望天鹅保护区天然朝鲜崖柏种群年龄结构及更新特征
2021-01-05苑景淇李成宏王梅芳周梅妹兰雪涵于忠亮杜凤国
苑景淇,李成宏,高 纯,王梅芳,周梅妹,孙 华,兰雪涵,于忠亮,杜凤国,3
(1.北华大学林学院,吉林 吉林 132013;2.长白县林业局,吉林 白山 134400;3.长白山特色森林资源保育与高效利用国家林业局重点实验室,吉林 吉林 132013)
朝鲜崖柏(ThujakoraiensisNakai)为柏科(Cupressaceae)崖柏属(ThujaL)常绿乔木,国家二级重点保护植物,是极度濒危物种,具有珍贵观赏价值和重要经济用途[1].近年来,朝鲜崖柏受人类活动等外部因素干扰生境遭到破坏,加之受繁殖生物学内因(如更新方式)影响,种群分布及数量呈逐渐减少趋势.很多学者对朝鲜崖柏种群进行了研究,主要集中在繁育技术[2]、种实性状[3-4]、种群动态变化特征[5]等方面,而对种群年龄结构及天然更新的研究报道较少.天然更新是由于自然因素或人类活动造成植物损失或破坏后的再生长,是种群得以增殖、扩散和延续的一个重要生态过程[6];种群年龄结构反映种群内不同年龄个体在其生境中的数量分布状态,是揭示种群生存现状、更新程度及预测发展趋势的重要依据[7].因此,探究天然种群年龄结构及天然更新特征对于朝鲜崖柏种群恢复与重建具有重要意义.本文以望天鹅保护区天然朝鲜崖柏为对象,研究其种群年龄结构、更新方式,以及幼苗、幼树高生长规律以及土壤种子库特征,可为制定朝鲜崖柏保护措施提供理论依据.
1 材料与方法
1.1 研究地概况
研究地位于吉林省长白县望天鹅风景保护区,该区地势偏高.气候属温带大陆性季风气候,冬寒夏暖、四季分明,年平均气温3.1~3.7 ℃,年降水量600~700 mm,无霜期115~125 d.朝鲜崖柏为该保护区二级重点保护植物,分布地环境条件见表1.伴生乔木树种主要有臭松(Abiesnephrolepis)、花楷槭(Acerukurunduense)、假色槭(Acerpseudosieboldianum)、枫桦(Betulacostata)等;林下灌木有刺参(Oplopanaxelatus)、刺五加(Acanthopanaxsenticosus)、刺蔷薇(Rosaacicularis)、卫矛(Euonymusalatus)等;草本植物主要有石松(Lycopodiumjaponicum)、舞鹤草(Maianthemumbifolium)、细毛碗蕨(Oxaliscorniculata)、酢浆草(Oxaliscorniculata)等.

表1 研究地环境条件Tab.1 Environmental conditions of study site
1.2 调查方法
1.2.1 天然更新
1)样地调查.调查时间为2019年8月,调查地主要在朝鲜崖柏种群集中分布地区,沿垂直坡方向设置3块连续的10 m×30 m调查样地,面积共900 m2.每块样地均分成12个5 m×5 m的小样方,共36个.全面调查样地内朝鲜崖柏,胸径DBH≥3 cm的植株记录植株胸径、树高和冠幅;DBH<3 cm的植株记录地径和株高.
2)年龄结构确定.以径级结构代替年龄结构研究种群动态已得到广泛应用[8-9].本研究参照刘建峰[10]划分崖柏径级结构的方法,并结合朝鲜崖柏生长特性,确定8个径级:株高H<1 m(幼苗级)为第1径级,H≥1 m且DBH<3 cm(幼树级)为第2径级;DBH≥3 cm的个体,每级递增3 cm,如3 cm≤DBH<6 cm为第3径级,依次类推;15 cm≤DBH<18 cm为第7径级,DBH≥18 cm的个体为第8径级.
3)幼苗、幼树更新方式及高生长规律调查.在测量朝鲜崖柏幼苗、幼树地径和株高的同时鉴别更新方式:实生苗或萌生苗.随机选取30株幼苗、幼树,通过数轮枝和实测第1侧枝的方法确定年龄[11],探究植株高生长和年龄的关系.
1.2.2 土壤种子库
前期调查发现6株朝鲜崖柏有结实迹象(2018年球果宿存),选择其中3株生长状况相似、彼此间距较大(>20 m)的朝鲜崖柏结实树,在其周围采集土壤种子库,基本信息见表2.以结实树为中心,分别沿东、西、南、北4个方向取样,每个方向设置3个取样点,分别距离结实树树干1、3和5 m,随机取3个30 cm×30 cm土样.每个取样点分3层,分别为凋落物层、0~2 cm和2~4 cm土层,共108个分层样品.按形态完好种子、腐烂种子和空粒种子3类进行分层统计,并应用四唑法检测完好种子活力.分别于2019年8月种子成熟前和2019年10月种子成熟后采集种子.

表2 朝鲜崖柏结实树基本信息Tab.2 Basic information of Thuja koraiensis seed bearing tree
1.3 数据分析
应用Excel 2016和SPSS 19.0软件对数据进行统计分析.
2 结果与分析
2.1 朝鲜崖柏种群年龄结构
朝鲜崖柏径级见表3.由表3可知:900 m2样地共有411株朝鲜崖柏,其中,幼苗(290株)和幼树(56株)数量较多,分别占总量的70.56%和13.63%,两者总占比为84.19%;7、8(DBH≥15 cm)径级个体数量最少,共11株,仅占总株数的2.67%;3~6(3 cm≤DBH<15 cm)径级个体数量接近,合计54株,占总量的13.14%.从整体看,朝鲜崖柏种群径级连续,幼苗、幼树级占比较高,且具备一定数量的中径级植株,高径级个体数量最少,种群发展趋势为增长型.

表3 朝鲜崖柏种群径级结构Tab.3 Diameter class structure of Thuja koraiensis population /株
朝鲜崖柏植株生长状况见表4.由表4可见:在样地调查中发现34株枯死朝鲜崖柏,总死亡率为7.64%.其中,第4径级(6 cm≤DBH<9 cm)枯死植株数量最多,为13株,枯死率最高,为48.15%,主要原因可能是第4径级朝鲜崖柏高为5~10 m,受主林层乔木树冠遮挡,林下郁闭度较大形成被压木,因得不到充足的光照而逐渐枯死;7、8径级(DBH≥15 cm)朝鲜崖柏植株枯死率较高,分别为36.36%和33.33%,树干中空或腐烂是植株枯死的主要原因,并且两个径级存活的朝鲜崖柏植株都存在树干受损的情况,致使生长状况不良;幼苗总体上长势较好,偶见枯死植株,枯死率最低,为1.36%.
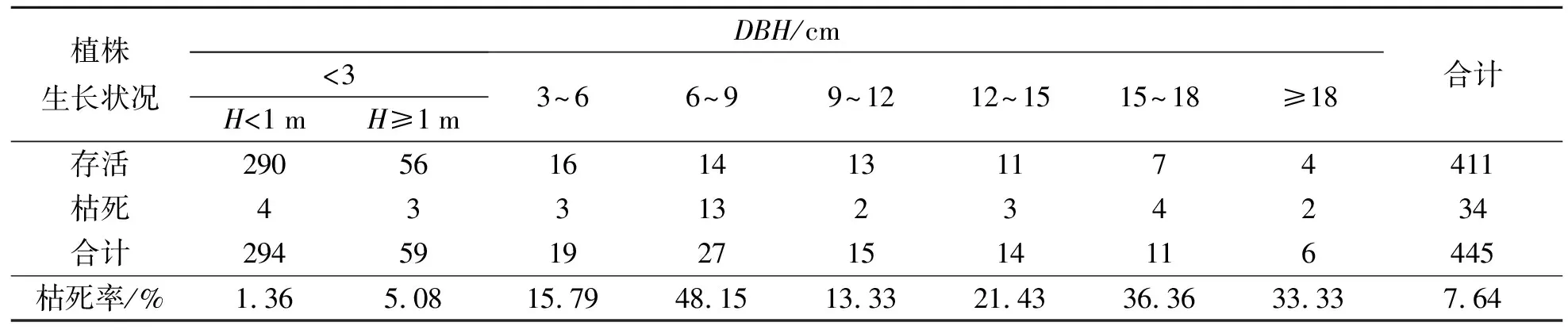
表4 朝鲜崖柏植株生长状况Tab.4 Growth of Thuja koraiensis /株
2.2 幼苗、幼树更新方式及特征
朝鲜崖柏幼苗平均地径0.97 cm,平均株高0.62 m;幼树平均地径1.93 cm,平均株高1.19 m,幼苗、幼树总更新密度为3 844株/hm2.调查发现,朝鲜崖柏种群内虽有结实树,但林下未见实生苗,幼苗、幼树均为萌生苗,且萌生方式有两种:一种是从植株根部萌发;另一种是侧枝与地面接触产生根系,逐渐形成新的植株[12].两种萌生方式形成的植株多呈匍匐状生长,根系间交错纵横,并且受较厚的凋落物和腐殖质影响,给实际调查中的精准区分带来了难度.
2.3 幼苗、幼树高生长规律
通过实测第1侧枝年轮和数轮枝数量的方式确定随机选取的30株萌生朝鲜崖柏幼苗、幼树年龄,并结合株高数据,选用常用的树木生长模型进行拟合[13].结果见表5.结果发现:朝鲜崖柏幼苗、幼树株高与年龄的回归模型拟合结果均为极显著水平.其中,三次回归曲线模型的决定系数最大(R2=0.905),拟合效果最好,最符合其生长趋势.由朝鲜崖柏幼苗、幼树株高与年龄的三次回归曲线模型(图1)可知:朝鲜崖柏幼苗、幼树株高随年龄的增加表现为快—慢—快的规律,具体为10 a以前增长速度较快,10~20 a增长速度减缓,20 a以后增长速度又逐渐加快.

表5 地径与年龄相关曲线拟合结果Tab.5 Fitting results of correlation curve between diameter and age
2.4 土壤种子库特征
由于望天鹅保护区朝鲜崖柏种群在2019年未结实,本文仅分析2019年8月采集的土壤样品中储存的朝鲜崖柏种子分布、数量及质量情况.
2.4.1 水平分布特征
朝鲜崖柏结实树周围不同水平方向、与结实树树干不同距离土壤中种子密度调查结果见表6.由表6可知:东、西、南、北4个方向上,土壤中朝鲜崖柏种子密度由大到小依次为南>东>西>北.经双因素方差分析可知:种子密度在水平方向上无显著差异(P=0.167>0.05).在距结实树1 m处,种子密度最大,且随着与结实树距离的增加,种子密度有减少的趋势,但差异不显著(P=0.051>0.05).

表6 土壤种子库密度水平分布Tab.6 Density and horizontal distribution of soil seed bank /(粒·m-2)
2.4.2 垂直分布特征
朝鲜崖柏土壤种子密度在不同土层深度的分布见表7.由表7可知:3株朝鲜崖柏结实树周围土壤中种子密度接近,没有显著差异(P=0.839>0.05).朝鲜崖柏种子主要分布在凋落物层,占比为70.18%;在0~2 cm土层中数量较少,占比为22.59%;在2~4 cm土层中数量最少,占比仅为7.23%.朝鲜崖柏土壤种子密度从凋落物层向土层逐步减小,且趋势明显,呈极显著差异(P=0.000<0.01).

表7 土壤种子库密度垂直分布Tab.7 Density and vertical distribution of soil seed bank
2.4.3 土壤种子质量
土壤种子库中种子的质量是影响种群更新潜力的主要因素.对不同层次(调落物层、0~2 cm土层、2~4 cm土层)种子数量和质量进行统计分析,结果见表8.由表8可知:在土壤分层样品中共发现996粒种子,其中腐烂种子数量最多(420粒),占种子总数的42.17%;空粒种子(404粒)数量次之,占种子总数的40.56%;完好种子(172粒)数量最少,仅占总数的17.27%.对完好种子进行TTC活力检测,结果表明,所有种子均没有活力.

表8 土壤中朝鲜崖柏种子质量Tab.8 Quality of Thuja koraiensis seeds in soil
3 结论与讨论
3.1 朝鲜崖柏种群年龄结构
望天鹅保护区天然朝鲜崖柏种群径级连续,小径级株数较多,随着径级升高株数逐渐下降;种群为增长型,总枯死率为7.64%.幼苗、幼树株数占比最高,为84.18%,枯死率最低,为2.02%;3~6径级(3 cm≤DBH<15 cm)中各径级株数接近,共占13.14%,其中第4径级(6 cm≤DBH<9 cm)植株因光照不足致使枯死率最高,为48.15%;7、8径级(DBH≥15 cm)朝鲜崖柏仅有11株,占比最低,为2.67%,且枯死率较高,为35.29%,植株生长状况不佳,表现为树干腐烂空心状况严重,甚至导致植株枯死.相比幼苗、幼树,中、高径级朝鲜崖柏受环境因素或自身生物学特性限制较大,致使数量更为稀少,濒危程度加剧.
3.2 朝鲜崖柏更新方式及幼苗、幼树高生长规律
在900 m2样地中未发现实生苗,幼苗、幼树以萌生方式更新,更新密度为3 844株/hm2.相比陈可贵[14]在1993年6 000株/hm2的调查结果减少了1/3,表明朝鲜崖柏更新密度下降严重.萌生幼苗、幼树的高生长随年龄增加呈快—慢—快的规律,但总体上高生长较缓慢.臭松是朝鲜崖柏的主要伴生树种,15 a和25 a生臭松株高分别约2.8 m和3.9 m[15],而同龄的朝鲜崖柏株高约0.8 m和1.2 m,在与臭松争夺光资源中不占优势.相比前人的调查,本次调查结果中朝鲜崖柏伴生的乔木、灌木种类及数量有所增加,致使朝鲜崖柏个体面临的资源竞争和环境阻力随之增大,加之自身生长缓慢,阻碍了朝鲜崖柏幼苗、幼树的生长发育.
3.3 朝鲜崖柏土壤种子库分布特征
朝鲜崖柏土壤种子库中,在水平分布上呈现东面(104粒/m2)和南面(114粒/m2)种子密度略大于西面(101粒/m2)和北面(91粒/m2)种子密度,随着与结实树树干距离的增加种子密度减小,但均无显著差异,产生这种现象的原因可能与坡向和风向有关.望天鹅保护区秋、冬季节主要为西风和西北风,调查的结实树地处东坡下坡位,属于背风坡,因此东面和南面的种子密度略大,并且朝鲜崖柏结实树所处林分郁闭度较大,调查的结实树未进入主林层,在主林层乔木和周围植株的庇荫下,林内风力较弱.种子在重力作用下更多地落在结实树树冠下,因此种子密度表现为1 m(114粒/m2)>3 m(101粒/m2)>5 m(93粒/m2),这与有种翅的长白落叶松种子近结实树分布的特点相似[16].在垂直分布上,种子密度表现为凋落物层(72粒/m2)>0~2 cm土层(23粒/m2)>2~4 cm土层(7粒/m2),不同层次间种子密度逐级递减,且呈极显著差异,该结论与部分研究者[16-18]发现的翅状种子在土壤垂直层的分布规律结果一致.
3.4 朝鲜崖柏土壤种子库质量
朝鲜崖柏土壤种子库平均种子密度为102粒/m2,种子转化成苗木的转化率为0.与其他裸子植物的土壤中种子密度(如东北红豆杉829粒/m2 [19],兴安落叶松1 048~2 069粒/m2 [17])相比,朝鲜崖柏种子库种子储备量较少.朝鲜崖柏种子质量极差,表现为发霉种子(42.2%)和空粒种子(40.6%)占比较高,形态完好种子(17.2%)占比较低,并且种子没有活力,属于瞬时种子库[6],这可能是朝鲜崖柏无实生苗的主要原因.有学者研究表明,林内湿度大易使种子发霉而丧失活力[20];小粒种子受枯落物层机械阻碍,无法与土壤接触而死亡[21];土壤中温度、湿度未满足种子萌发条件,随时间推移,种子生命力逐渐减弱[22].除朝鲜崖柏种子自身空粒多、成熟度差外,环境因素也可能是影响种子活力的原因.
土壤种子无活力和种群未见实生苗反映出朝鲜崖柏存在严重的实生更新障碍,因此,应开展朝鲜崖柏结实规律及土壤中种子寿命的相关研究,以便更好地保育朝鲜崖柏.
