粗茎秦艽种子萌发过程的转录组及关键因子分析
2021-01-05马子豪吕金盈王长生李爱暖
杨 晓,马子豪,马 婕,吕金盈,何 娟,王长生,李爱暖,陈 晨,曾 锐*
粗茎秦艽种子萌发过程的转录组及关键因子分析
杨 晓1,马子豪1,马 婕1,吕金盈1,何 娟1,王长生2,李爱暖1,陈 晨3*,曾 锐1*
1. 西南民族大学药学院,四川 成都 610041 2. 石柱土家族自治县农业农村委员会,重庆 409100 3. 西南民族大学青藏高原研究院,四川 成都 610041
研究粗茎秦艽种子萌发的转录组信息及影响因子。利用高通量测序技术测定粗茎秦艽种子萌发前、萌发中及萌发后3个阶段的转录组,并通过生物信息学技术进行分析。共获得了149 463条Unigenes,平均长度为601.88 bp,其中79 412个Unigene获得注释。6943个基因差异表达,大部分表现为萌发后上调表达,其中5188个富集到GO条目中,1815个富集到KEGG通路中,大多参与光反应过程、细胞壁合成、脂质代谢及次生代谢中。单碱基到六碱基核苷酸的SSR重复类型均有检出,SSR发生频率为15.13%,出现频率为18.81%,平均每3199 bp就含有1个SSR位点,SSR重复类型丰富,数目较多。粗茎秦艽种子萌发过程有大量调控基因参与,光照、激素为重要的调控因子,SSR专用标记的开发切实可靠,且为粗茎秦艽的次生代谢调控研究奠定了基础。
粗茎秦艽;种子;转录组;代谢通路;SSR标记;RNA-seq
秦艽性味辛、苦、平,归胃、肝、胆经,首载于《神农本草经》,列为中品,具祛风湿、清湿热、止痹痛和退虚热的功效[1],为我国常用的重要药材。粗茎秦艽Duthie ex Burk.为《中国药典》2015年版秦艽确定的基原之一,是我国目前栽培量最大的秦艽品种[2]。近年来临床用药量增加,过度采挖导致秦艽野生资源日渐枯竭[3],秦艽价格不断上涨迫切需要开展秦艽规模化人工栽培。粗茎秦艽以有性繁殖为主,其种子自然萌发发芽率低,萌发周期差异大[4],导致秦艽繁殖能力低,是困扰人工栽培的重要问题。近年来,主要用过温水催芽、赤霉素浸种[5],CO2激光处理[6],不同芽床[7],光照、水分等环境因子[8]等生理手段研究秦艽种子萌发的机制,其分子机制研究未见报道。
转录组为特定细胞或组织在功能状态或某一发育阶段下转录出来的所有RNA的集合[9]。高通量测序技术的迅速发展使得无参考基因组植物基因表达信息的获得变得越来越简便、快捷。近年来,转录组测序技术(RNA-Seq)越来越多的运用到种子萌发机制的研究中,如在西洋参[10]、黄芪[11]等种子萌发机制中的研究。本研究拟通过粗茎秦艽种子萌发前、中、后3个不同时期的转录组数据,初步分析粗茎秦艽种子萌发过程的相关基因、途径以及重要调控因子,为探索秦艽种子萌发的分子机制奠定基础。
1 材料与试剂
1.1 材料
试验所用粗茎秦艽种子于2016年采自四川省阿坝州黑水县沙石多乡甲足村(102°55′11.95″E,32°7′37.14″N,海拔2710 m)粗茎秦艽种植基地3年生成熟种子,经西南民族大学刘圆教授鉴定为龙胆科植物粗茎秦艽Duthie ex Burk.的成熟种子。
1.2 仪器与试剂
本实验借助Illumina HiSeq 2000测序平台,建库采用illumina原装试剂盒。1%琼脂糖凝胶电泳分析RNA降解程度以及是否有污染,Nanodrop 2000(Thermo Fisher公司,美国)检测RNA的纯度,Qubit 2.0(Invitrogen公司,美国)对RNA浓度进行精确定量,Agilent 2100(Agilent公司,美国)精确检测RNA的完整性,采用Estscan(3.0.3)软件预测其开放读码框,从而得到这部分基因编码的核酸序列和氨基酸序列,采用Primer3(2.3.5)进行简单重复序列(simple sequence repeats,SSRs)引物设计。
2 方法
2.1 种子的萌发
选取饱满种子于12 h光照,恒温25 ℃下萌发,分别取将萌发前(饱满的种子,未进行萌发,编号GC-MFQ-01、GC-MFQ-02、GC-MFQ-03)、萌发中(种子萌发至13 d,刚出现露白,编号GC-MFZ-01、GC-MFZ-02、GC-MFZ-03)、萌发后(从露白开始计时,用时5 d,胚根长至0.3~0.5 cm,编号GC-MFH-01、GC-MFH-02、GC-MFH-03)3个阶段(图1)。每个阶段取3个生物样品重复,迅速放入液氮中固定,−80 ℃冰箱储藏备用。
2.2 RNA抽提及转录组测序
样品总RNA提取按照改良的Trizol的方法进行[12],检测达到测序质量后构建文库。库检合格后,由北京诺禾致源科技股份有限公司Illumina HiSeqTM2000平台进行建库测序。
2.3 转录组分析
利用Trinity[13]对clean reads进行拼接,取每条序列中最长的转录本作为Unigene,以此进行后续的分析。基于BLAST[14]法取值<1e−5给出Unigene的蛋白数据库NR注释、SWISSPROT注释,蛋白相邻类的聚簇(KOG)功能注释、GO(gene ontology)分类和京都基因与基因组百科全书(KEGG)代谢通路分析。Unigene表达量的计算使用FPKM(fragments Per kb per Million reads)法[15]。

A-萌发前 B-萌发中 C-萌发后
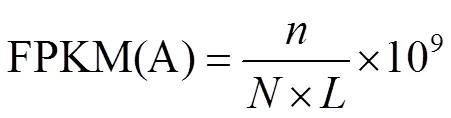
A为基因名,FPKM(A)为基因A的表达量,为比对到基因A的fragments数,为比对到所有基因的总fragments数,为基因的长度
利用MISA软件[16]对粗茎秦艽种子Unigenes进行SSRs检测,并进行统计分析。
3 结果与分析
3.1 转录组质量分析与Unigene拼接、注释
对粗茎秦艽种子的132 532 476条转录组raw data进行过滤,各样品的clean bases为5.99~6.80 Gb,Q30均在94.71%以上,GC含量平均值为43.71%(表1)。取每条基因中最长的转录本为Unigene,共获得149 463条Unigene,平均长度为601.88 bp,N50为909,长度在1 kb以上的有22 147条,表明组装结果较可靠。

表1 样品转录组clean data统计
raw data为原始测序序列;clean data为去除接头、无法确定碱基信息及低质量reads的序列数据;clean bases为测序序列的长度;Q30为碱基正确识别率达99.9%的序列比例;GC含量为碱基G和C占总的碱基数量的百分比
Raw Data was the original sequencing sequence; Clean Data was the sequence data with unconnected base information and low-quality reads; Clean bases was the length of the sequencing sequence; Q30 was the sequence ratio with a correct base recognition rate of 99.9%; GC content was the percentage of bases G and C to the total number of bases.
对获得的Unigene进行注释,共有79 412个Unigene获得注释信息,约占Unigene总数的53.13%(图2-B),与NR数据库比对结果见图2-A,其中,与咖啡Pierre ex Froehn.的同源性最高,达21.7%,其次为葡萄L. 为5.5%,油菜L. 为5.2%,芝麻L. 为4.3%,烟草L为3.0%,其余物种均小于3%。

图2 粗茎秦艽转录组NR数据库比对物种分布(A) 及注释统计 (B)
3.2 粗茎秦艽种子萌发过程中差异表达基因分析
差异基因共6943个。与粗茎秦艽种子未萌发的转录组对比,萌发中共有差异基因5960个,萌发后共有差异基因5619个,萌发后与萌发中共有差异基因587个,共同差异基因为386个。对共同差异表达的基因进行聚类分析,大部分差异基因表现为萌发中、萌发后上调表达。差异基因韦恩及聚类分析见图3。
3.3 差异表达基因的GO和KEGG富集分析
对粗茎秦艽种子萌发过程中的差异基因进行GO分类富集分析,结果表明,共有5188个差异基因富集到GO条目中,其中光合作用、淀粉代谢过程、蔗糖代谢过程为生物过程的优势富集条目,富集到的差异基因分别占总基因数的1.40%、1.35%和1.35%;光系统II和细胞壁为细胞组分的优势富集条目,富集到的差异基因分别占总基因数的0.94%和0.83%;CH-OH基团为供体,NAD/NADP为受体的氧化还原酶活性和醇类为受体的磷酸酶活性为分子功能的优势富集条目,富集到的差异基因分别占总基因数的17.15%和15.42%(图4)。对粗茎秦艽种子萌发过程中的差异基因进行KEGG富集分析,结果表明,共有1815个差异基因富集到KEGG通路中,其中淀粉和蔗糖的代谢(参与该通路的差异基因占比为5.29%)、植物激素信号转导(4.41%)、苯丙素类的生物合成(3.97%)、氨糖和核糖代谢(3.53%)、半胱氨酸和蛋氨酸代谢(2.81%)、光合作用的碳固定(2.75%)、脂肪酸代谢(2.70%)、戊糖-葡萄糖醛酸转换(2.37%)、光合作用(2.26%)和萜类化合物骨架的生物合成(1.82%)为KEGG富集途径的Top10(图5)。

图3 差异基因维恩(A)及聚类分析(B)

图4 差异基因的GO富集分析

图5 差异KEGG富集(top20)
3.4 种子萌发的差异代谢及调控过程分析
粗茎秦艽为环烯醚萜类、黄酮类等次生代谢产物含量丰富的药用植物,因此,对粗茎秦艽种子萌发过程中的代谢过程进行转录分析。较多的差异基因参与光反应过程、细胞壁合成、脂质代谢及次生代谢中(图6)。植物通过感受外界及自身的信号调控自身的发育,秦艽种子萌发过程中参与赤霉素、生长素、细胞分裂素等调控过程的差异基因数目较多,同时,大量的转录因子也参与了种子萌发过程的调控。通过以上统计分析,光照条件为秦艽种子萌发重要的外界条件,赤霉素、生长素、细胞分裂素等通过单独或联合调控萌发。
种子萌发过程中,含氮化合物、异戊二烯类、黄酮类等次生代谢过程中基因表达差异,统计结果见表2。转录因子调控着基因的表达,大量的转录因子在种子萌发过程中差异表达,统计发现,Auxin Response Factor家族、bHLH家族、Constans-like zinc finger家族、MYB-domain家族、WRKY家族、Aux/IAA家族、bZIP家族、NAC-domain家族为差异基因数目较多的转录因子家族,结果见表3。
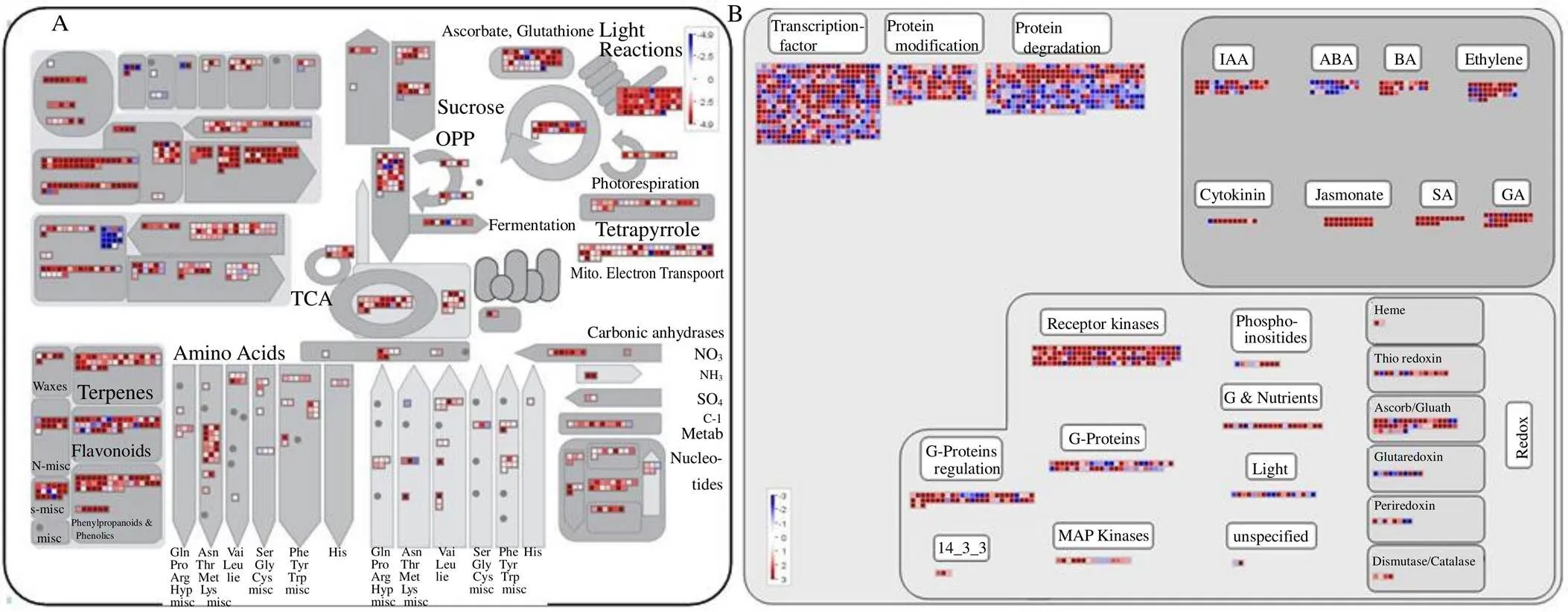
A-差异代谢富集 B-差异调控富集

表2 粗茎秦艽种子萌发过程次生代谢差异基因统计
3.5 SSRs特征分析
利用软件MISA对粗茎秦艽转录组149 463条Unigene序列进行SSR位点分析,单碱基到六碱基核苷酸重复类型均有检出。其中,28 120个SSR分布于22 613条Unigene序列中,SSR发生频率(含有SSR的Unigene数目与总Unigene的数目之比)为15.13%,出现频率(检出SSR个数与总Unigene数目之比)为18.81%,平均每3199 bp就含有1个SSR位点(图7)。其中18 167条Unigenes只包含单个SSR位点,4446条Unigenes包含1个以上SSR位点。粗茎秦艽SSR重复类型丰富,数目较多。
从SSR位点数量上看,出现最多的为一至三核苷酸重复,占到总SSR位点数量的98.45%。其中单核苷酸重复比例最高,可占到62.42%,A/T为优势重复类型,占单核苷酸重复类型的95.66%;其次为三核苷酸和双核苷酸重复,分别为19.46%和16.59%,AAG/GAA为三核苷酸优势重复类型,占三核苷酸重复类型的7.78%,AT/TA为二核苷酸优势重复类型,占二核苷酸重复类型的54.91%。四、五、六核苷酸重复类型的数量很少,总计占比1.55%(表4)。

表3 粗茎秦艽种子萌发过程差异转录因子统计

图7 粗茎秦艽种子转录组SSR重复类型统计
4 讨论
4.1 秦艽有效成分合成基因的筛选
粗茎秦艽的主要活性成分有环烯醚萜类、黄酮类、萜类及苯丙素类等[17-19],对粗茎秦艽种子萌发过程的转录组进行统计发现,大量的次生代谢相关基因差异表达,包括含氮化合物、异戊二烯类、黄酮类等,与陈俊可等[18]、王长生等[19]对于粗茎秦艽有效成分的鉴定结果一致。因此,在后续研究中,应对该部分基因进行克隆及功能验证,以期解析粗茎秦艽有效成分的基因合成路径,为粗茎秦艽有效成分的生物合成奠定基础。

表4 粗茎秦艽种子转录组的SSR位点分析
4.2 秦艽种子萌发水平的机制初探
植物激素是一类调节植物生理反应的活性物质[20]。赤霉素[21]、脱落酸[22]、乙烯[20]均可参与种子休眠的调控;生长素促进细胞的分裂与分化[23];细胞分裂素可调控芽的形成[24]。统计粗茎秦艽种子萌发过程差异表达基因,大量的生长素、赤霉素调控基因差异表达,因此,生长素、赤霉素在粗茎秦艽种子萌发过程中起重要作用。与侯格平等[5]使用了赤霉素浸种秦艽种子,提高了其萌发率相符合。另外,光反应过程、碳固定、淀粉和蔗糖代谢相关基因大量差异表达,表明光照应为粗茎秦艽种子萌发的重要条件。因此,后续试验应继续验证光照、植物内源激素对秦艽种子萌发的调控功效,以期提高粗茎秦艽种子萌发水平。
4.3 秦艽专属SSR标记的开发
SSR标记因其可重复性、多等位性、共显性遗传、相对丰度和良好的基因组覆盖率等特点,在植物遗传育种中广泛的应用[25]。利用转录组开发的SSR标记均位于编码区,因此,更有效地解释了不同品种的表型和功能的多样性,目前厚朴[26]、云锦杜鹃[27]等已利用转录组成功开发出有效的SSR标记。分析粗茎秦艽转录组的SSR特征,表明SSR类型丰富、数量可观,为粗茎秦艽遗传多样性研究及分子育种提供研究基础。
综上,研究粗茎秦艽种子萌发过程的转录组学,获得了大量的重要信息,后续将通过验证各个因子对于粗茎秦艽种子萌发的影响,揭示粗茎秦艽种子萌发的机制,探索提高其萌发率及成活率的技术手段。
利益冲突 所有作者均声明不存在利益冲突
[1] 中国药典 [S]. 一部. 2015, 270.
[2] 聂燕琼, 李海彦, 孙娜, 等. 粗茎秦艽资源研究进展 [J]. 中国现代中药, 2012, 14(5): 37-40.
[3] 郭江帆, 孙佳, 魏朔南. 秦艽种子内源萌发抑制物的初步探究 [J]. 种子, 2013, 32(7): 39-43.
[4] 滕红梅, 曹晓燕, 王喆之. 不同培养条件及预处理对秦艽种子萌发的影响 [J]. 种子, 2008, 27(11): 87-88.
[5] 侯格平, 姜青龙, 甄东升. 不同培养基质和物化处理对秦艽种子发芽成苗影响的研究 [J]. 农业科技通讯, 2014(2): 94-97.
[6] 薛慧君. UV-B辐射、CO2激光对秦艽生理、生长和有效成分的影响及其加工储藏方法的评价 [D]. 西安: 西北大学, 2004.
[7] 彭云霞. 发芽床对秦艽种子萌发特性的影响 [J]. 甘肃农业科技, 2014(12): 23-25.
[8] 侯茜, 胡锋, 张帆, 等. 环境因子对濒危药用植物秦艽种子萌发的影响 [J]. 中国现代中药, 2016, 18(2): 178-180.
[9] 齐楠, 卢娜. 植物化感作用机理及其在园林植物配置中的应用分析 [J]. 现代园艺, 2018(22): 111-112.
[10] Qi J, Sun P, Liao D,. Transcriptomic analysis of American ginseng seeds during the dormancy release process by RNA-Seq [J]., 2015, 10(3): e0118558.
[11] 于倩. 基于转录组对黄芪种子萌发过程及其内源激素分析 [D]. 呼和浩特: 内蒙古大学, 2016.
[12] Young M D, Wakefield M J, Smyth G K,. Gene ontology analysis for RNA-seq: Accounting for selection bias [J]., 2010, 11(2): R14.
[13] Grabherr M G, Haas B J, Yassour M,. Trinity: reconstructing a full-length transcriptome without a genome from RNA-Seq data [J]., 2011, 29(7): 644-652.
[14] Altschulsf, GishI W, Miller W,. Basic local alignment search tool [J]., 1990, 215(3): 403-410.
[15] Trapnell C, Williams B A, Pertea G,. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation [J]., 2010, 28(5): 511-515.
[16] Pertea G, Huang X, Liang F,. TIGR gene indices clustering tools (tgicl): a software system for fast clustering of large EST datasets [J]., 2003, 19(5): 651-652.
[17] Xu Y, Li Y, Maffucci K G,. Analytical methods of phytochemicals from the genus[J]., 2017, 22(12): 1-40.
[18] 陈俊可, 曾锐. 基于UPLC-ESI-HRMSn的代谢组学技术对粗茎秦艽不同部位化学成分研究 [J]. 中草药, 2018, 49(10): 2328-2335.
[19] 王长生, 董红娇, 包雅婷, 等. UPLC-Q-Exactive四级杆-静电场轨道阱高分辨质谱联用快速分析粗茎秦艽化学成分 [J]. 中草药, 2016, 47(18): 3175-3180.
[20] 王春阳. 七种主要植物激素信号通路的起源与进化 [D]. 泰安: 山东农业大学, 2013.
[21] Hartweck L M. Gibberellin signaling [J]., 2008, 229(1): 1-13.
[22] Miyakawa T, Fujita Y, Yamaguchi-shinozaki K,. Structure and function of abscisic acid receptors [J]., 2013, 18(5): 259-266.
[23] Davies P J. Plant hormones: biosynthesis, signal transduction, action [M]., 2010.
[24] Mok D W, Mok M C. Cytokinin metabolism and action [J]., 2001, 52(1): 89-118.
[25] Chen S, Luo H, Li Y,. 454 EST analysis detects genes putatively involved in ginsenoside biosynthesis in[J]., 2011, 30(9): 1593-1601.
[26] 代娇, 时小东, 顾雨熹, 等. 厚朴转录组SSR标记的开发及功能分析[J]. 中草药, 2017, 48(13): 2726-2732.
[27] 杨彬, 许蔷薇, 牛明月, 等. 云锦杜鹃转录组SSR分析及其分子标记开发 [J]. 核农学报, 2018, 32(12): 2335-2345.
Transcriptome and key factors analysis ofin seed germination process
YANG Xiao1, MA Zi-hao1, MA Jie1, LYU Jin-ying1, HE Juan1, WANG Chang-sheng2, LI Ai-nuan1, CHEN Chen3, ZENG Rui1
1. School of Pharmacy, Southwest Minzu University, Chengdu 610041, China 2. Shizhu Tujia Autonomous County Agriculture and Rural Committee, Chongqing 409100, China 3. Institute of Qinghai-tibetan Plateau, Southwest Minzu University, Chengdu 610041, China
To explore the transcriptome information and key factors ofin seed germination process.The transcriptome of the stage before germination, during germination and after germination ofwere determined by high-throughput sequencing and analyzed by systemic bioinformatics.A total of 149 463 unigenes were obtained, with an average length of 601.88 bp, and 79 412 unigenes were successfully annotated. A total of 6943 genes were differentially expressed, most of which were up-regulated after germination. Among them, 5188 genes were enriched in the GO and 1815 genes were enriched in the KEGG pathway, most of which were involved in the light reaction, cell wall synthesis, lipid metabolism and secondary metabolism. The 1—6 nucleotides SSR repeat types were all detected. The frequency of SSR occurrence was 15.13% and the frequency of appearing was 18.81%. On average, every 3199 bp contained one SSR site. Abundant SSR repeat types and a large number of SSR repeat types were obtained.A large number of genes were involved in regulating the germination process of. Light and plant hormones were important regulatory factors. The special SSR markers development was effective and reliable through transcriptome information. This study provided a reference for further understanding of regulation ofsecondary metabolism.
Duthie ex Burk.; seeds; transcriptome; metabolim pathway; SSR markers; RNA-seq
R282.12
A
0253 - 2670(2021)01 - 0219 - 08
10.7501/j.issn.0253-2670.2021.01.026
2020-06-06
国家重点研发计划(2017YFC1700705);四川省留学人员科技活动项目择优资助经费计划(2018-68);四川省重点研发项目(2019YFS0174);四川省科技支撑项目(14ZC2103);西南民族大学研究生创新型科研项目(CX2020SZ77)
杨 晓(1997—),硕士研究生,主要从事中药资源研究。Tel: 13258279138 E-mail: xiaoyeeyang@yeah.net
陈 晨(1987—),助理研究员,博士,主要从事药用植物发育分子机理及民族药资源研究。Tel: (028)83372832 E-mail: lilychenqhu@163.com
曾 锐(1976—),教授,主要从事中药和民族药研究。Tel: (028)85522099 E-mail: rzeng@swun.edu.cn
[责任编辑 时圣明]
