云南锦斑蛾成虫触角、喙管和跗节感器超微结构
2021-01-12李根层赵昱杰李嘉莉鲁国艳刘乃勇
李根层, 赵昱杰, 李嘉莉, 鲁国艳, 刘乃勇
(西南林业大学, 云南省森林灾害预警与控制重点实验室, 昆明 650224)
自然状态下,昆虫时刻接收着来自外界环境中寄主或非寄主植物、配偶、天敌等释放的各种化学信号(植物挥发物和信息素),进而完成寄主搜寻和定位、两性交配、敌害避免等重要生命活动(Altner and Loftus, 1985; Lopesetal., 2005)。昆虫对外界化学信号的捕捉和识别主要由嗅觉和味觉系统完成,通过触角、喙管、跗节等主要化感器官表面着生的具有多孔结构的化学感受器,外界环境中的气味分子从感受器上的微孔进入并激活嗅觉或味觉感受神经元,从而将化学信号转换为电信号(Leal, 2013)。根据化学感受器的着生方式、形态特征和功能,鳞翅目(Lepidoptera)昆虫的感受器可划分为毛形感器(sensilla trichodea)、刺形感器(sensilla chaetica)、锥形感器(sensilla basiconica)、鳞形感器(sensilla squamiformia)、腔锥形感器(sensilla coeloconica)、Böhm氏鬃毛(Böhm bristles)、栓锥形感器(sensilla styloconica)、耳形感器(sensilla auricillica)、板形感器(sensilla placodea)等(Schneider, 1964; 马瑞燕和杜家纬, 2000; 杨慧等, 2008b)。
鳞翅目是昆虫纲第二大目,包括蛾类和蝶类,触角、喙管和跗节是其主要的化感器官,负责感受和识别外界环境中的各种气味分子(Heppner, 2008)。触角是鳞翅目昆虫的主要嗅觉信号分子处理器,其表面着生有不同类型的感器,负责识别植物挥发物、性信息素、二氧化碳(carbon dioxide, CO2)等气味分子(Hansson and Stensmyr, 2011)。此外,鳞翅目有喙亚目成虫还具有喙管,是其味觉信号分子的处理器(Krenn, 2010);类似地,跗节也具有味觉功能(Calasetal., 2009)。与触角相比,喙管和跗节表面着生的感器种类和数量相对较少。唐丽萍等(2016)在竹斑蛾Artonafuneralis触角上鉴定到刺形感器、毛形感器、芽形感器、栓锥形感器和腔锥形感器,其中芽形感器仅分布在雌蛾触角上。茶尺蠖Ectropisobliqua触角上共有8类16种亚型的感器,而跗节上仅有3类5种亚型的感器(Maetal., 2016)。王娜等(2012)和薛爽(2014)研究了夜蛾亚科(Noctuinae)的5种昆虫和金翅夜蛾亚科(Plusiinae)的24种昆虫的喙管,发现前者喙管上有刺形感器、锥形感器和栓锥形感器;后者喙管上具有刺形感器、锥形感器、栓锥形感器和腔锥形感器。类似地,关于触角、喙管和跗节感器的研究在夜蛾科(Noctuidae)(刘金香等, 2009; Zenkeretal., 2011; Xue and Hua, 2014; Changetal., 2015)、卷叶蛾科(Tortricidae)(Waltersetal., 1998; 杨慧等, 2008a)、麦蛾科(Gelechiidae)(Bawinetal., 2017; 徐伟等, 2019)、斑蛾科(Zygaenidae)(Liuetal., 2018)等蛾类昆虫中也有报道。
云南锦斑蛾Achelurayunnanensis隶属于鳞翅目斑蛾科锦斑蛾属Achelura,分布于我国西南部至印度等区域;其幼虫主要危害樱属Cerasus, 李属Prunus, 球花石楠Photiniaglomerata和酸木瓜Chaenomelessinensis等多种蔷薇科(Rosaceae)植物,严重时可将整株树木叶片全部吃光,造成园林景观破碎化。目前,关于云南锦斑蛾的研究仅限于其形态特征、生物学特性等报道(黄圣卓等, 2013; 荣昌鹤等, 2016; 田茂寻等, 2018),其化学感受器及化学通讯方面的研究尚属空白。鉴于触角、喙管和跗节等主要化感器官在昆虫化学通讯中的重要性,本研究采用光学显微镜和扫描电镜等技术研究了云南锦斑蛾雌雄成虫触角、喙管和足跗节的形态特征及其感器种类、数量和超微结构,并比较了雌雄虫间感器的差异。研究结果有助于明确云南锦斑蛾成虫化学感受的分子机制,同时也为深入研究云南锦斑蛾成虫的觅偶、避敌、产卵、取食等行为机制奠定基础,最终可为其绿色防控技术的研发提供理论指导。
1 材料与方法
1.1 供试昆虫
云南锦斑蛾采自云南省昆明市区球花石楠植物上(25°02′11″N, 102°42′31″E,海拔1 891 m)。野外采集云南锦斑蛾的幼虫或蛹,带回实验室置于饲养箱内(饲养条件:温度26±1℃,相对湿度60%±5%,自然光周期),蒙上纱布并盖上带有小孔的盖子进行饲养。待成虫羽化后,将雌雄成虫分开饲养,并喂以10%的蜂蜜水。
1.2 成虫及其化感器官的外观形态观察
收集羽化后第4天的云南锦斑蛾雌雄成虫及其触角、喙管和跗节,用于雌雄虫外部形态特征观察。首先,将成虫在泡沫板上固定整姿并制作针插标本,放置于室内自然干燥,用照相机进行拍照。其次,取下成虫的触角、喙管和跗节,将其置于75%的乙醇溶液中,用SG9200HE超声波清洗器洗涤3次,每次3 min,除去组织表面粘附物。然后,待样品完全干燥后,用SteREO Discovery V20体视显微镜对触角、喙管和跗节进行观察和拍照。
1.3 触角、喙管和跗节上的感器鉴定
取羽化后第4天的云南锦斑蛾雌雄成虫各5头,每头成虫分别剪下1根触角、1根喙管和1只前、中、后足跗节,用于雌雄虫感器种类鉴定。首先,将取下的触角、喙管和跗节浸入75%的乙醇溶液中,采用超声波清洗器洗涤除去组织表面粘附物。而后,用100%乙醇脱水5 min,取出放在无菌操作台内风干。接着,将完全干燥后的雌雄虫触角、喙管和跗节分别用导电胶固定在样品台上,用日立E-1010型号的离子溅射仪进行镀金。最后,在日立S-3000N扫描电镜下进行观察和拍照,加速电压为15 kV或20 kV。
触角、喙管和跗节感器的鉴定和命名主要参照Schneider(1964), Faucheux(2013)和Calas等(2009)的方法,根据扫描电镜图片对每种感器进行鉴定。
1.4 数据处理与分析
采用ImageJ软件(NIH, Bethesda, MD, 美国)测量每种感器的长度。如果某种感器在雌雄虫化感器官上均有分布,则雌雄虫触角上各测量至少4根;如果某种感器仅在雌虫或雄虫触角上分布,则测量至少8根以上。采用IBM SPSS Statistics 20软件计算每组数据的平均值和标准误,并以t检验对雌雄触角同种感器的长度进行差异显著性分析。
2 结果
2.1 成虫及其化感器官的形态特征
2.1.1成虫及其触角的形态特征:云南锦斑蛾雌雄成虫虫体呈黑灰色,头为黑色,复眼大;翅薄,近透明,雄虫翅面颜色较雌虫深,翅面有两条红色斑纹,分别位于前翅基部及近基部1/3处(图1: A, B)。成虫触角为双栉齿状,黑色,由柄节、梗节和鞭节组成,长度上从大到小依次为:鞭节、梗节、柄节。雌雄虫触角柄节和梗节各1节,鞭节雄虫共104节,雌虫共83节,雌雄虫鞭节上均具有许多侧枝(图1: C)。
2.1.2喙管的形态特征:云南锦斑蛾雌雄成虫喙管不用时呈螺旋状,盘缩卷曲在头部下方,从基部至顶端逐渐变细(图2: A, B)。扫描电镜结果表明,云南锦斑蛾雌雄成虫的口器为虹吸式,下颚的外颚叶极度延长,左右2个外颚叶通过背连接片和腹连接片连锁,形成喙管;雌雄成虫喙管在大小和形状上无明显差异,整体表面较为粗糙,密布表皮突起物(图2: B, C)。
2.1.3跗节的形态特征:云南锦斑蛾雌雄成虫跗节均为5节,其中第1跗节较长;第2与第3跗节几乎等长,但均短于第1跗节;第4跗节短小,端部稍膨大;第5跗节小,心形,基部具2个侧爪,侧爪构造简单,无附齿(图3: A, B)。
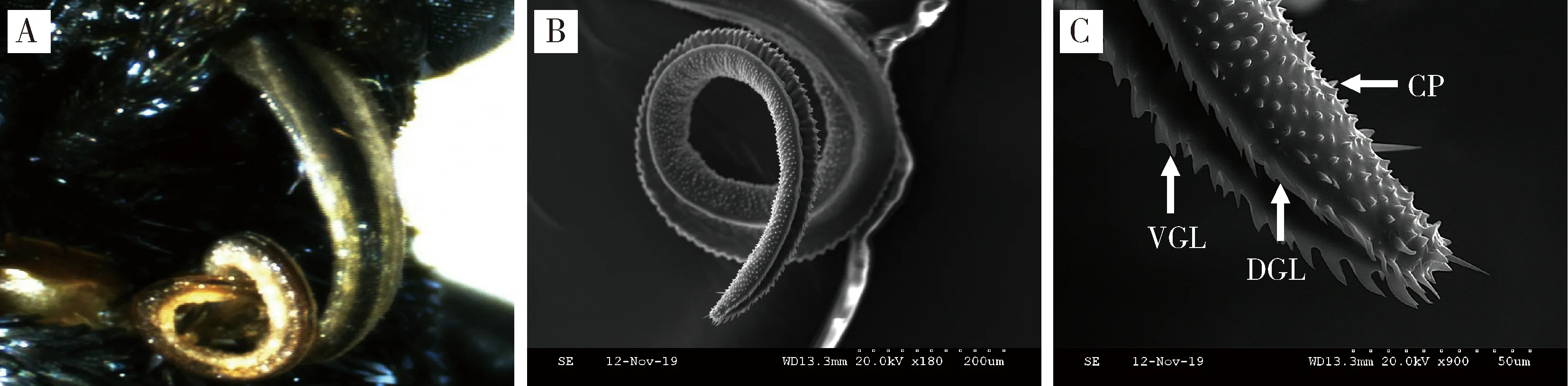
图2 云南锦斑蛾成虫的喙管
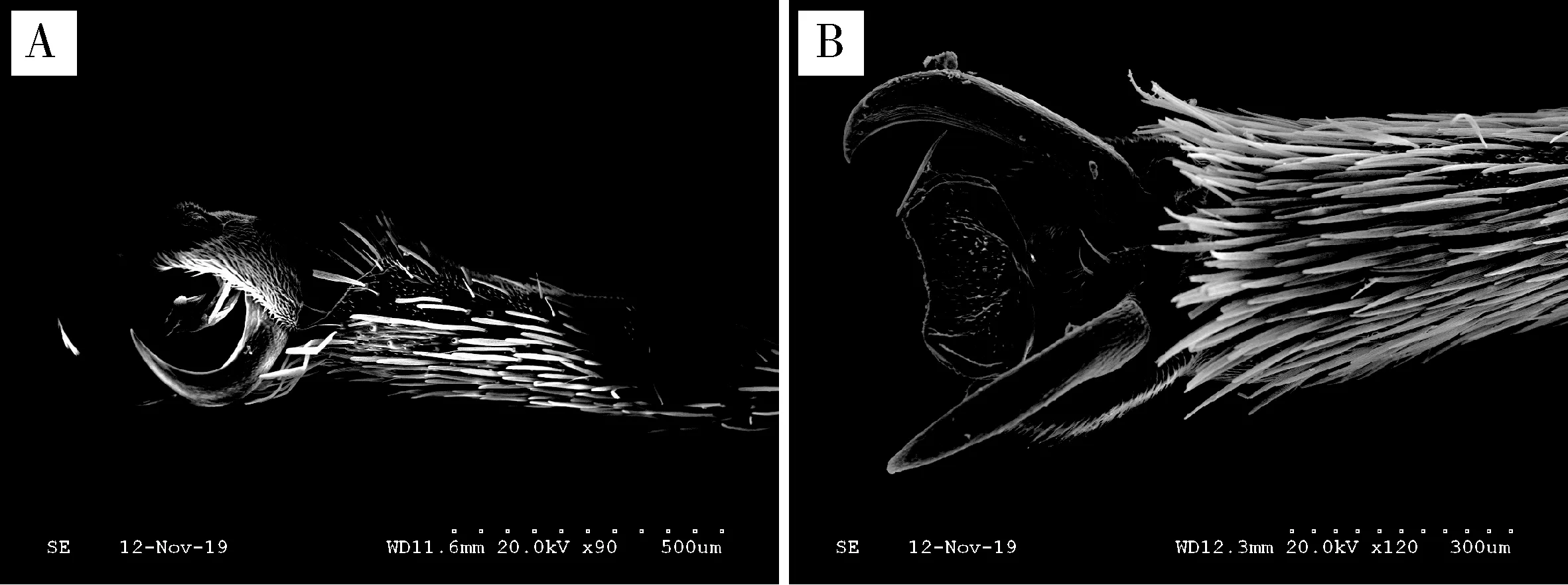
图3 云南锦斑蛾成虫跗节
2.2 成虫触角的感器类型
通过扫描电镜观察,发现云南锦斑蛾雌雄成虫触角上共有7类17种感器,包括毛形感器(3种)、锥形感器(5种)、刺形感器(3种)、Böhm氏鬃毛(2种)、耳形感器(1种)、腔锥形感器(2种)和鳞形感器(1种),其中毛形感器、锥形感器、刺形感器、Böhm氏鬃毛和腔锥形感器均存在不同亚型。
2.2.1毛形感器(sensilla trichodea, ST):毛形感器是大部分蛾类昆虫触角表面分布最广、数量最多的感器,呈毛发状,感器表面具倒“V”字形纵脊,具有机械和化学感受双重功能(Schneider, 1964; 马瑞燕和杜家纬, 2000; 杨慧等, 2008b; Zhangetal., 2013)。毛形感器是云南锦斑蛾雌雄成虫触角表面数量最多的感器,根据其外部形状和大小可将其分为3种亚型。
毛形感器I型(ST-I):雌虫感器长49.32±0.88 μm,雄虫感器长39.96±0.31 μm,雌虫感器长度显著大于雄虫(P<0.05);直立或微斜立于触角表面,基部稍膨大,向端部逐渐变细,端部尖细,感器整根弯曲;广泛分布于触角各节(图4: A; 表1)。
毛形感器II型(ST-II):雌虫感器长35.55±5.36 μm,雄虫感器长79.87±3.48 μm,雄虫感器长度显著大于雌虫(P<0.05);着生于基窝中,紧贴触角表面或夹角小于30°,由基部向端部逐渐变细,端部尖细,感器基部弯曲,表面具有纵纹(图4: B; 表1)。
毛形感器III型(ST-III):雌虫感器长41.92±0.31 μm,雄虫感器长94.92±7.61 μm,雄虫感器长度显著大于雌虫(P<0.05);着生于突出的基窝中,细长,由基部向端部逐渐变细,端部弯曲,收缩不明显(图4: C; 表1)。
2.2.2锥形感器(sensilla basiconica, SB):一般具有圆台形或圆锥状隆起的底座,中央着生形状不同的小锥状感器,主要负责昆虫对寄主或非寄主植物气味的感受和识别(Schneider, 1964; 马瑞燕和杜家纬, 2000; 杨慧等, 2008b)。云南锦斑蛾雌雄成虫触角表面共有5种亚型的锥形感器,数量仅次于毛形感器。
锥形感器I型(SB-I):雌雄虫感器分别长14.25±0.68和17.09±1.65 μm;深陷于较宽的基窝内,表面光滑,基部稍宽扁,端部钝圆,整根弯曲呈“C”形或稍弯曲(图4: D, E; 表1)。
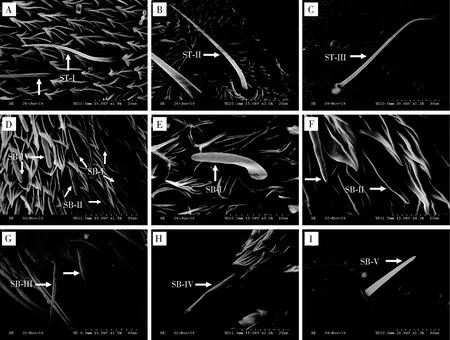
图4 云南锦斑蛾成虫触角毛形感器和锥形感器的类型及形态
锥形感器II型(SB-II):雌雄虫感器分别长14.65±0.25和17.09±1.65 μm;斜生于基窝内,与触角表面夹角小于30°,基部微粗且隆起,小锥状感器比SB-I细,端部钝圆(图4: D, F; 表1)。
锥形感器III型(SB-III):雌雄虫感器分别长29.25±2.05和24.39±5.46 μm;着生于凹陷的基窝中,底座圆台形,表面光滑,端部较SB-I和SB-II尖细且稍分为两支;零星分布于触角柄节与梗节,数量稀少(图4: G; 表1)。
锥形感器IV型(SB-IV):雌雄虫感器分别长24.88±0.63和30.13±1.10 μm;底座不明显,表面光滑,端部较SB-III钝;大量分布于鞭节附枝深陷的基窝中(图4: H;表1)。
锥形感器V型(SB-V):雌雄虫感器分别长37.58±1.44和48.67±1.09 μm;着生于深陷、狭窄的圆台形基窝内,表面光滑,由基部向端部逐渐变细,端部尖(图4: I; 表1)。
2.2.3刺形感器(sensilla chaetica, SCH):刺状或镰刀状,顶端较尖,基部钝圆,有臼状窝,表面有明显的条纹;不同亚型间长度变异较大,分布于触角鞭节的背面、腹面和侧面,具有感受机械刺激、化学刺激和温湿度的功能(马瑞燕和杜家纬, 2000; 杨慧等, 2008b)。云南锦斑蛾雌雄成虫触角上分布有3种亚型的刺形感器。
刺形感器I型(SCH-I):雌雄虫感器分别长34.17±0.59和26.41±7.65 μm;刚毛状,着生于浅窝内,基部粗壮,端部收缩明显且尖锐,表面具有明显纵纹;分布于触角柄节及梗节(图5: A; 表1)。
刺形感器II型(SCH-II):雌雄虫感器分别长30.54±0.78和32.52±0.75 μm;着生于臼状窝,基部向端部逐渐变细,端部尖细锐利且向触角端部方向弯曲;分布于触角柄节及梗节(图5: B; 表1)。
刺形感器III型(SCH-III):雌雄虫感器分别长47.56±1.88和38.36±1.05 μm,差异显著(P<0.05);刚直粗壮,基部向端部逐渐变尖,表面有明显的纵纹;分布于触角鞭节背面及附枝(图5: C; 表1)。
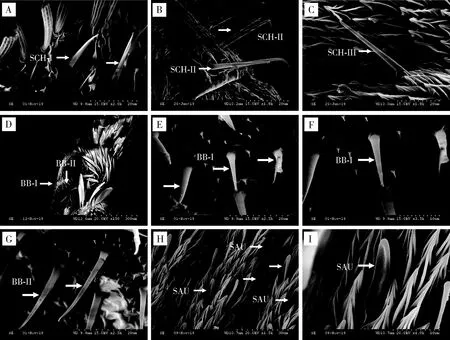
图5 云南锦斑蛾成虫触角刺形感器、Böhm氏鬃毛和耳形感器的类型及形态
2.2.4Böhm氏鬃毛(Böhm bristles, BB):表面光滑,似芽形,末端尖而弯曲,具有机械感受功能(Schneider, 1964; Meriveeetal., 2002)。云南锦斑蛾雌雄成虫触角上有2种亚型的Böhm氏鬃毛,主要分布于雌雄成虫触角柄节的基部,数量较少。
Böhm氏鬃毛I型(BB-I):该种感器仅在雄虫触角上有分布,感器长11.18±0.69 μm;直立于触角表面,表面光滑,由基部向端部逐渐变细,端部收缩明显(图5: D, E, F; 表1)。
Böhm氏鬃毛II型(BB-II):雌雄虫感器分别长21.20±2.08和22.48±2.86 μm;直立或微斜立于触角表面,表面光滑,基部稍膨大,向端部逐渐变细,端部尖细锐利,微微弯曲(图5: D, G; 表1)。
2.2.5耳形感器(sensilla auricillica, SAU):外形类似禾本科植物卷心叶片状,具有耳状凹槽,可能具有接受声波及探测植物气味的作用(杨慧等, 2008b; Rohetal., 2016)。云南锦斑蛾雌雄成虫触角上仅有1种耳形感器,分别长19.42±7.01和19.85±0.25 μm;大量着生于触角附枝基窝中,与触角表面夹角小,近乎平行,表面内陷呈耳形,端部钝圆(图5: H, I; 表1)。
2.2.6鳞形感器(sensilla squamiformia, SSQ):形状类似剑鞘,表面结构与普通鳞片相似,但比一般鳞片狭且端部较尖,基部有臼状窝,表面具有纵脊(杨慧等, 2008b)。云南锦斑蛾雌雄成虫触角表面仅有1种鳞形感器,分别长97.16±4.75和94.43±0.69 μm;纵纹明显,且每两条纵纹间具横纹,两者形成网状结构,分布于除附枝外的整根触角(图6: A, B; 表1)。
2.2.7腔锥形感器(sensilla coeloconica, SCO):触角表皮下陷形成的浅圆形腔,中央着生感觉锥,具有感受水蒸气、二氧化碳、湿度变化、植物气味等作用(den Otter and Thomas, 1979; Altner and Prillinger, 1980)。云南锦斑蛾雌雄成虫触角表面有2种亚型的腔锥形感器,且雌虫触角的腔锥形感器数量多于雄虫。在长度上,两者均表现为雄虫显著大于雌虫(P<0.05)(表1)。
腔锥形感器I型(SCO-I):雌雄虫感器分别长3.51±0.16和7.84±0.27 μm;着生于浅圆形腔中,感觉锥小,乳突状,顶端收缩明显、钝圆,表面具纵纹,有孔(图6: C; 表1)。
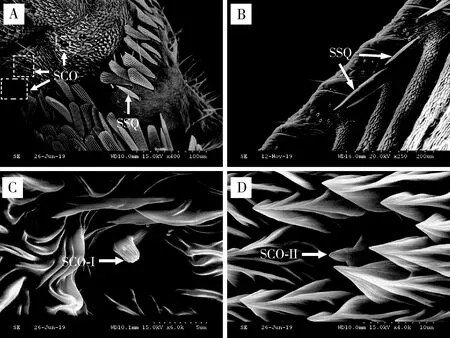
图6 云南锦斑蛾成虫触角鳞形感器和腔锥形感器的类型及形态
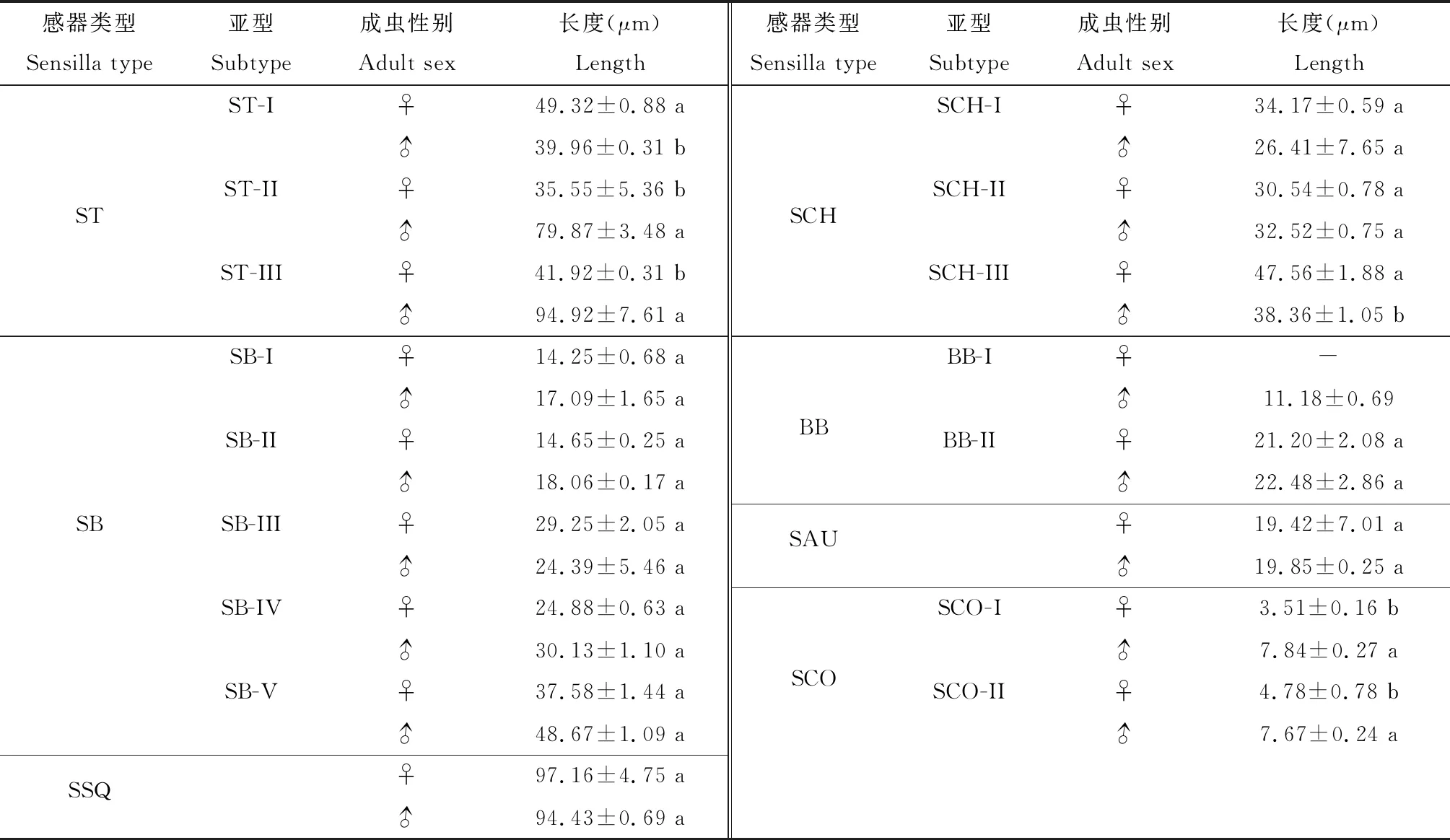
表1 云南锦斑蛾不同性别成虫触角不同类型感器的长度
腔锥形感器II型(SCO-II):雌雄虫感器分别长4.78±0.78和7.67±0.24 μm;着生于浅圆形腔中,基部膨大,感觉锥小,凿状,端部稍尖,表面具纵纹,有孔(图6: D; 表1)。
2.3 成虫喙管感器类型
通过扫描电镜观察,发现云南锦斑蛾雌雄成虫喙管上共有2类5种感器,包括锥形感器(4种)和刺形感器(1种),其中锥形感器存在不同亚型。
2.3.1锥形感器I型(SB-I):雌雄虫感器分别长5.61±0.89和3.15±0.36 μm;基窝圆台形,较短,直立,端部钝圆,外壁光滑(图7: A, B, C, D; 表2)。
锥形感器II型(SB-II):雌雄虫感器分别长10.70±1.46和14.04±0.59 μm;着生于圆柱形基窝内,较SB-I长,表面光滑,端部较SB-I尖(图7: A, B, C, E; 表2)。
锥形感器III型(SB-III):该种感器仅在雌虫触角上有分布,感器长18.83±0.12 μm;深陷于狭窄的圆台形基窝内,基部粗,端部尖细锐利,表面光滑;分布在雌虫喙管顶端(图7: F; 表2)。
锥形感器IV型(SB-IV):雌雄虫感器分别长25.45±1.38和26.42±1.94 μm;着生于深陷、宽阔的圆台形基窝内,小锥状感受器尖锐,表面光滑,尖端稍稍弯曲(图7: A, G; 表2)。
2.3.2刺形感器(SCH):雌雄虫感器分别长37.43±4.31和33.27±1.44 μm;尖刺状,斜立于喙管表面,表面具纵纹,由端部向基部逐渐变细,端部尖锐;分布于喙管正面和侧面、数量少(图7: A, H, I; 表2)。
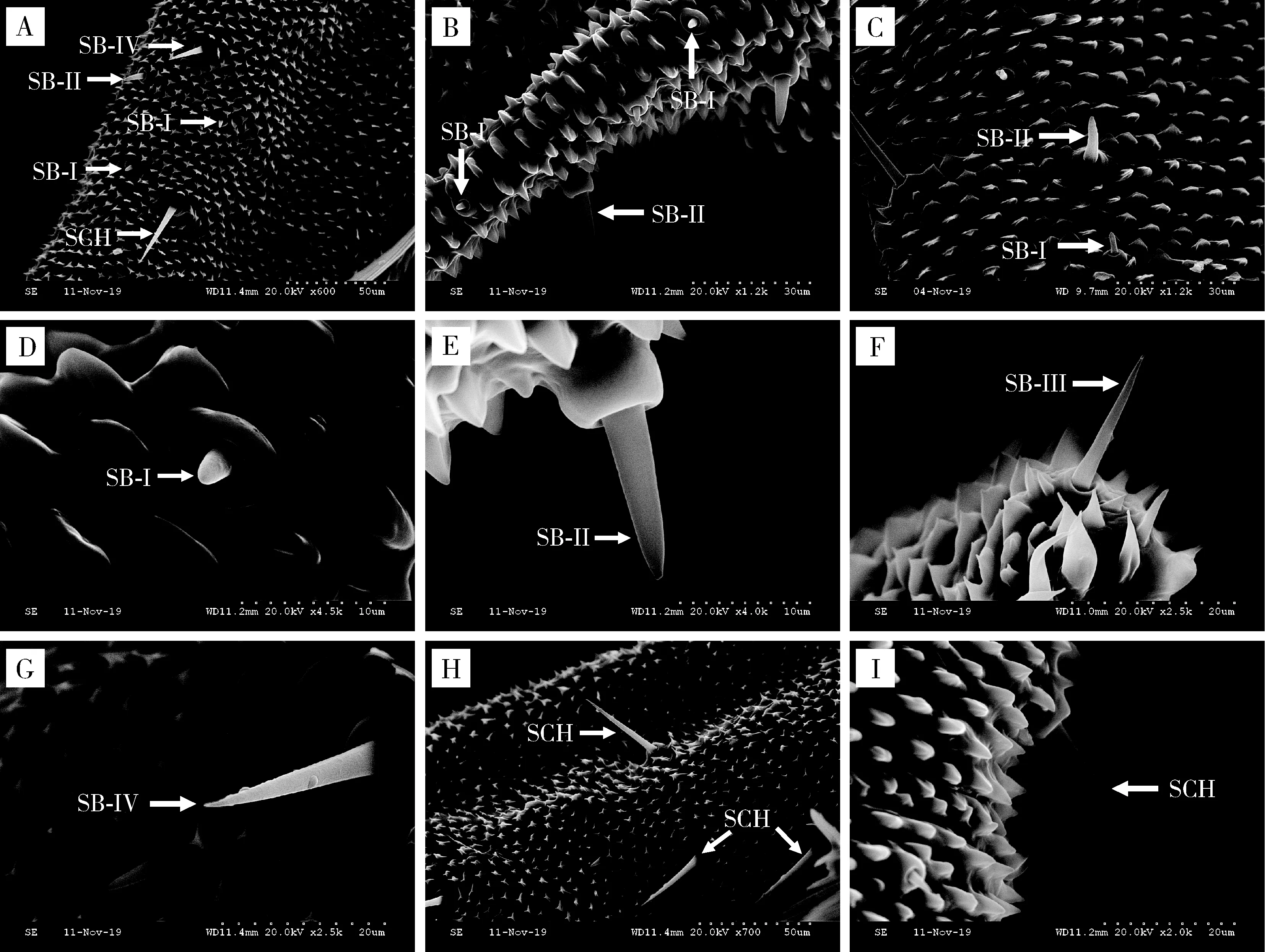
图7 云南锦斑蛾成虫喙管锥形感器和刺形感器的类型及形态
2.4 成虫跗节感器类型
通过扫描电镜观察,发现云南锦斑蛾雌雄成虫跗节上共有5类13种感器,包括毛形感器(4种)、鳞形感器(2种)、锥形感器(4种)、刺形感器(1种)和Böhm氏鬃毛(2种),其中毛形感器、鳞形感器、锥形感器和Böhm氏鬃毛具有不同亚型。
2.4.1毛形感器I型(ST-I):该种感器仅在雌虫触角上有分布,感器长59.45±4.60 μm;斜生于跗节表面的基窝内,与跗节表面夹角小于30°,稍稍弯曲,端部较尖(图8: A; 表3)。
毛形感器II型(ST-II):雌雄虫感器分别长66.05±0.18和59.25±2.57 μm;着生于深陷的基窝中,与跗节表面夹角在60°~90°之间,基部较粗,整根弯曲, 端部尖细; 分布于两侧爪间(图8: B, C;
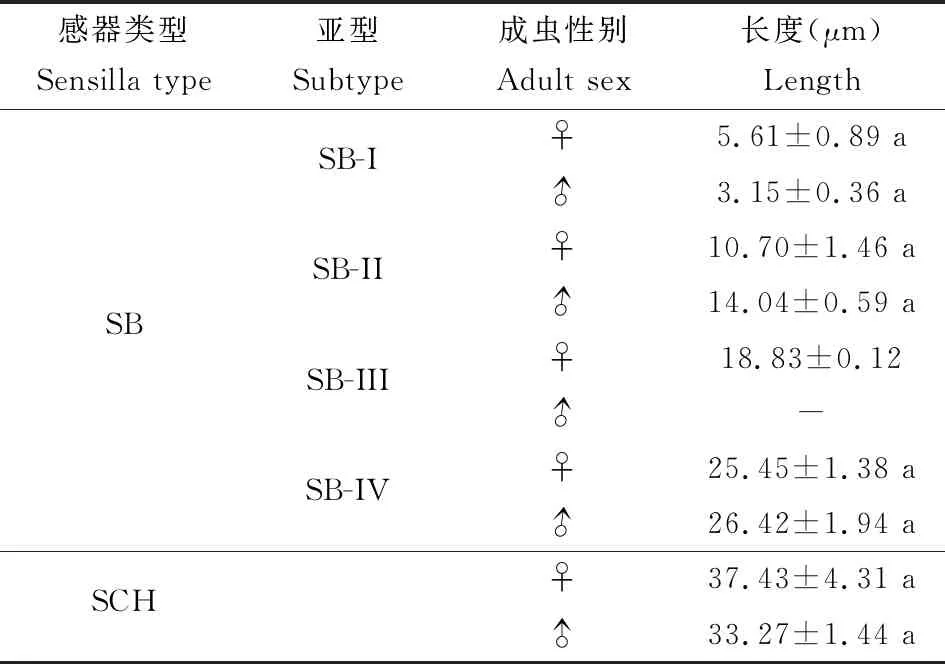
表2 云南锦斑蛾不同性别成虫喙管不同类型感器的长度
表3)。
毛形感器III型(ST-III):雌雄虫感器分别长74.03±2.83和97.48±0.71 μm,雄虫感器长度显著大于雌虫(P<0.05);着生于跗节表面的凹窝内,细长,微微弯曲或不弯曲,端部较尖(图8: D; 表3)。
毛形感器IV型(ST-IV):雌雄虫感器分别长63.51±6.31和112.41±3.32 μm,雄虫感器长度显著大于雌虫(P<0.05);毛发状,与跗节表面呈一定夹角,着生于跗节表面的凹窝内,细长,明显弯曲,表面具纵纹(图8: E, F; 表3)。
2.4.2鳞形感器I型(SSQ-I):雌雄虫感器分别长83.34±4.09和105.76±4.36 μm;着生于臼状窝内,整体圆锥,表面纵纹明显,端部钝圆(图8: G, H; 表3)。
鳞形感器II型(SSQ-II):雌雄虫感器分别长111.22±5.69和105.76±4.36 μm;着生于臼状窝内,整体剑鞘状,表面纵纹明显,端部钝宽,与触角中SSQ相似(图8: G, I; 表3)。
2.4.3锥形感器I型(SB-I):该种感器仅在雄虫触角上有分布,感器长30.59±6.22 μm;直立着生于圆台形基窝内,表面光滑,尖端分为两个分支,且两个分支较尖细(图9: A, B, C; 表3)。
锥形感器II型(SB-II):雌雄虫感器分别长44.97±2.73和26.48±2.05 μm,雌虫感器长度显著大于雄虫(P<0.05);直立着生于圆台形基窝内,呈钉状,表面光滑,尖端收缩明显且分为两个分支,但分支无SB-I明显(图9: A; 表3)。
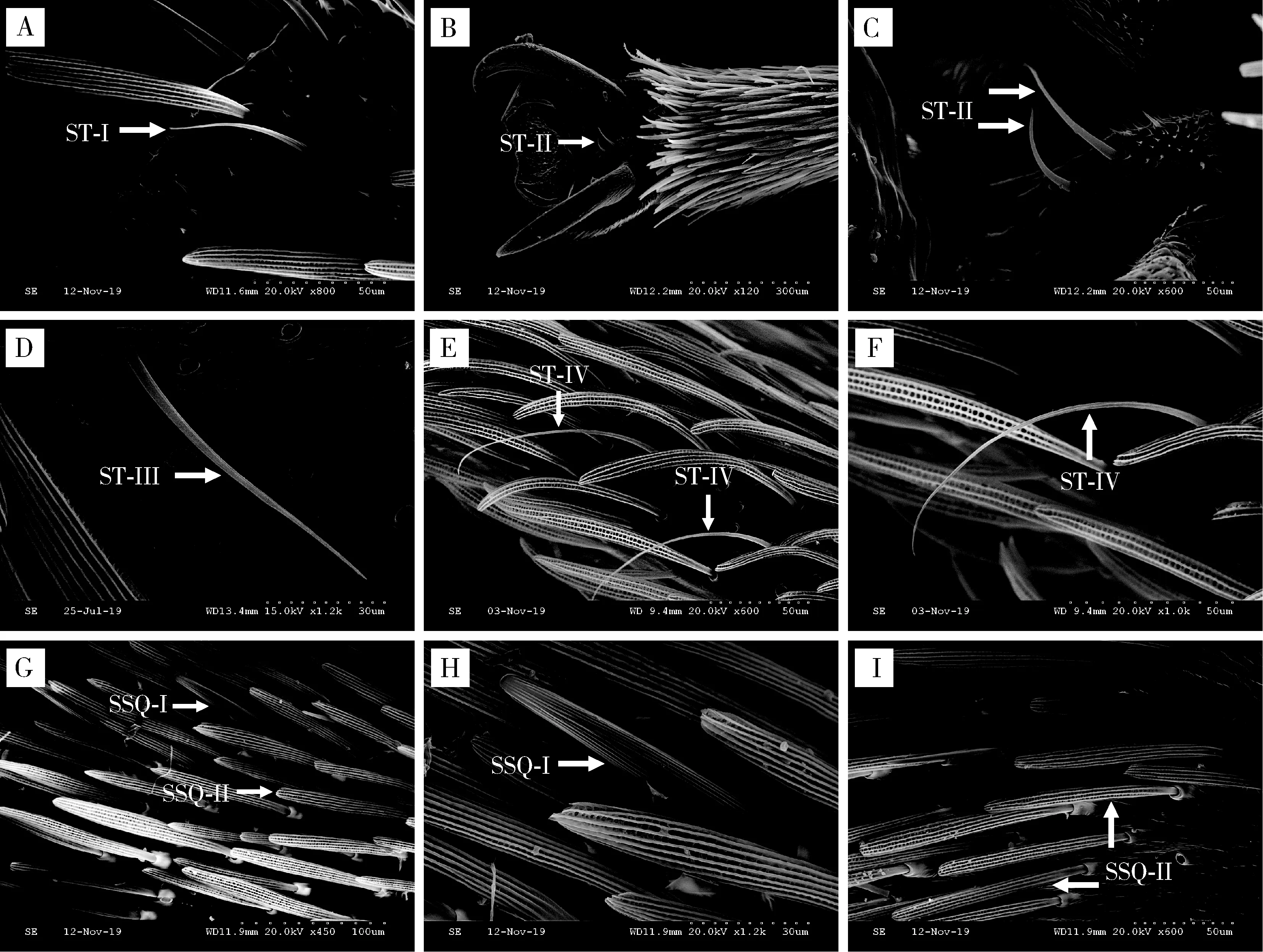
图8 云南锦斑蛾成虫跗节毛形感器和鳞形感器的类型及形态
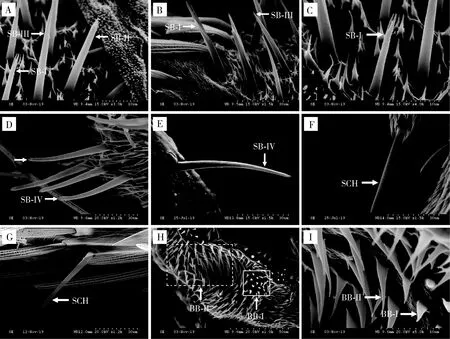
图9 云南锦斑蛾成虫跗节锥形感器、刺形感器和Böhm氏鬃毛的形态及类型
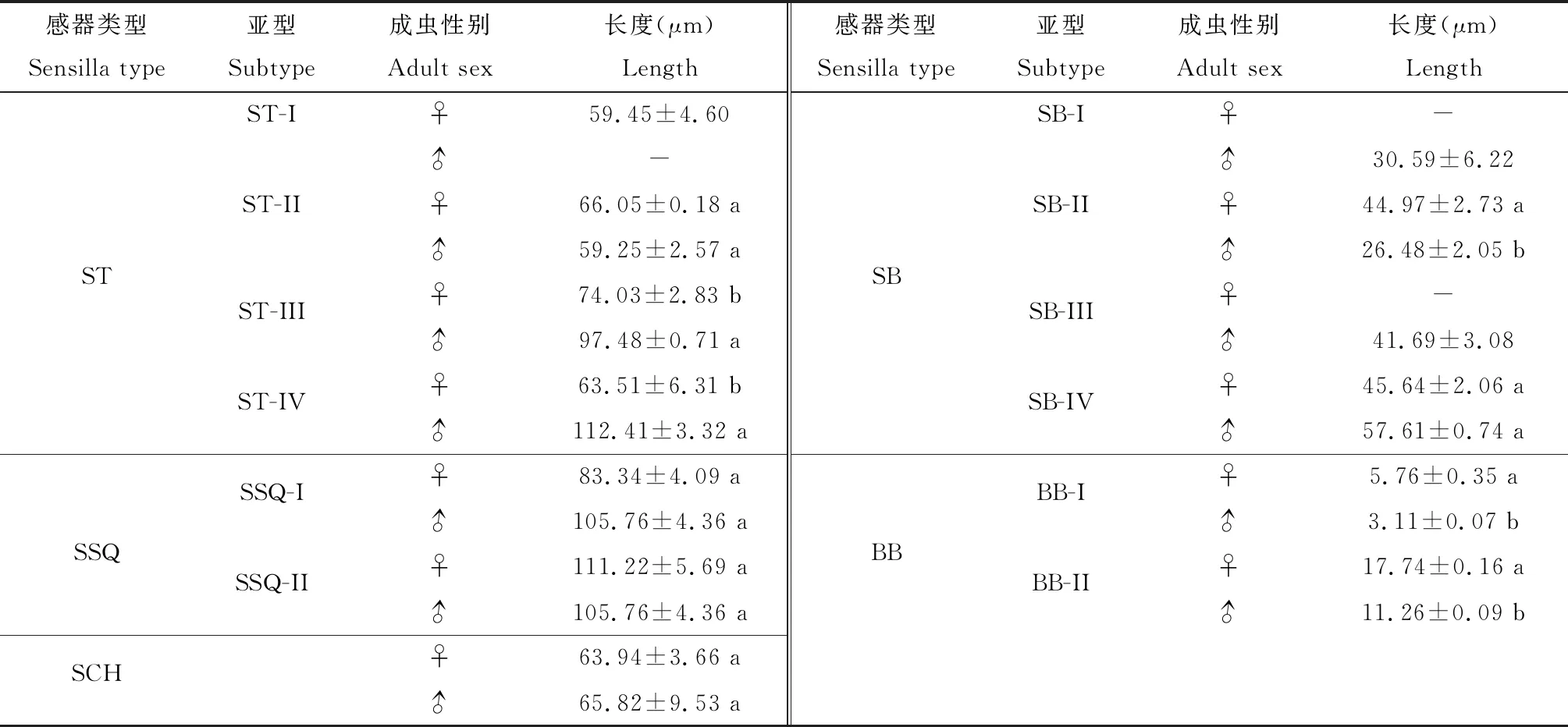
表3 云南锦斑蛾不同性别成虫跗节不同类型感器的长度
锥形感器III型(SB-III):该种感器仅在雄虫触角上有分布,感器长41.69±3.08 μm;直立或斜立于圆台形基窝内,表面光滑,较SB-I和SB-II长,尖端迅速缢缩成尖刺状(图9: A, B; 表3)。
锥形感器IV型(SB-IV):雌雄虫感器分别长45.64±2.06和57.61±0.74 μm;着生于圆台形基窝内,表面光滑,微微弯曲,顶端稍钝(图9: D, E; 表3)。
2.4.4刺形感器(SCH):雌雄虫感器分别长63.94±3.66和65.82±9.53 μm;刚直着生于跗节表面的基窝内,由基部向端部逐渐变细,稍弯曲,顶端尖(图9: F, G; 表3)。
2.4.5Böhm氏鬃毛I型(BB-I):雌雄虫感器分别长5.76±0.35和3.11±0.07 μm,雌虫感器长度显著大于雄虫(P<0.05);芽状,表面光滑,基部粗壮,顶端收缩明显,尖而稍弯曲(图9: H, I; 表3)。
Böhm氏鬃毛II型(BB-II):雌雄虫感器分别长17.74±0.16和11.26±0.09 μm,雌虫感器长度显著大于雄虫(P<0.05);斜生于跗节表面,表面光滑,基部较粗,端部尖细,较BB-I长(图9: H, I; 表3)。
2.5 雌雄成虫触角、喙管和跗节感器的差异
通过比较云南锦斑蛾雌雄虫化感器官的感器,发现成虫触角上的感器种类和数量均多于喙管和跗节上的(表1, 2和3)。在触角上,雄虫触角ST的数量多于雌虫;BB-I仅在雄虫触角上有分布;大部分同种亚型感器的平均长度为雄虫长于雌虫,其中雄虫ST-II,ST-III和SCO-I的长度是雌虫的两倍余(表1)。在喙管上,SB-III型仅在雌虫触角上有分布,雌雄成虫喙管上同种亚型的感器平均长度近似相等(表2)。此外,跗节上毛形感器和锥形感器在种类上具有性二型现象,其中SB-I和SB-III仅在雄虫跗节上有分布,而ST-I仅分布于雌虫跗节;长度上,雄虫跗节的ST-IV约为雌虫的2倍,而雌虫跗节的BB-I约为雄虫的2倍。
3 讨论
云南锦斑蛾是为害球花石楠、樱花树、红叶李、酸木瓜等蔷薇科园林植物的重要食叶害虫。近年来,云南锦斑蛾在我国西南地区持续发生危害,严重时常将整株植物的顶端叶片全部吃光,严重影响城市园林景观。因此,亟待研发绿色环保且有效的害虫防治措施。目前,针对该种害虫仅限于其形态特征和生活史(田茂寻等, 2018)、幼虫空间分布型(荣昌鹤等, 2015)、幼虫体表分泌物及活性测定(Owadaetal., 1999; 黄圣卓等, 2013)、幼虫杀虫剂抗性(荣昌鹤等, 2016)等方面的报道。本研究采用光学显微镜和扫描电镜技术,对云南锦斑蛾雌雄成虫触角、喙管和足跗节的形态特征及其感器种类、数量和超微结构进行了观察,并比较了雌雄虫间感器的差异,研究结果对于明确云南锦斑蛾化学感受机制,以及开发以化学通讯为基础的绿色防控技术具有重要指导意义。
研究发现,云南锦斑蛾成虫触角上具有7类17种感器,而喙管和跗节上分别为2类5种和5类13种,少于触角上的感器种类。考虑到蛾类昆虫主要用喙管吮吸花蜜、露水或果汁获取营养物质(Krenn, 2010),食物相对单一,因此感器种类最少。跗节在更多的时候需要接触到寄主或非寄主植物表面以及产卵引诱剂或驱避剂等(Calasetal., 2009),其感器种类较喙管多;而触角是云南锦斑蛾的主要嗅觉器官,需要感受外界环境中寄主或非寄主植物释放的挥发物、配偶产生的性信息素、天敌产生的气味、CO2等各种气味分子(Hansson and Stensmyr, 2011),可能需要更多不同类型的感器参与感受和识别不同类型的化合物。
在云南锦斑蛾触角上,感器种类数量多于同科的竹斑蛾(5类)(唐丽萍等, 2016)和朱红毛斑蛾Phaudaflammans(6类)(Liuetal., 2018),以及非斑蛾科的草地螟Loxostegesticticalis(5类)(尹姣等, 2004)和栎黄枯叶蛾Trabalavishnougigantina(5类)(靳泽荣等, 2016),但是少于松瘿小卷蛾Cydiazebeana(8类)(杨慧等, 2008a)、茶尺蠖(8类)(Maetal., 2016)和杨背麦蛾Anacampsispopulella(8类)(徐伟等, 2019)。与竹斑蛾相比,云南锦斑蛾触角上缺少芽形感器和栓锥形感器,但是新鉴定到锥形感器、耳形感器、鳞形感器和Böhm氏鬃毛(唐丽萍等, 2016);而相比于朱红毛斑蛾,云南锦斑蛾触角上缺少栓锥形感器,但是多耳形感器和鳞形感器(Liuetal., 2018),说明斑蛾科不同种间感器类型存在一定差异,从某种程度上可能反映了不同斑蛾科昆虫生境及寄主植物的差异性。与触角不同,鳞翅目蛾类昆虫喙管和跗节的感器研究相对较少,而在斑蛾科昆虫中仍未见报道。研究表明,鳞翅目昆虫的喙管上主要具有6种感器——锥形感器、刺形感器、栓锥形感器、腔锥形感器、丝状感器和钟形感器,其中前3种感器较为常见(Faucheux, 2013)。云南锦斑蛾喙管上仅有锥形感器和刺形感器,属于鳞翅目有喙亚目昆虫常见的感器类型(Zenkeretal., 2011; 薛爽, 2014; Xue and Hua, 2014; 谢佼昕, 2019)。在跗节上,云南锦斑蛾具有5类感器,多于核桃举肢蛾Atrijuglanshetaohei(4类)(刘桂湘等, 2018)和茶尺蠖(3类)(Maetal., 2016),但是少于蓝桉尺蛾Mnesampelaprivata(8类)(Calasetal., 2009)。因此,云南锦斑蛾是斑蛾科昆虫中首次报道喙管和跗节感器的昆虫,研究结果可为后续斑蛾科其他昆虫同类化感器官感器的鉴定和比较研究提供借鉴。
与大部分昆虫触角感器研究(尹姣等, 2004; Maetal., 2016; 唐丽萍等, 2016; Bawinetal., 2017; Liuetal., 2018; 尹宁娜等, 2019)类似,毛形感器是云南锦斑蛾触角上分布最广、数量最多的感器;其次是锥形感器和刺形感器。考虑到云南锦斑蛾成虫两性识别、寄主搜寻和定位、产卵和取食寄主识别等重要生命活动主要涉及到性信息素、植物气味、产卵引诱剂或驱避剂等的感受和识别,结合蛾类昆虫毛形感器、锥形感器和刺形感器的功能以及云南锦斑蛾触角上3种感器的数量,推测这些感器是云南锦斑蛾的主要嗅觉感器和机械感器,负责感受和识别外界环境中的气味和物理刺激。此外,云南锦斑蛾雄虫触角上的毛形感器较雌虫多,分析认为可能与雄虫需要寻找和定位雌虫有关,较多的毛形感器有利于提高择偶率。类似的结果在同科的竹斑蛾(唐丽萍等, 2016)和朱红毛斑蛾(Liuetal., 2018)中也有发现,说明雄虫触角毛形感器较雌虫多可能是斑蛾科大部分昆虫的共有特征。在家蚕Bombyxmori中,单感器记录研究表明腔锥形感器中的神经元能够被醛类、醇类和萜烯类等植物气味激活,说明这种感器参与到家蚕寄主或非寄主植物挥发物的嗅觉感受和识别过程(Pophof, 1997)。云南锦斑蛾雌虫触角上的腔锥形感器数量多于雄虫,与苹果蠹蛾Cydiapomonella(赵骁等, 2012)、香梨优斑螟Euzopherapyriella(马涛等, 2013)、桃蛀螟Conogethespunctiferalis(李倩等, 2014)和二点委夜蛾Athetislepigone(田彩红等, 2015)研究结果类似,说明这种感器可能参与到这些蛾类雌虫产卵寄主寻找等特定的生理活动。
鳞翅目昆虫的喙管是成虫获取营养物质的味觉器官,主要用于吮吸花蜜、果汁、动植物的腐烂汁液、泪液等(Krenn, 2010)。喙管的表面着生有不同类型的感器,具有感受机械刺激、蔗糖、乙醇等功能,例如小红蛱蝶Vanessacardui喙管上的刺形感器可感受花的直径和花冠深度(Krenn, 1998);柑橘凤蝶Papilioxuthus喙管上的锥形感器能够感受蔗糖溶液(Inoueetal., 2009);大红蛱蝶Vanessaindica和斐豹蛱蝶Argyreushyperbius喙管上的栓锥形感器对乙醇敏感,说明该种感器中具有感受乙醇分子的味觉神经元(muraetal., 2008)。云南锦斑蛾成虫主要以吮吸花蜜为食,喙管上的锥形感器可能具有感受花蜜中糖类等化合物的作用,而刺形感器可能参与感知外界环境引起的机械刺激。与喙管功能类似,鳞翅目昆虫的跗节也主要具有味觉功能。电生理研究表明,蓝桉尺蛾和西方蜜蜂Apismellifera的跗节感器能够感受糖类、盐、氨基酸或苦味化合物(Calasetal., 2009; de Brito Sanchezetal., 2014);棉铃虫Helicoverpaarmigera跗节上的9种感器对蔗糖、葡萄糖、果糖、麦芽糖、肌醇和20种常见氨基酸均有不同程度的反应,暗示这些感器在棉铃虫成虫的取食、产卵等生命活动中具有重要功能(Zhangetal., 2010)。据此,推测云南锦斑蛾跗节上的感器也可能具有感受糖类、盐或氨基酸等化合物的作用。
