家蚕非滞育红卵突变体Re-nd的遗传分析
2021-01-12张宇靖张海燕沈关望吴金鑫贾林帮毛雪芹焦梦瑶
张宇靖, 张海燕, 沈关望, 吴金鑫, 贾林帮,毛雪芹, 焦梦瑶, 林 英,4,5,*
(1. 西南大学, 家蚕基因组生物学国家重点实验室, 重庆 400716; 2. 西南大学生物学研究中心, 重庆 400716;3. 四川省农业科学院植物保护研究所, 成都 610066; 4. 重庆市蚕桑学重点实验室, 重庆 400716;5. 重庆市蚕丝纤维新材料工程技术研究中心, 重庆400716)
家蚕Bombyxmori是作为遗传背景研究较为深入的鳞翅目模式昆虫,具有重要经济价值和基础科学研究价值,其形态性状一直备受关注。家蚕卵色是家蚕形态学的重要组成部分,在家蚕遗传学研究中占有重要地位,主要由卵壳色和浆液膜色组成。野生型家蚕卵的颜色在产卵初期呈现淡黄色,主要是由卵黄和黄色色素3-羟基-犬尿氨酸(3-hydroxy-kynurenine)透过卵壳所呈现出的颜色,不同品种、来源以及滞育性的卵颜色深浅稍有不同。随着卵的发育,3-羟基-犬尿氨酸经过酶促反应生成眼色素(ommochromes),在浆液膜上形成褐色色素,随着浆液膜内的色素颗粒不断沉积,浆液膜色也逐渐由淡褐色加深为深褐色,最终与卵壳一起形成卵的固有色(向仲怀, 2005)。
家蚕卵色的形成主要和家蚕体内色氨酸等氨基酸代谢及眼色素的形成有关,而积累在家蚕卵浆液膜内色素颗粒上的各种眼色素就来源于氨基酸代谢的产物,包括奥敏类(ommins)和奥马丁类(ommatins)以及它们的前体物质(Koga and Osanai, 1967; Linzenetal., 1974; Sawadaetal., 2000; Bannoetal., 2010)。经典的奥马丁类包括眼黄素(xanthommatin)、奥马丁D(dihydro-xanthommatin sulfate, ommatin D)以及视红质(dihydro-xanthommatin-O-β-d-glucoside, rhodommatin)(Linzen, 1974)。因此,研究眼色素的生物合成代谢对研究家蚕卵的着色机制具有重要作用。
家蚕突变品系中存在多种卵色突变品系(向仲怀, 2005),为研究家蚕卵色遗传学提供了很好的材料。目前,已通过定位克隆、RNA双链干涉、转基因修复等方法确定了卵色相关的突变体有w-1,w-2,w-3,w-3(oe),pe,re等(Quanetal., 2002a, 2002b, 2007; Komotoetal., 2009; Tatematsuetal., 2011; Osanai-Futahashietal., 2012, 2016),这些已确定突变基因的家蚕卵色突变体均与眼色素合成通路相关。家蚕白卵突变体w-1的突变基因与果蝇的cinnabar基因同源,由于缺乏犬尿氨酸-3-单加氧酶(kynurenine 3-monooxygenase, KMO)导致3-羟基-犬尿氨酸的合成受阻,不能形成眼色素从而导致卵的颜色偏白(Quanetal., 2002a, 2007)。白色卵突变体w-2和w-3是由于white基因的突变引起ABC转运蛋白(ATP-binding cassette transporter)功能缺失,使眼色素前体3-羟基-犬尿氨酸不能转运到色素颗粒进一步形成眼色素,导致w-2和w-3突变体的蚕卵因缺乏眼色素而呈现淡黄色(Quanetal., 2002b; Komotoetal., 2009; Tatematsuetal., 2011)。家蚕桃红眼白卵突变体pe是由于cardinal基因的突变而引起cardinal编码的吩噁嗪酮合成酶(phenoxazinone synthetase, PHS)的功能缺失,吩噁嗪酮合成酶是眼色素中奥敏类色素合成所必需的酶,同时也是催化3-羟基-犬尿氨酸形成眼黄素的合成酶,其功能缺失导致pe突变体的蚕卵缺乏奥敏类色素,而3-羟基-犬尿氨酸能发生自身氧化形成少量的眼黄素而呈现淡黄偏微红色(Osanai-Futahashietal., 2016)。红卵突变体re是Bm-re基因的突变导致所编码的MFS转运体蛋白(major facilitator superfamily transporter)的功能缺失引起。在眼色素生物合成通路中,MFS转运体蛋白可介导转运其下游奥敏类色素合成所必需的底物半胱氨酸或甲硫氨酸等含硫氨基酸进入色素颗粒,如果MFS转运体蛋白功能缺失将导致re突变体的卵缺乏奥敏类色素,仅含有奥马丁类色素,最终呈现出红色(Toyamaetal., 1913; Tirellietal., 1946; Kawase and Aruga, 1954; Sanai-Futahashietal., 2012)。目前关于眼色素生物合成后期3-羟基-犬尿氨酸进一步代谢合成奥马丁类的过程尚不完全清楚。
家蚕非滞育红卵突变体Re-nd是由本实验室林英等(2007)利用化学诱变剂N-甲基-N-亚硝基脲(N-methyl-N-nitrosourea, MNU)处理家蚕品种C108而诱导突变产生的新型非滞育家蚕卵色突变体,属于浆液膜色突变。该突变体在滞育的情况下与正常卵色一样是褐色,只有在非滞育的情况下才呈现出艳丽的红色。在家蚕中发现的滞育情况下呈现红卵表型的突变体也有很多(向仲怀, 2005; 范晓东等, 2006),但在非滞育的情况下具有艳丽的红色表型的突变体,目前就只报道了非滞育红卵突变体Re-nd。因此,对Re-nd突变体的研究,将有助于阐释和完善家蚕眼色素代谢通路,也为将来应用色素相关基因创制新品种提供分子标记和突变体表型生物标记。为此,鉴于家蚕非滞育红卵突变体Re-nd的遗传特性尚不明确,本研究将通过杂交及遗传分析,探明Re-nd突变体的遗传特征,为后续的基因克隆和功能应用研究奠定基础。
1 材料与方法
1.1 试虫来源
供试家蚕分别为野生型滞育二化型家蚕品种大造(Dazao)和非滞育多化型品种N4,家蚕突变体品系滞育白卵突变体w-2、桃红眼白卵突变体pe和红卵突变体re以及非滞育红卵突变体Re-nd(林英等, 2007),均由西南大学家蚕基因组生物学国家重点实验室保存。供试家蚕材料孵化幼虫的催青条件为: 温度25℃、光周期12L∶12D,催青周期为10 d左右。所有家蚕供试材料均在25℃、自然光的条件下以新鲜桑叶饲养。
1.2 试虫纯度检测
大造、N4、Re-nd、w-2、pe和re共6种供试家蚕材料均已是不发生遗传表型性状分离的纯系。以单蛾区饲养的方式进行饲养,即对同一蛾圈蚕卵孵化的幼虫(同胞后代)进行饲养,至成虫期(羽化出蛾时期)进行交配制种(同胞交配),每一交配后代产生的蚕卵集中附着于产卵纸上,形成一个单一蛾圈中的蚕卵。在产卵第5天调查同一蛾圈内的蚕卵卵色的分离情况,在此时期蚕卵的卵色表现为野生型蚕卵卵色或突变体蚕卵卵色的稳定表型(即固有色表型);进入滞育的蚕卵胚胎停止发育,而非滞育的蚕卵胚胎继续发育至产卵后大约10 d孵化出幼虫,由此对6种供试家蚕材料的遗传表型进行纯度鉴定。
1.3 杂交实验
选取各供试品种/突变体的纯系(未发生卵色分离、品系纯度合格)的单一蛾圈的蚕卵进行催青,孵化幼虫,单蛾区饲养,分别以Re-nd为母本和父本与其他卵色品系的亲本进行杂交,调查正交和反交所产生的F1代蚕卵的卵色表型。
选取亲本杂交产生的F1代进行催青,孵化幼虫,单蛾区饲养,以同蛾区内的F1代进行同胞交配,调查同胞交配产生的F2代蚕卵的卵色表型。
选取亲本杂交产生的F1代进行催青,孵化幼虫,单蛾区饲养,分别以F1代为母本和父本与Re-nd的亲本进行回交,并调查回交所产生的B1代蚕卵的卵色表型。
杂交、同胞交配及回交的每一代制2张蚕种(28个成对交配蚕蛾所产生的28个蛾圈蚕卵/张蚕种),用于卵色调查。
2 结果
2.1 Re-nd突变体与野生型大造和N4的杂交和遗传分析
非滞育的Re-nd分别与野生型滞育的二化品种大造和非滞育多化品种N4杂交,无论正交还是反交,其F1代的非滞育卵均表现为淡红色,排除Re-nd突变体是母性遗传或假母性遗传的可能(图1: A, B)。F1代同胞交配产生的F2代的非滞育卵出现卵色分离,有红色、淡红色和淡黄色,且每个蛾圈中的红色卵(包括淡红色卵)与淡黄色卵的数量比基本符合3∶1(图1: A, B及局部放大示意图);以F1代为母本和父本,分别与大造和N4回交,产生的B1代的非滞育卵均表现出卵色分离,有红色和淡黄色,且每个蛾圈中的红色卵与淡黄色卵的数量比基本符合1∶1(图1: A, B及局部放大示意图)。上述杂交后代F2代(即F1代同胞交配后代、回交后代)的性状分离符合孟德尔遗传定律,证实了Re-nd突变体属于独立的显性遗传,其非滞育卵纯合体为艳丽的红色,杂合体为淡红色。
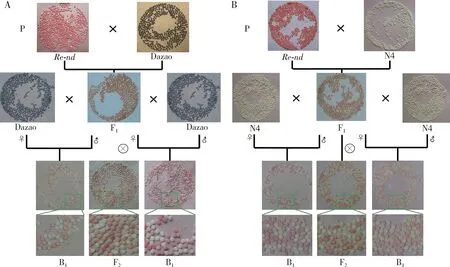
图1 家蚕突变体Re-nd与野生型品种大造(A)和N4(B)的杂交
2.2 Re-nd突变体与白卵突变体w-2的杂交和遗传分析
非滞育Re-nd与滞育白卵突变体w-2杂交,无论正交还是反交,其F1代均表现为淡褐色滞育卵色;F1代同胞交配产生的F2代出现卵色分离,滞育卵有正常深褐色、淡褐色、黄色以及淡黄偏微红色(图2: A),而Re-nd的滞育卵纯合体为深褐色、杂合体为淡褐色,w-2滞育卵为淡黄色。F2代卵色分离出现淡黄偏微红色卵,类似于pe突变体的滞育卵色;其F2代非滞育卵有红色、淡黄偏微红色、橙黄色以及淡黄色(图2: A),而w-2非滞育卵的颜色为淡黄色(Zhangetal., 2017),Re-nd的非滞育卵纯合体为红色、杂合体为淡红色,卵色分离出现橙黄色(图2: A)。推测Re-nd的突变基因与w-2的突变基因存在基因互作[在分子遗传学上理解,此经典遗传学上的“基因互作”关系,可能表现为眼色素代谢途径中上下游代谢底物的基因/酶的上下游关系(Tatematsuetal., 2011; Osanai-Futahashietal., 2012, 2016)],二者互作在一定程度上补偿了w-2突变体的缺陷,使色素代谢得以往下游底物进行,从而呈现更下游的色素代谢底物的颜色。暗示Re-nd的突变基因位于w-2突变基因的下游(范晓东等, 2010; Tatematsuetal., 2011),即3-羟基-犬尿氨酸进入色素颗粒之后的眼色素合成过程(图2: B)。
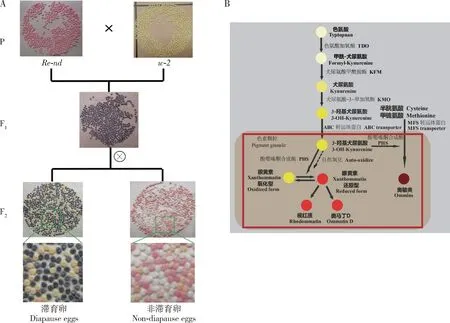
图2 家蚕突变体Re-nd与w-2的杂交及突变基因关系
2.3 Re-nd突变体与桃红眼白卵突变体pe的杂交和遗传分析
非滞育Re-nd与滞育桃红眼白卵突变体pe杂交,无论正交还是反交,其杂交F1代均表现为淡褐色滞育卵,F1代同胞交配后产的F2代全部是滞育卵,出现卵色分离,有正常深褐色、淡褐色、淡黄偏微红色以及红色(图3),pe滞育卵的颜色为淡黄偏微红色,而F2代滞育卵中卵色分离出现红色,卵色与re突变体的滞育卵色一致(图3: A及局部放大示意图)。推测Re-nd的突变基因与pe的突变基因存在基因互作,Re-nd突变体在一定程度上补偿了pe突变体的缺陷,眼色素合成得以往下游代谢底物进行,从而呈现更下游的代谢底物的色素颜色。暗示Re-nd的突变基因处于pe突变基因的下游(范晓东等, 2010; Osanai-Futahashietal., 2016),即参与3-羟基犬尿氨酸进一步代谢合成奥马丁类和奥敏类色素的过程(图3: B)。
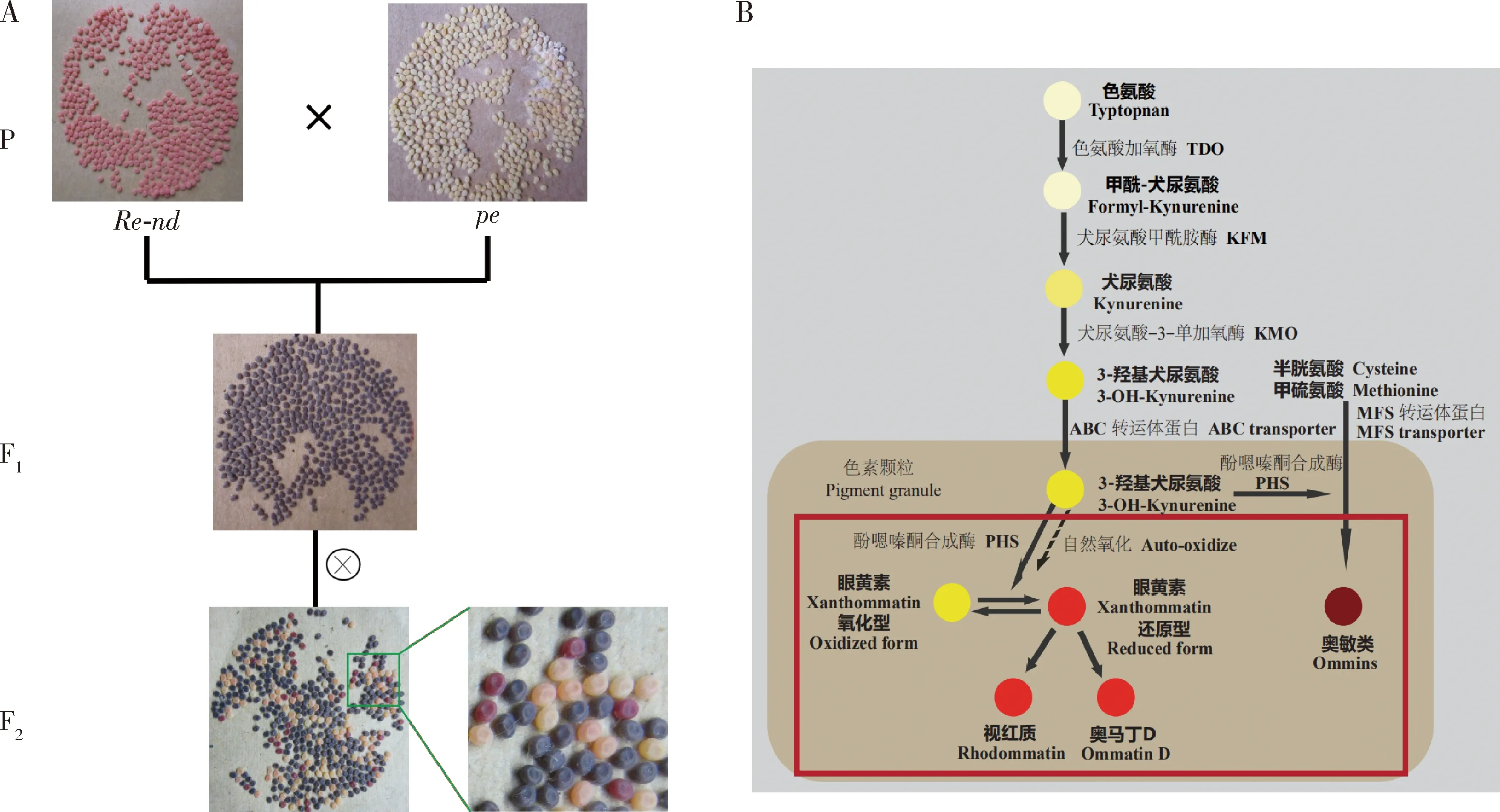
图3 家蚕突变体Re-nd与pe的杂交(A)及突变基因关系(B)
2.4 Re-nd突变体与红卵突变体re的杂交和遗传分析
非滞育Re-nd突变体与滞育红卵突变体re进行杂交,无论正交还是反交,杂交F1代均为淡褐色滞育卵色,F1代同胞交配后产的F2,以及与re回交产的B1代也全部是滞育卵,出现卵色分离,F2代有正常深褐色、淡褐色以及红色,B1代有正常深褐色和红色,而re滞育卵的颜色为红色,并没有分离出新的卵色(图4: A及局部放大示意图)。推测Re-nd的突变基因与re的突变基因同时存在的情况下,对蚕卵稳定着色没有改变,表明其色素合成途径出现了类似问题。而Re-nd突变体在滞育的情况下,卵色为正常的褐色,表明奥敏类色素的合成是正常的,暗示Re-nd突变基因与re突变基因参与了不同的色素代谢分支(Osanai-Futahashietal., 2012),即Re-nd突变基因可能参与了3-羟基犬尿氨酸进一步代谢合成奥马丁类色素的过程(图4: B)。
3 讨论

图4 家蚕突变体Re-nd与re的杂交(A)及突变基因关系(B)
眼色素是昆虫色素的主要成分之一,通常以颗粒的形式存在,主要表现为黄色、红色、棕色或紫褐色,多与昆虫的复眼、卵、表皮和翅的着色相关。由于眼色素及其前体物质具有易于辨认的颜色,眼色素合成相关基因常被用作分子标记(Klemenzetal., 1987; Fridell and Searles, 1991),在家蚕中,白色卵突变体w-1的突变基因Bm-Kmo可用于转基因的筛选标记,但由于其突变卵色为淡黄色,与野生型非滞育卵的卵色及未受精卵的卵色相近,不易区分;且w-1属于假母性遗传,作为分子标记应用受限;桃红眼白卵突变体pe以及深红眼红卵突变体re的突变基因也可以用作分子标记,然而这些卵色突变体均为隐性突变,作为分子标记不利于杂合子的筛选。非滞育红卵突变体Re-nd通过十几代的单蛾圈同胞交配筛选,最后获得了一个稳定遗传的突变体纯系。在对突变体与野生型大造和N4的杂交分析中,也观察到滞育卵产生,卵色都表现为褐色和淡褐色卵。
本研究关注的Re-nd是在非滞育的情况下表现出红卵,前期研究中对其进行初步的杂交遗传分析,推测Re-nd可能是隐性非致死性遗传(林英等, 2007)。在本研究中,通过Re-nd与野生型滞育的二化品种大造和非滞育多化品种N4杂交,F1代再分别以雌雄正反回交(半同胞交配)大造、N4亲本和进行F1代自交(同胞交配),统计这些杂交后代非滞育表型下单个蛾圈中蚕卵的卵色分离(图1),已可明确证实Re-nd属于独立的显性遗传,矫正了作者等之前鉴定的隐性遗传的看法(林英等, 2007)。通过表型观察,该突变体相对于其他卵色突变基因的筛选标记,其颜色鲜明且在非滞育情况下以显性呈现,这易于筛选转基因杂合子,作为分子标记在应用方面有潜力。
同时,本研究通过将Re-nd与其他已知突变基因的卵色突变品系进行杂交,根据F2代的卵色分离情况,以推测Re-nd突变基因可能位于卵色代谢途径的位置。由于已知卵色突变体在滞育的情况下,w-2卵呈现淡黄色、pe卵呈现淡黄偏微红色、re卵呈现红色;但在非滞育的情况下,这几个突变品系的卵都呈现淡黄色(Zhangetal., 2017)。而Re-nd突变体在滞育情况下呈现褐色,在非滞育情况下呈现艳丽的红色 (林英等, 2007)。通过对Re-nd突变体杂交遗传分析(图2~4),暗示了Re-nd突变基因应位于w-2及pe突变基因的下游,并且有别于re突变基因,可能参与了眼色素生物合成后期3-羟基-犬尿氨酸进一步代谢合成奥马丁类(ommatins)色素的过程。目前对家蚕卵色的眼色素生物合成这一后期代谢途径的研究尚不明确,且已知代谢途径的研究均是在卵发生滞育的状态下进行的,对非滞育状态下的研究更不清楚。所以家蚕非滞育红卵Re-nd突变体的意义就在于它属于显性突变体,并且是在非滞育情况下呈现艳丽红色卵的表型突变。因此,本研究结果为进一步通过定位克隆解析Re-nd突变体的突变基因提供了重要依据。利用现代分子生物学技术深入研究Re-nd突变基因的功能,将有助于阐释和完善家蚕色素代谢通路,探明非滞育情况下家蚕卵的着色机理。
