金铁锁WRKY 转录因子基因鉴定及其蛋白序列特征和功能特性分析
2021-01-08张爱丽杨耀文李国栋钱子刚
江 舟,张爱丽,杨耀文,李国栋,钱子刚*
(1.云南中医药大学基础医学院,云南 昆明 650500;2.云南中医药大学中药学院,云南 昆明 650500)
转录因子是一类DNA 结合蛋白,能特异性结合基因启动子顺式作用元件,激活或抑制转录,通过靶基因来调控相关基因的表达[1]。WRKY 是一类植物所特有的转录因子[2],因N 端具有高度保守性的WRKYGQK 区域,C 端具有锌指C2H2或C2HC 结构而命名[3],能调控植物次生代谢产物合成、生长发育、抗病反应、抗胁迫能力[4-6]。
萜类作为天然次生代谢产物,广泛存在于植物中,尤其三萜类是中草药重要有效成分,具有较高的药用价值[7]。研究表明,许多植物萜类代谢途径的关键酶的表达可能受同一个转录因子的调控,故萜类代谢途径中有转录因子的存在[8]。目前,转录因子调控药用植物次生代谢产物的研究仅在红豆杉、黄花蒿、长春花、人参、西洋参[9-13]等少数几个植物中报道过。
金铁锁Psammosilene tunicoides W.C.Wu et C.Y.Wu为石竹科金铁锁属多年生植物[14],以根入药[15],是我国西南区域特有的传统药用植物,具有消炎、止痛、散瘀、祛风湿功效,是云南白药主要组成药材之一,前期大量研究表明其主要有效成分是齐墩果烷型五环三萜皂苷[16-20]。由于该药材显著的药理活性及药用价值,相关资源已开始匮乏,故揭示相关转录因子在其所含三萜皂苷合成途径中的调控机制,定向增加该成分积累显得尤为重要,但相关转录因子家族信息及关键酶基因表达、调控作用机理仍是未知领域。因此,本实验利用课题组前期已获得的金铁锁转录组数据,得到10 条含全长开放阅读框 (ORF) 的WRKY 家族成员相关信息,通过生物信息学方法分析其基因结构及蛋白序列特征和功能特性,为进一步研究该药材WRKY 转录调控机制及三萜皂苷合成机理提供分子依据。
1 方法
1.1 金铁锁WRKY 基因结构分析 金铁锁WRKY 转录因子(PtWRKY) 序列来自课题组测序得到的转录组数据库,使用以2 年生根和叶为材料通过Illumina Hiseq 2500 平台测序,ABYSS 软件拼接获得。以人参、西洋参WRKY CDS 为查询序列,通过BLASTn 进行同源搜索,ORF Finder (http://www.ncbi.nlm.nih.gov/ gorf/orFig.cgi) 在线筛选具有完整开放阅读框(ORF) 的WRKY 序列,SMART (http://smart.embl.de/) 和 Pfam (http://pfam.sanger.ac.uk) 确定选出的序列具有WRKY 保守结构域和DNA 结合结构域,Gene Structure Display Server (http://gsds.cbi.pku.edu.cn/) 对金铁锁WRKY 基因结构进行分析。
1.2 金铁锁WRKY 蛋白序列理化特征分析 金铁锁WRKY 蛋白理化性质的基本参数通过http://expasy.org/protparam/在线分析获得。
1.3 金铁锁WRKY 蛋白基序分析和预测 通过MEME(http://meme-suite.org/) 在线预测金铁锁WRKY 转录因子基序(motif),参数设定motif 最大值为3,宽度为3~100,并联合Clustal Omega (http:/ www.ebi./ ac.uk/Tools/) 对WRKY 转录因子保守结构域进行综合比对(参数OUTPUT FORMAT,ClustalW with character counts)。
1.4 金铁锁WRKY 转录因子聚类分析及进化树构建 从NCBI 数据库中下载人参、西洋参WRKY 氨基酸序列用于同源性分析,通过MEGA6.0 的Neighbor-Joining (参数Bootstrap method,Replications1000) 进行进化树构建。
1.5 金铁锁WRKY 转录因子结合位点预测及分析 通过JASPAR (http://jaspar.genereg.net/) 在线预测分析金铁锁WRKY 转录因子的转录结合位点(TFBS) 信息。
1.6 金铁锁WRKY 表达分析 根据转录组测序所得数据分析金铁锁WRKY 基因的组织表达模式,其丰度以PRKM为计量单位(Reads per kilobase per million mapped reads)。
2 结果
2.1 WRKY 基因结构分析 通过同源比对及开放阅读框(ORF) 预测,从PtWRKY 转录组数据库273 条序列中得到10 条具有完整ORF 的PtWRKY 序列,开放阅读框612 bp(DN23488_ c0g1)~1 116 bp (DN23438_ c0g4)。通过基因结构分析(图1) 发现,DN21294_ c0g3、DN23488_c0g1、DN23488_ c0i2、DN23488_ c0i6 均含有1 个内含子,10 条基因序列均含有非编码区,5′ UTR 0 bp (DN21294_c0g3,DN23488_ c0i6)~685 bp (DN19980_ c0g2),3′UTR 176 bp (DN24254_ c0g3)~1 030 bp (DN23488_c0i6)。
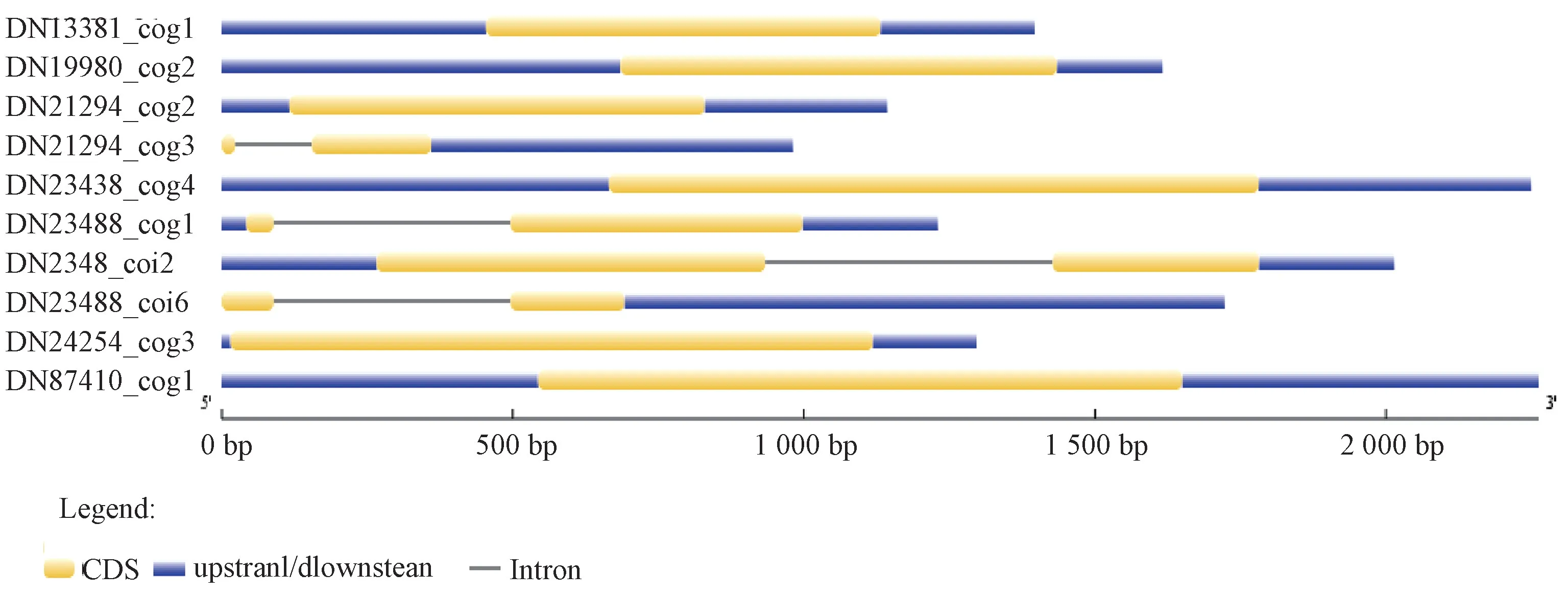
图1 金铁锁WRKY 基因结构分析
2.2 WRKY 蛋白序列理化特征分析 10 条开放阅读框包括612~1 116 bp 的PtWRKY 序列,编码蛋白203~371 aa,分析该序列理化特征,结果见表1。由此可知,蛋白相子量最小为23.0 kD,最大为43.1 kD,理论pI 平均值为7.68,其中有5 条蛋白的小于7,表明鉴定出的PtWRKY 蛋白偏酸性;所有PtWRKY 蛋白的亲水系数均为负数,表明该基因家族编码蛋白均亲水,稳定性较差。
2.3 WRKY 转录因子蛋白基序分析和预测 Eulgem 等[3]将WRKY 蛋白分为3 类:第Ⅰ类含有2 个WRKY 结构域,锌指结构为C2H2型;第Ⅱ类含有1 个WRKY 结构域,锌指结构也为C2H2型,第Ⅲ类有1 个WRKY 结构域,锌指结构为C2HC 型;Zhang 等[21]又进一步将第Ⅱ类WRKY 蛋白分为5 个亚型,分别为Ⅱa、Ⅱb、Ⅱc、Ⅱd、Ⅱe。本实验采用MEME 对金铁锁WRKY 的基序(motif) 进行在线预测分析,发现有3 个motif (图2A);Clustal Omega 对10 个PtWRKY 进行综合比对,分析motif 结构(图2B),发现除了DN21294_ c0g2 和DN23488_ c0i6 缺失锌指结构以外,DN13381 _ c0g1、DN19980 _ c0g2、DN24254 _ c0g3、DN21294 _ c0g3、DN23488 _ c0g1、DN23488 _ c0i2、DN87410_ c0g1、DN23438_ c0g4 都具有WRKY 保守结构域WRKYGQK、C2H2锌指结构,属于第Ⅱ类WRKY 蛋白。进一步分析该类蛋白的结构域特征,发现DN13381_ c0g1 属于Ⅱa 亚型,DN21294_ c0g3、DN23488_ c0g1、DN23488_c0i2 属 于Ⅱc 亚 型,DN23438_ c0g4、DN87410_ c0g1、DN24254_ c0g3 属于Ⅱd 亚型,DN19980_ c0g2 属于Ⅱe 亚型,无Ⅱb 亚型。另外,Ⅱa、Ⅱd、Ⅱe 亚型的锌指序列为C-X5-C-X23-H-X1-H,而IIc 亚型的锌指序列为C-X4-CX23-H-X1-H。

表1 金铁锁WRKY 蛋白理化特征
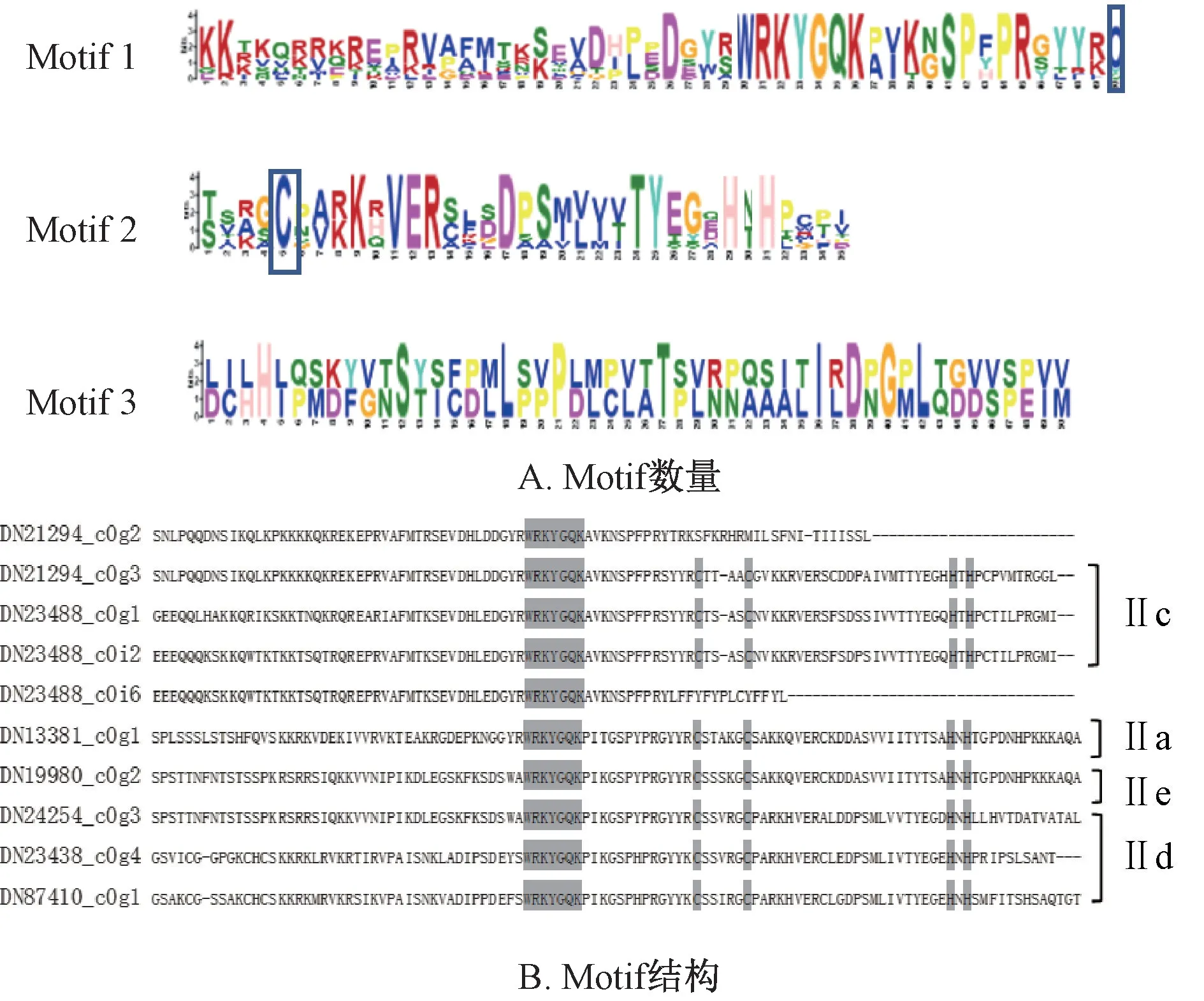
图2 金铁锁WRKY 蛋白基序分析
2.4 WRKY 转录因子聚类分析及进化树构建 通过MEGA6.0,以 10 个 PtWRKY、9 个人参 WRKY(PgWRKY)、3 个西洋参WRKY (PqWRKY) 转录因子序列构建系统进化树,通过已知的人参、西洋参WRKY 功能鉴定同源金铁锁WRKY,从而预测了解后者功能,结果见图3。由此可知,12 个WRKY 转录因子分为4 个大类Class1~Class4,金铁锁、人参WRKY 转录因子优先聚在一起,DN19980_ c0g2、PgWRKY9,以 及DN13381_ c0g1、PgWRKY28 被聚为一类,表明这2 种植物WRKY 转录因子有较高的亲缘关系和同源性,有着类似的功能。
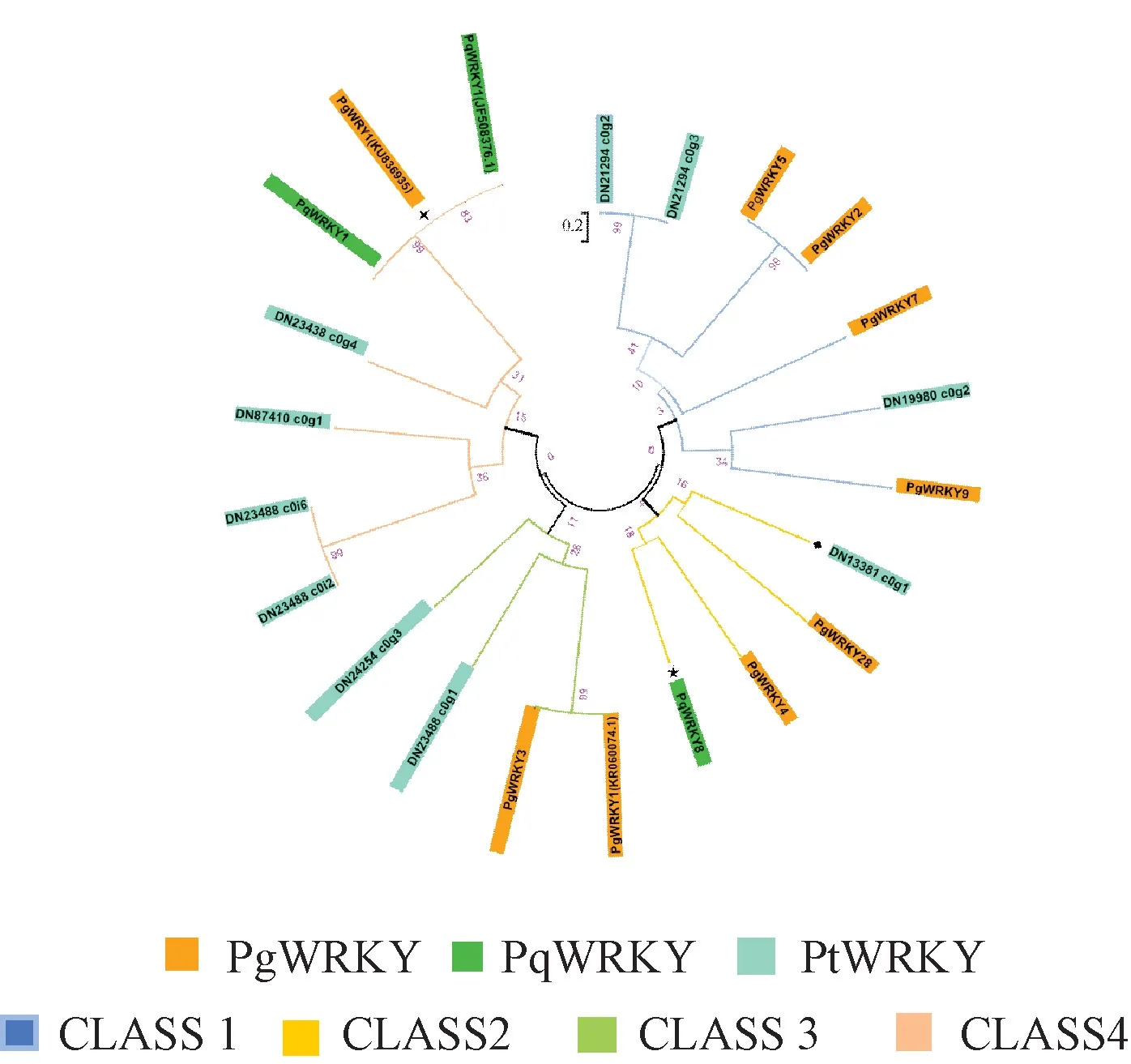
图3 金铁锁、人参、西洋参WRKY 蛋白的系统进化树
2.5 WRKY 转录因子结合位点预测及分析 转录因子蛋白调控特定基因的表达是通过与目的基因5′端上游部分的转录因子结合位点TFBS (transcription factor binding site) 特异性结合来实现的[22],研究表明TFBS 具有保守性,有相同或相似的该序列就可结合相同的转录因子[23]。将PtWRKY 转录因子蛋白序列输入JASPAR,筛选序列版本信息 和 e-value 值 后 (序列版 本 MA1093.1、MA1094.1、MA1087.1,e-value 值1.67014e-14、1.5632e-31、1.46487 e-31),预测得到3 个均属于WRKY 家族的TFBS 序列信息,分别为DN21294_ c0g2、DN21294_ c0g2、DN23488_c0i2 所有,相关报道见于拟南芥WRKY8,WRKY 75、WRKY 45[24],PFM Matrix (碱基频数分布矩阵) 见图4A1、4B1、4C1。通过TFFM (双核苷酸关联序列) 分析 (图4A2、4B2、4C2) 发现,前2 个TFBS 序列在3~7 位有相同的保守碱基(图4A1~4B2);后1 个在3~8 位均有保守碱基,在第8 位有差异,分别为C 和T。再分析motif 富集峰型图,发现在峰的相对位置0 处均出现了1 个尖锐峰(图4D1~4D3),表明该峰是转录因子motif 与TFBS 的结合位置。
2.6 WRKY 基因表达分析 对金铁锁10 个WRKY 基因在不同组织中的表达模式进行分析,发现PtWRKY 基因在根和叶中均有表达(PRKM>0) (图5),大多数在根中表达较高,只有Ⅱa 亚型DN13381_ c0g1、Ⅱe 亚型DN19980_c0g2 在叶中的表达量更高;Ⅱc 亚型DN23488_ c0g1、DN23488_ c0i2,Ⅱd 亚 型DN23438_ c0g4、DN24254_c0g3 在根中的表达量均大于20,其中DN23488_ c0i2 高达69.24,同时Ⅱc 亚型DN21294_ c0g3,Ⅱd 亚型DN87410_ c0g1、锌指结构缺失的DN23488_ c0i6 和DN21294_ c0g2表达量也均大于7,表明金铁锁根中高表达PtWRKY 基因主要属于Ⅱc、Ⅱd 亚型。
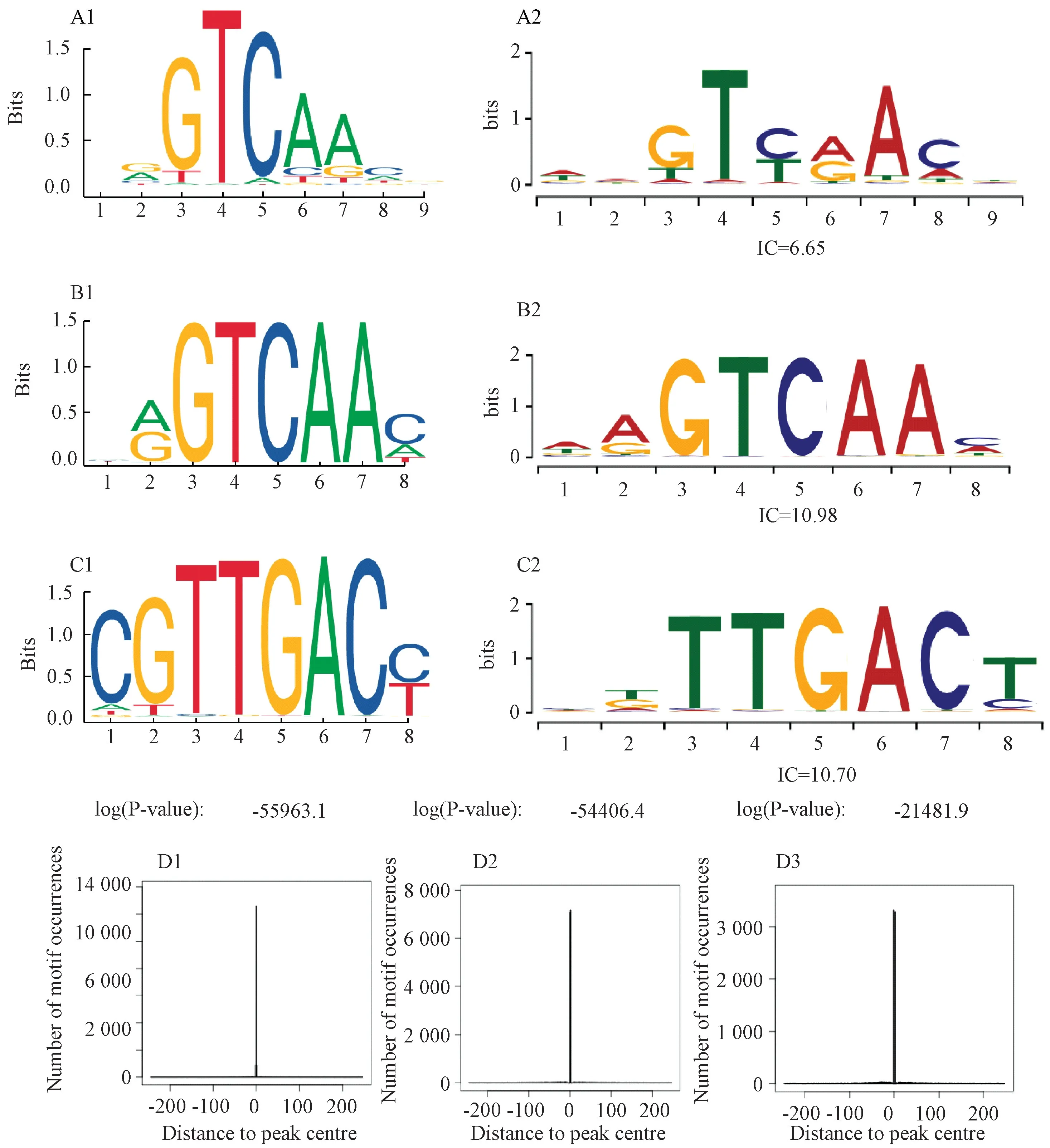
图4 金铁锁WRKY 转录因子结合位点的分析
3 讨论
金铁锁有效成分齐墩果烷型五环三萜皂苷结构复杂,在抗菌、消炎、止痛等方面的药理作用明确,是多种中成药的重要组成部分。长期以来,获取该药材方式几乎都靠野生采挖,但由于其分布狭窄,种群稀少,自然生长缓慢,导致自然资源面临枯竭。鉴于此,本实验开展金铁锁Pt-WRKY 转录因子家族的研究,以期获得可能参与其所含三萜皂苷合成转录的PtWRKY 基因,探索该成分合成调控规律,为实现其定向积累奠定基础,也为其获得提供新途径,从而实现该类濒危珍稀药材的可持续利用。
本实验分析了10 个金铁锁PtWRKY 基因,发现其中8个只具有1 个能与靶DNA 序列W-box 结合的保守结构域WRKYGQK,锌指结构为C2H2,归为Ⅱ类WRKY 蛋白,而其余2 个缺失了锌指结构。植物WRKY 转录因子家族存在锌指结构缺失或不完整的情况,如水稻第Ⅳ类WRKY 蛋白的锌指结构就出现缺失或不完整[25],而小豆WRKY 也存在N 端锌指结构整体缺失的情况[26]。
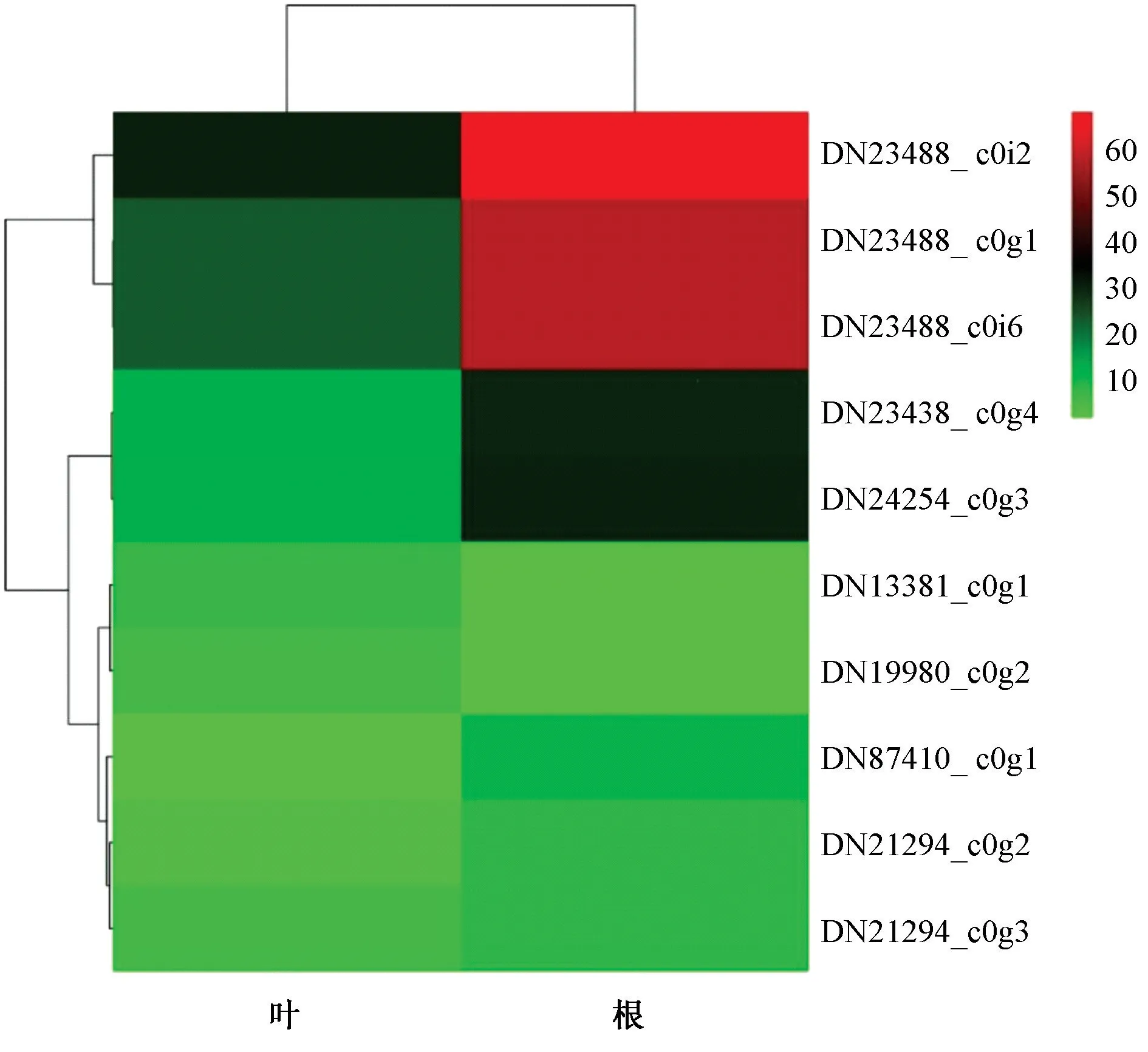
图5 金铁锁WRKY 基因在叶、根中的表达
聚类分析表明,DN19980_ c0g2、PgWRKY9 被聚为一类,说明这2 个WRKY 转录因子具有较近的亲缘关系及相似的功能。Xiu 等[27]从NGS 转录组中克隆了8 个WRKY 基因,命名为PgWRKY2-9,其中通过激素和NaCl 处理发现PgWRKY9 均有应答,表明它是胁迫应答转录因子,也提示DN19980_ c0g2 转录因子可能与金铁锁的抗逆性有关。PqWRKY1 (JF508376.1) 是从西洋参中克隆和鉴定得到的一个西洋参胁迫应答转录因子,参与其所含三萜皂苷合成的调控[28];PgWRKY1 (KU836935) 是人参的胁迫应答转录因子,通过调节相关酶基因的表达,调控其所含三萜皂苷的合成[13]。聚类分析显示,金铁锁DN23438_ c0g4 转录因子与PgWRKY1 转录因子、PqWRKY1 归为一类,表明它可能参与了其所含三萜皂苷的转录合成。
通过预测与分析转录因子结合位点,本实验寻找到3个报道于拟南芥的潜在TFBS 序列特征信息,motif 富集峰型图显示,MA 1093.1、MA1094.1、MA1087.1 的TFFM 峰型图都只存在1 个尖锐锋,只具有1 个motif 的结合位置,是否还有其他潜在的TFBS 序列和motif 结合位置尚不明确,需完成后续金铁锁全基因组测序后,调取相关金铁锁三萜皂苷合成途径关键酶基因的启动子来进行综合分析。表达模式分析发现,IIc、IId 亚型在金铁锁的根中有着较高的表达量,提示这两个亚型相似的表达模式可能与该药材生长发育及五环三萜皂苷在其根内的合成有关。
