素合成,具体的调控机制还有待进一步实验确认。
在食品和粮食贮藏过程中,水活度是影响真菌毒素的关键因素,也是贮藏可控条件之一。解析水活度调节黄曲霉产毒的机制有助于建立抑制黄曲霉生长和产毒的综合防控体系,具有重要意义。
2.4 碳源与氮源的影响
碳源、氮源、脂质、氨基酸、微量元素等营养元素能够影响黄曲霉毒素的产生,其中碳源和氮源营养对黄曲霉毒素合成的影响最显著。
碳源代谢过程为黄曲霉毒素合成提供了基本碳单位,糖代谢的终产物(乙酰辅酶A)是黄曲霉毒素合成的最主要的起始单位,从底物供给水平上影响了毒素合成[7]。不同作物和食品中的碳源差异直接影响黄曲霉毒素污染程度。笔者团队以玉米、大米、花生为基质培养黄曲霉,转录组和蛋白组联合分析发现,不同作物基质中的碳源(淀粉含量)差异是影响不同基质产毒量的最主要原因[34]。进一步,Yu等[35]发现一个和蔗糖利用相关的基因簇与黄曲霉毒素合成基因簇紧密相连,基因簇的物理位置关系佐证了蔗糖对黄曲霉毒素合成的诱导。因此,对黄曲霉碳代谢的研究是解析黄曲霉毒素发生规律的重要组成内容。不同碳源类型影响毒素产量。简单糖,如葡萄糖、蔗糖、果糖、麦芽糖等有利于黄曲霉毒素的形成,而淀粉、山梨糖、乳糖、纤维素等复杂多糖抑制毒素合成[5, 7]。碳代谢阻遏途径(carbon catabolic repression,CCR)是调控黄曲霉适应不同碳源,选择利用最适碳源的关键系统[36-37]。CreA是CCR途径的核心转录因子,负责启动下游多个水解酶的转录,调控真菌的碳源利用过程[36-37]。CreA除了通过底物供给水平的调控影响黄曲霉毒素合成,还可以在转录水平调控毒素合成基因簇的表达[37]。creA敲除突变体在完全培养基上毒素产量显著下降,而没食子酸通过抑制CreA的表达影响毒素合成基因簇表达和毒素产量[37-38]。由此可见,CreA正调控黄曲霉合成基因簇的表达。对构巢曲霉杂色曲霉素合成基因簇上多个基因的启动子区分析,发现多个可与CreA结合的保守结构域5′-SYGGRG-3′[37-39],推测黄曲霉毒素合成基因簇也有类似的保守结合位点。根据上述信息,我们认为CreA是调控黄曲霉毒素合成的重要转录因子,是联接碳代谢和黄曲霉毒素合成的重要节点。此外,CCR途径包括多个调控蛋白,如激酶Snf1和SchA、磷酸酶Reg1、泛素化酶CreD、去泛素化酶CreB和CreC等,均可通过多个翻译后修饰过程影响CreA活性[36-37],并可能进一步通过CreA调控毒素合成过程。笔者团队初步研究也发现Snf1和Reg1正调控黄曲霉毒素合成和毒素基因簇转录表达。碳源代谢、CCR途径与黄曲霉毒素合成过程有着复杂的关联,而相关研究还较少,需要更多研究才能阐明碳代谢与毒素合成之间的复杂关系。

2.5 pH值的影响
pH值是影响真菌次生代谢过程的另一个重要环境因素,合成不同次生代谢产物均有其最适的pH值范围。一般来说,环境条件偏酸时,真菌次生代谢产物产量提高,在黄曲霉、寄生曲霉、构巢曲霉、赭曲霉、纯绿青霉上都有着类似的表型[5,18]。黄曲霉毒素合成的最适pH值在3.4~5.5之间[5]。Nancy等[46]发现,当调整培养基pH值等于4.0时,黄曲霉毒素是原培养基毒素产量的10倍;而将pH值从4.0提高到8.0,寄生曲霉和构巢曲霉的产毒量均显著下降。寄生曲霉中的研究还发现,pH值低于6.0时,B族黄曲霉毒素的产量会提高;而较高pH值条件则诱导G族黄曲霉毒素的生物合成[46-47]。PacC是曲霉属真菌响应pH值变化的主要转录调控因子,碱性条件下PacC表达水平提高[48-49]。在碱性条件下,PacC被水解加工后激活,转运到细胞核,与靶标基因的启动子区结合,碱性pH值相关基因被激活转录,同时抑制酸性pH值相关基因表达[49]。寄生曲霉中aflR启动子区发现PacC可能的结合位点,推测碱性条件下PacC可能作为转录的负调控因子结合于aflR启动子区,抑制aflR转录表达[50]。但在构巢曲霉上敲除pacC却导致其杂色曲霉素产量显著降低[51]。可见PacC对毒素的合成调控是一个复杂的过程,可能与其他调控因子共同作用完成。除了PacC,丝状真菌pH调控途径还发现其他6个调控蛋白,均有着能感应pH值信号的Pal结构域,这些调控因子功能和作用机制还未解析[49],需要更多相关研究才能完全解开pH值变化与黄曲霉毒素合成过程之间的关联。
2.6 氧化胁迫作用
环境条件的变化会改变真菌细胞的活性氧状态(reactive oxygen species,ROS)平衡,但过高的活性氧水平可能损伤细胞的蛋白、脂质和DNA。为了避免活性氧爆发带来的损害,真菌有着复杂的氧化胁迫抵御系统。多个转录因子在真菌遭受氧化胁迫时诱导表达,并激活下游相关酶促反应[52]。事实上,对黄曲霉产生菌来说,黄曲霉毒素和其中间产物(NOR、VerA、OMST等)都是被高度氧化的分子,黄曲霉毒素合成也被认为是黄曲霉降低活性氧水平、抵御氧化胁迫的一种有效方式[53]。此前研究已证明,培养过程加入双氧水,适当提高真菌活性氧水平,能促进黄曲霉毒素的产量提高;而施用抗氧化剂,例如肉桂醛、丁香酚、抗坏血酸等,都能够在一定程度抑制黄曲霉毒素合成[5,54-55]。
已有大量研究解析了氧化胁迫调控黄曲霉毒素合成的机制。研究表明:过氧化氢酶基因catA和超氧化物歧化酶基因mnsSOD与黄曲霉毒素合成基因簇基因aflA、aflM和aflP的启动子区均包含了4个氧化应激相关转录因子(AtfB、SrrA、AP-1和MsnA)的结合位点,推测氧化应激相关酶和毒素合成酶的表达共转录[53]。因此,可能的调控机制为:黄曲霉毒素产生菌受到氧化胁迫时,相关转录因子被激活,表达水平提高,从而促进下游氧化应激酶类的转录,同时也增强了黄曲霉毒素合成基因簇的转录表达和黄曲霉毒素的合成。AtfB是CREB/ATF家族蛋白,被认为是能同时调控抗氧化酶和毒素合成酶的最重要共调节子,能够与毒素合成基因簇上的多个CRE位点(5′TG/TACGTC/AA3′)结合,从而激活毒素合成基因转录[56-57]。体外的ChIP和EMSA实验也证明,AtfB确实与7个黄曲霉毒素合成基因的启动子区互作结合[57]。锌指类转录因子AtfA能够与AtfB互作,共同调控下游基因的转录,AtfA正调控合成基因簇[58]。有趣的是单独敲除一个氧化应激转录因子,往往导致毒素产量提高。如突变体Δap-1和突变体ΔmsnA的黄曲霉毒素B1产量均提高[56]。推测这些突变体对活性氧的敏感度提高,反而促进了黄曲霉毒素的合成。该结果也进一步佐证了黄曲霉毒素在黄曲霉产生菌中发挥的抗氧化作用。综合多个研究的结果表明,AtfA、AtfB、SrrA、AP-1和MsnA组成氧化胁迫响应调控网络,当黄曲霉产生菌受到氧化胁迫时,刺激信号通过多个转录因子组成的调控网络影响真菌的氧化应激响应过程和黄曲霉毒素合成过程。
压力激活的蛋白激酶途径和促分裂原激活的蛋白激酶途径(SAPK/MAPK)是传递氧化应激信号、激活上述转录因子的重要环节[56]。SAPK/MAPK信号途径在酵母、丝状真菌以及高等真核生物中都高度保守,由三个激酶、两步磷酸化过程组成[56]。磷酸信号从上游MAP3K和MAP2K依次传递到MAPK,并由MAPK磷酸化下游转录因子的丝氨酸/苏氨酸残基,进而激活下游靶标蛋白[56]。除了氧化应激,MAPK通路也参与渗透胁迫和温度响应过程[59-60]。此外,氧化应激转录因子不仅仅被MAPK途径影响,同时也被cAMP-PKA途径调控。而调控细胞cAMP水平的PkaA、PkaC、PkaR等因子可与MAPK激酶SakA和MpkC互作。因此,cAMP-PKA途径与MAPK途径关系密切[61]。碳代谢也与cAMP信号途径相耦联,同时碳代谢相关信号也会影响MAPK通路的磷酸信号传递[61]。可见,光照、水活度(渗透势)、温度、碳源等环境因子之间有着复杂的互相影响和交叉调控过程,MAPK磷酸信号传递途径是联结不同环境信号刺激的共同通路,而这种机制也部分的解释了不同的环境变化却导致同样表型的原因。
光照、温度、水活度、碳氮源、pH、氧化胁迫等环境条件会直接影响黄曲霉毒素合成基因簇表达,或通过相关调控因子间接影响毒素合成过程,见图2。环境因子对黄曲霉毒素合成的影响是一个复杂的过程,众多调控蛋白参与其中。现阶段的研究结果只能部分的解释调控过程,很多调控机制还未解析。对相关调控机制的解析有助于在食品或农作物储藏、加工过程中减少或抑制黄曲霉毒素发生,有的放矢的防控黄曲霉毒素污染。
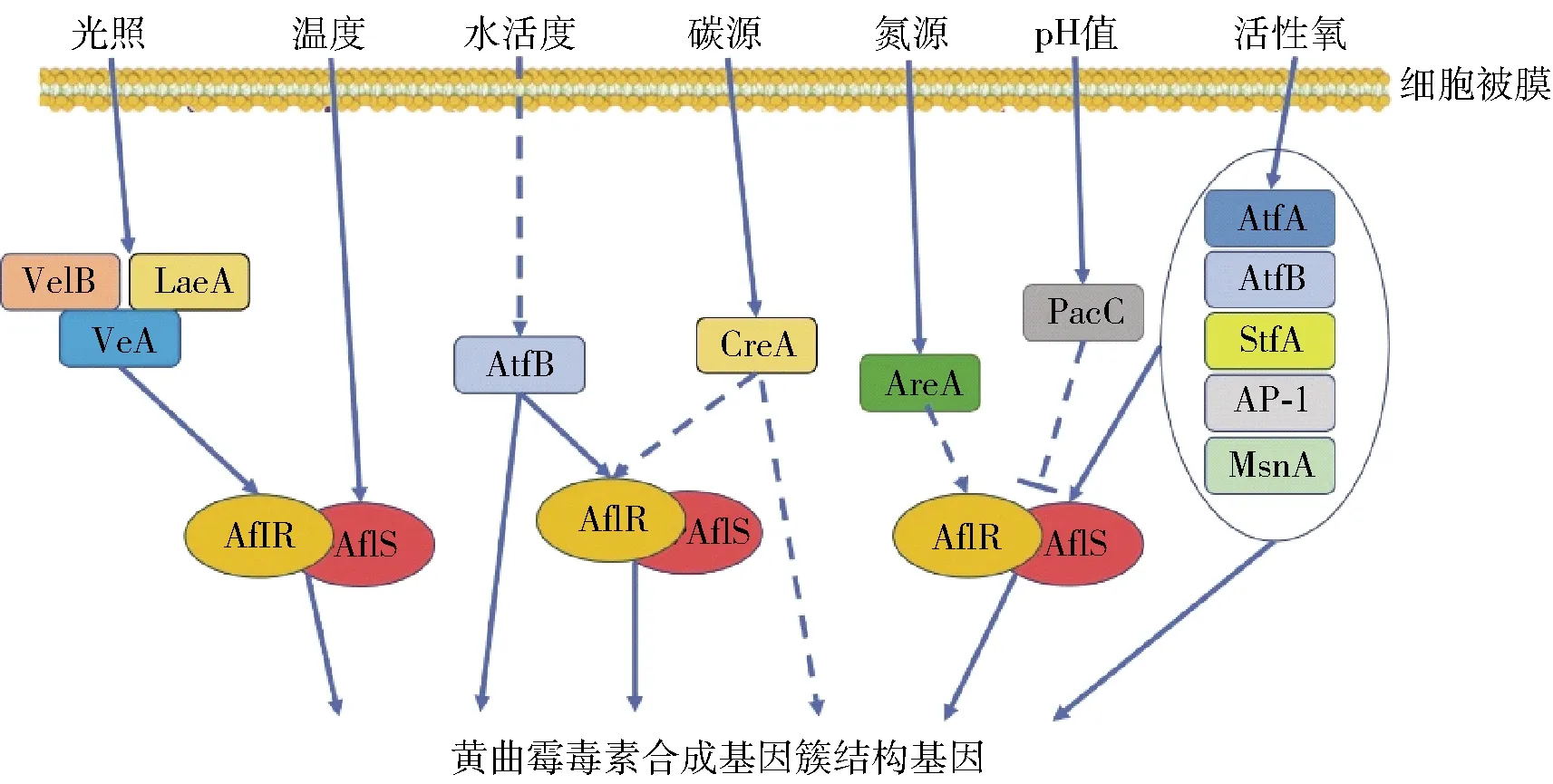
图2 环境因子对黄曲霉毒素的调控机制
3 黄曲霉菌和黄曲霉毒素的防控策略
黄曲霉防控技术包含2个方面的研究,即主动预防(黄曲霉生长控制)和被动脱除(黄曲霉毒素脱除)。防控方法主要有物理法(吸附剂、光、辐射等)、化学法(氨气熏蒸法、碱处理、臭氧降解、提取挥发油等)和生物法(微生物拮抗、微生物吸附和降解等)。
3.1 物理法
3.1.1吸附剂吸附
吸附剂主要利用其自身的孔状结构,将固相-气相、固相-液相等二相体系中的吸附物富集于吸附剂表面,使黄曲霉毒素与被污染的农产品分离[62]。目前,市面上黄曲霉毒素吸附剂主要包括硅铝酸盐类(膨润土、黏土、沸石、蒙脱石、水合硅铝酸盐等)、酵母细胞壁、活性炭、新型复合吸附剂等[63]。研究表明,蒙脱土对AFB1具有很强的吸附能力。蒙脱土对牛奶中的黄曲霉毒素(5 μg/kg)去除效率达到80%,而不会影响牛奶的营养[64]。Magnoli等[65]对阿根廷3种蒙脱石在肉鸡饲料中吸附AFB1的效果进行研究,发现饲料中添加蒙脱石能轻肉鸡的中毒症状。李新等[66]比较了沸石粉与“沸石粉+酵母细胞壁”两种吸附剂减轻肉鸡黄曲霉毒素中毒的效果,发现沸石粉和“沸石粉+酵母细胞壁”均能在较低AFB1污染水平(1 mg/kg)下有效缓解其负面影响,但沸石粉对2 mg/kg的AFB1的缓解作用强于酵母细胞壁。但有些吸附剂在吸附毒素的过程中也会吸附很多营养物质,并且有可能在动物体内代谢过程中解吸附,黄曲霉毒素重新暴露,对动物造成危害。因此物理吸附法应进一步优化并明确吸附剂的代谢过程,降低解吸附率,尽可能减少吸附剂对营养物质的吸附[67]。
3.1.2光调控
光调控技术可不同程度的抑制黄曲霉生长及其次级代谢产物合成,同时又具有操控性强、能耗低、辐射低、污染小等优点,因此具有重要应用潜力[68]。程翠利[67]研究了蓝、黄、绿、红光对黄曲霉及黄曲霉毒素的影响,发现蓝光、黄光处理可显著抑制黄曲霉菌落萌发和对还原糖与蛋白的吸收,其中蓝光可造成黄曲霉细胞壁、细胞膜及细胞器表面损伤,增加线粒体数目,造成细胞内环境紊乱。在毒素合成方面,蓝光组可使AFB1、AFB2、CPA、STR、AFG1、AFG2分别下降 96.33%、91.56%、92.27%、90.74%、61.78%、28.22%;黄光组分别下降 89.00%、75.58%、92.27%、90.74%、61.78%、45.41%;而红光和绿光却能促进黄曲霉生长及毒素合成[67]。
3.1.3辐照降解
电子束辐照技术能够产生低能或高能电子束射线,利用高能脉冲直接作用破坏活体生物细胞内DNA或通过间接作用使小分子和水发生辐解[69]。因此可以杀灭黄曲霉菌或降解黄曲霉毒素。王瑞琦等[70-71]和李苑等[72]采用240 kGy的电子束处理花生粕,AFB1的降解率达57.49%(初始量为120.24 μg/kg),同时花生粕中的粗蛋白、粗脂肪、氨基酸总量、总糖和灰分含量变化不大,还原糖、总黄酮含量还有所增加。辐照设备简单,价格低廉,但是能量损耗巨大,高剂量的辐照也可能对操作人员带来潜在的身体伤害,因此并未被广泛应用[69]。
3.2 化学法
3.2.1传统化学降解
黄曲霉在强碱性条件下能迅速分解,破坏其内酯结构,进而转化为香豆素钠盐等[73]。赵国斌等[74]用氨气熏蒸法降解黄曲霉,结果显示该方法效果显著,最高降解率达95.06%。李娟[75]使用强氧化剂次氯酸钠能将花生中的AFB1含量降低至零,但该方法对花生品质有较大影响。陈冉等[76]用臭氧处理被AFB1污染过的花生,脱毒效果显著,但也会降低花生品质。传统的化学脱毒效果显著,但因毒性强,污染大,会破坏营养品质,对环境造成不可逆影响等缺点,所以逐渐被淘汰。
3.2.2植物精油抑制
植物精油是由一系列挥发性有机物如酚类、萜类、醛酮类、醇类、酸类及芳香族化合物组成的混合物,是一类油状具有挥发性的芳香味物质。主要来源于高等植物的叶、枝、木浆或树皮组织,也广泛见于苔藓类植物[77]。很多植物精油具有较高的生物安全性,被美国食品药品监督管理局规定为“一般认为安全”(generally recognized as safe,GRAS)[78]。因植物精油在抗真菌方面具有高效,绿色、环保的优势而受到人们的广泛关注。
植物精油可有效抑制黄曲霉的生长和黄曲霉毒素的合成,其主要原因可能是多种植物精油能够破坏黄曲霉细胞内发生一系列的氧化反应,使菌体通过应激反应产生过氧化物,使细胞膜上的脂质发生过氧化,细胞膜受损,最终使得黄曲霉细胞死亡[79]。也有报道称,植物精油含有的酚类及萜烯类化合物可以有效破坏黄曲霉的遗传物质,使线粒体的内部结构发生变化,对黄曲霉的线粒体的有氧呼吸功能产生严重影响[80]。另外,植物精油也可以通过抑制黄曲霉菌丝及分生孢子的产生和萌发,抑制黄曲霉的繁殖与生长和发育[81]。有报道称,胡芦巴油可有效抑制黄曲霉的生长,最小抑菌浓度为0.87 μg/mL,当浓度为1 000 μg/mL时,对黄曲霉菌的抑制率可达100%[82]。分别将60 mg的肉桂精油、山苍子精油、丁香精油处理20 g黄曲霉污染的玉米,密封熏蒸30 d后,玉米中AFB1脱毒率分别为81.86%,77.76%,68.25%,对黄曲霉菌抑制产毒具有良好的效果[83]。孜然精油使黄曲霉生长降低81.9%,AFB1合成量下降83.01%,胞内总过氧化物含量减少33.75%,而且黄曲霉的菌丝和孢子都遭到了破坏[84]。罗勒精油也被报道可有效抑制黄曲霉的生长,当罗勒精油的浓度在1 500~2 000 μg/kg时,稻谷中的黄曲霉生长完全受到抑制[85]。
目前植物精油的研究已经不仅仅局限于植物精油本身,而是将其通过特殊处理如做成纳米乳剂,固定化等方式提高其抑制黄曲霉生长以及毒素合成的活性,以达到更好的商业和工业应用的目的。例如,杉木精油也具有抗真菌活性,被壳聚糖包埋后的杉木精油进一步提高了其抑制黄曲霉的效果[86]。
3.3 生物法
3.3.1微生物拮抗
对黄曲霉菌及毒素的防控研究中,报道的具有拮抗作用的菌株极多。但由于应用场景主要为粮食和饲料,因此我们主要关注生物安全性相对高的菌株,如乳酸菌、枯草芽孢杆菌、解淀粉芽孢杆菌、木霉菌等。Zhao等[87]筛选到一株植物乳杆菌(Lactobacillusplantarum)IAMU80070可有效抑制黄曲霉的生长,发现植物乳杆菌分泌的几丁质酶破坏了黄曲霉菌丝和孢子的细胞结构,抑制了黄曲霉生长及产毒。金黎明等[88]采用单因素试验和响应面试验对枯草芽孢杆菌的发酵条件进行了优化,成功将其对黄曲霉的抑菌能力提高了1.9倍。张学雯等[89]鉴定出解淀粉芽孢杆菌(Bacillusamyloliquefaciens)中具有抑菌活性的脂肽类抗生素分别为C14和C15的bacillocnycin。徐杨玉等[90]研究发现绿色木霉(Trichodermaviride)的发酵液不仅能够抑制黄曲霉生长还具有降解AFB1的作用。
3.3.2细胞壁吸附
多种微生物对黄曲霉毒素具有吸附作用。目前多采用乳酸菌、酵母菌、糙皮侧耳等益生菌菌体对AFB1进行处理,形成复合物,从而减轻AFB1对人和动物造成的危害[91-92]。这类益生菌的高效、对人畜无害、对环境友好的特点,在黄曲霉毒素污染防控方面受到了人们的广泛关注。菌体能吸附AFB1的主要原因可能是细胞壁上肽聚糖和多糖等成分通过形成非共价键与AFB1结合[91-92]。酿酒酵母(Saccharomycescerevisiae)在PBS小肠缓冲液中的菌浓度达到1010CFU/mL时,可在5 min内吸附99.3%的AFB1[93]。开菲尔乳杆菌(Lactobacilluskefir)在牛奶中繁殖到8.4×107CFU/mL时,24 h可以吸附82%的AFB1(1 μg/mL)[94]。杏鲍菇(Pleurotuseryngii)也被报道有吸附AFB1的能力。在28 ℃,pH值分别为3.7和7.4条件下,48 h杏鲍菇可完全吸附AFB1[95]。热力学研究表明:这是一个自发的物理吸附过程,杏鲍菇菌丝对AFB1的去除率最高可达85.13%, 推测真菌细胞壁上的羧酸或磷酸盐配体以及羟基和酰胺功能基团与AFB1分子形成了相对弱的非共价键[95]。
3.3.3微生物降解
在分离得到黄曲霉毒素降解微生物中,各种毒素降解酶起到了关键作用。目前分离纯化出的酶包括锰过氧化物酶[96],F420依赖型还原酶[97],水解酶[98],还原酶[99]和细胞色素P450酶[100],有关降解途径见图3。漆酶(Lac)和锰过氧化物酶(MnP)能够高效降解AFB1,相关研究最多,如杏鲍菇粗提物中含有高活性漆酶(4 U/g)和低活性锰过氧化物酶(0.4 U/g),在pH值为8时,反应3 d可以降解90% AFB1,反映出极好的应用潜力[101]。在最新的研究中,白腐真菌色齿毛菌(Cerrenaunicolor)6884中分离纯化出一种漆酶,在pH 7.0,45 ℃下与5 μg/mL AFB1反应24 h,可全部降解AFB1,降解产物鉴定为AFQ[102];寡养单胞菌属(Stenotrophomonas)菌株CW117在24 h内可全部降解45 μg/mL的AFB1,降解活性成分位于培养上清液,且降解活性随着温度的升高而升高,降解活性在90 ℃时仍保持稳定,研究发现是漆酶和小分子氧化物共同对AFB1发挥降解作用,该菌株有着较大应用前景[103];缓慢葡萄球菌(Staphylococcuslentus)被报道可以抑制黄曲霉生长并降解AFB1,分别在高粱和花生中添加10 mL/kg和25 mL/kg的菌液,黄曲霉的生长受到完全抑制,而液体培养的缓慢葡萄球菌可以降解96.54%的AFB1,但并未检测到特定降解产物[104]。
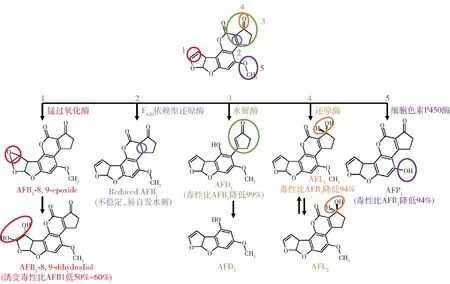
图3 黄曲霉毒素降解途径
4 结论与展望
黄曲霉广泛侵染玉米、花生等农产品,侵染后还会产生强毒性、高致癌性的黄曲霉毒素等真菌毒素,严重威胁了食品安全和人畜健康。因此,摸清黄曲霉及其毒素的发生发展规律,开发绿色、安全、高效的真菌毒素防控技术和产品尤为重要。
黄曲霉毒素的产生受到宿主、光照、温度、水活度等环境因素影响,当环境无法达到真菌的产毒条件时,可能会降低真菌的产毒量甚至不产毒。因此,在不影响农产品生产及储藏的情况下寻找一种最佳的防控真菌产毒的条件是农业生产中追求的目标,如:光调控、辐照等。随着生物技术的发展,利用微生物组学发掘、分子生物学调控分析、遗传学改造等多种生物技术进行黄曲霉及其毒素的生物防控已成为研究重点。且生物防治方法对农产品的营养物质损失较小,对环境污染少,防控特异性高,是具有较高防控效果和应用前景的黄曲霉毒素防控策略。然而,目前国内外黄曲霉及其毒素的生物防控技术还处于前期基础研究阶段,在农业生产中的产业化应用才刚刚起步。利用微生物防控黄曲霉、生物酶降解黄曲霉毒素的关键在于发掘出能分泌高活性抑菌物质或解毒酶的菌株,并在实际应用中构建出一个有利于拮抗菌生长和脱毒酶反应的环境。因此,通过各种生物技术来获得更优质和适用范围更广的拮抗菌和解毒酶是今后研究的重点方向。
