热应激对荷斯坦奶牛泌乳能力和乳成分乳脂脂肪酸组成的影响
2020-12-09李征陈胜楠葛萃萃郝欣王海鉴张惠君
李征,陈胜楠,葛萃萃,郝欣,王海鉴,张惠君
(1.广州风行乳业股份有限公司,广州510510;2. 浙江公正检验中心有限公司,杭州310009;3. 北京海关技术中心,北京100026;4. 安捷伦科技(中国)有限公司,北京100102;5.中国检验认证集团澳洲分公司, 悉尼-墨尔本)
0 引 言
热应激是指机体对不利于其正常生理活动的高温环境所产生的一系列非特异免疫应答反应的总和[1]。奶牛热应激是指当奶牛受到高温刺激,作用于下丘脑-垂体-肾上腺皮质系统所引起的机体非特异性防御反应和特异性障碍在内的全身性适应症[2]。
热应激使奶牛的新陈代谢发生紊乱、采食量和产奶量下降、乳成分改变、免疫力降低等。近年来,随着奶牛养殖集约化的快速发展及全球气温变暖,导致热应激对奶牛生产性能降低、繁殖率下降等现象日趋严重,并成为阻碍我国奶业发展的最活跃因素之一。
关于热应激对奶牛的影响已有不少报道,但主要集中于评定奶牛耐热性指标的研究。本研究通过对比热应激与非热应激奶牛的泌乳能力和乳脂脂肪酸的变化,为更全面的了解热应激对奶牛的影响以及制定相应的营养调控措施提供理论依据。
1 实 验
1.1 奶牛选择与分组
选择体重相近、产奶量相近、同胎次、健康的均处于泌乳中期的荷斯坦奶牛24 头作为阴性对照组,集中进行第1 次采样。在热应激比较严重的8 月上旬再挑选体重相近、产奶量相近、同胎次、健康的均处于泌乳中期的荷斯坦奶牛24 头作为阳性对照组,进行第2 次采样。2组试验奶牛的饲养水平与管理方法一致。
1.2 实验日粮
实验奶牛的日粮组成参照NRC(2001)配制。基础日粮由青贮玉米、苜蓿干草、精料补充料组成,粗精比为49︰51,日粮配方及营养水平如表1所示。
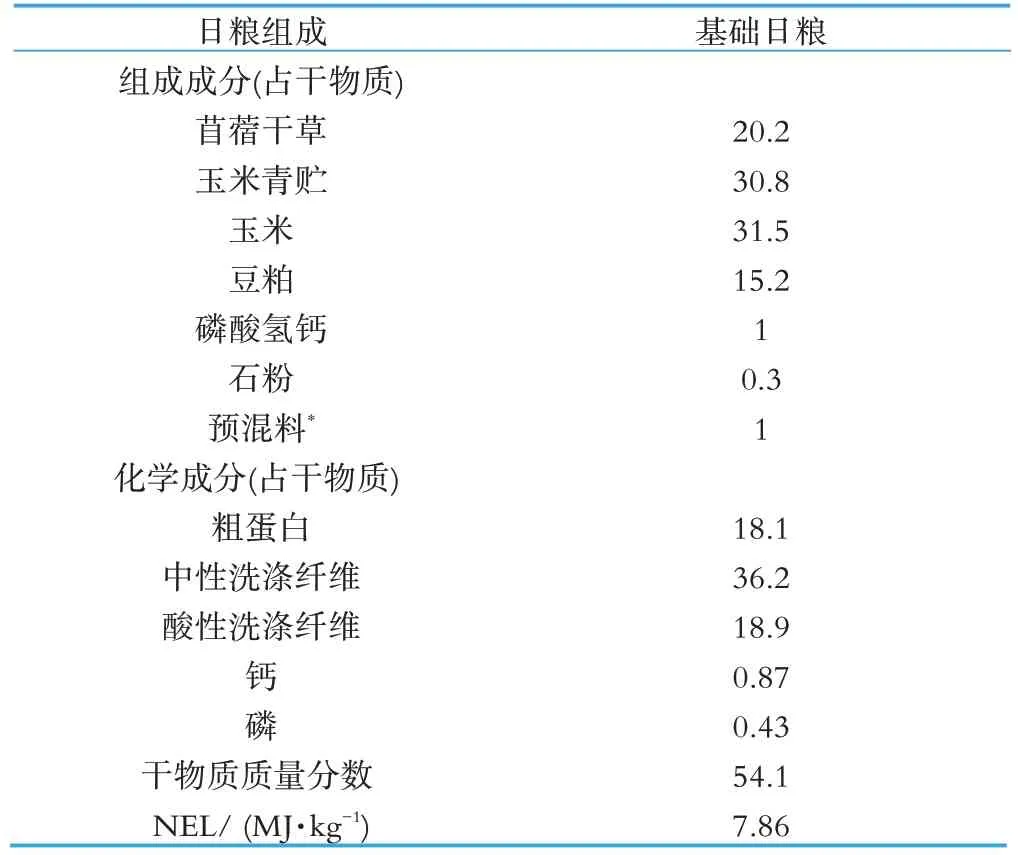
表1 日粮组成及营养成分 %
1.3 样品的采集及测定指标
在采样日的上、下午连续取奶样按1∶1 原则混合后装于2 个奶样瓶。一个奶样立即用美国Bentley 2000 型红外线牛奶成分分析仪测定乳成分(包括乳脂率、乳蛋白率和乳糖率);另一个奶样在-20 ℃冷冻保存,等试验结束后测定脂肪酸。并记录2 组试验奶牛在采样日的产奶量。
1.4 脂肪酸的分析
脂肪酸的分析参照HJ Zhang[3-4]的气相色谱法。
1.4.1 仪器与试剂
安捷伦公司5977B 气质联用:色谱柱为HP-88(Agilent)毛细管柱(100 m×0.25 mm,液膜厚度0.25 μm);载气为高纯氦气,载气流速为1 mL/min;离子源为EI源;电离电压70 eV,质荷比范围为29~500。脂肪酸甲酯标品为Matreya公司产品。
1.4.2 色谱条件
进样口和离子源温度为230 ℃。采取程序升温的方法,升温条件为:初温70 ℃,保持2 min,以8 ℃/min的速率升温至120 ℃,再以15 ℃/min 的速率升温至160 ℃,保持40 min,然后以5 ℃/min升温至215 ℃,保持10 min。进样量为1.0 μL,分流比为40∶1。
1.4.3 样品的提取和甲酯化
乳脂脂肪酸的提取根据Hara 和Radin[5]的方法,再按照张惠君[6]的方法对乳脂中的脂肪酸进行甲酯化,具体操作如下。
首 先配 CH2Cl∶2CH3OH(体 积 比 为=2∶1)溶液 ,5 mL 奶样+5 mL 溶液,混匀后超声 20 min,然后 4 000 r/min,倒去上层乳液,吸取下层CH2Cl2溶液至离心管,待CH2Cl2溶液挥干,然后甲酯化。
取约 0.45 g(50 uL)乳脂,加 2 mL 正己烷、70 uL 醋酸甲酯和 1 mol/L 的 KOH/CH3OH 溶液 70 uL,室温下放置10 min,再加入35 uL 草酸甲醇溶液作反应终止剂。然后取混合溶液的上清液1 mL,冷藏以备气质进样。
1.5 数据处理
用SPSS 13.0版软件进行单因素方差分析处理,采用F检验。
2 结 果
2.1 热应激对产奶量和乳成分的影响
由表2 可见,夏季热应激奶牛的产奶量与非热应激奶牛存在显著差异,产奶量在热应激时期极显著降低(P<0.01)。热应激时牛奶中的乳成分也会发生明显的变化,热应激奶牛的乳脂率极显著低于非热应激奶牛(P<0.01),乳蛋白率也显著下降(P<0.05),乳糖率有降低的趋势,但差异不显著(P>0.05)。
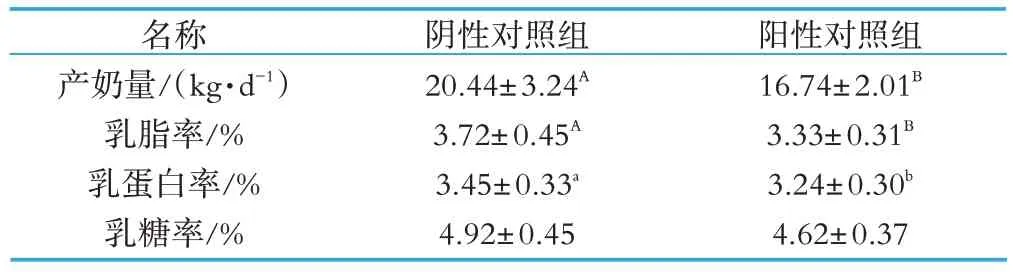
表2 热应激对奶牛产奶量和乳成分的影响
2.2 热应激对乳脂脂肪酸的影响
试验奶牛乳脂脂肪酸的组成见表3。热应激对乳脂中单个脂肪酸质量分数的影响不是很大,只有C18∶1脂肪酸的质量分数在热应激时期显著下降(P<0.05)。但总饱和脂肪酸(SFA)和总不饱和脂肪酸(UFA)的质量分数在热应激时期有明显的变化。热应激奶牛的总饱和脂肪酸质量分数显著高于非热应激奶牛(P<0.05),而总不饱和脂肪酸质量分数显著低于非热应激奶牛(P<0.05)。
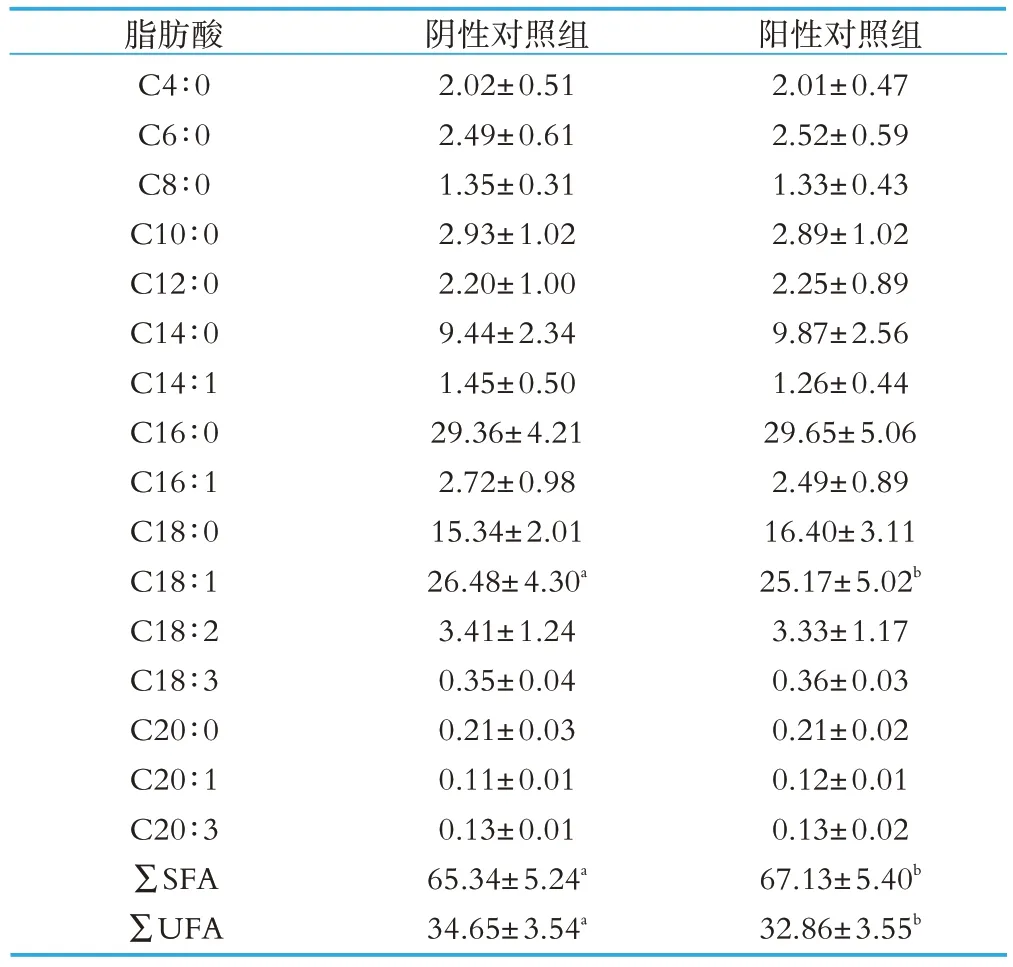
表3 热应激与非热应激奶牛乳脂脂肪酸的组成g(每100g脂肪酸中)
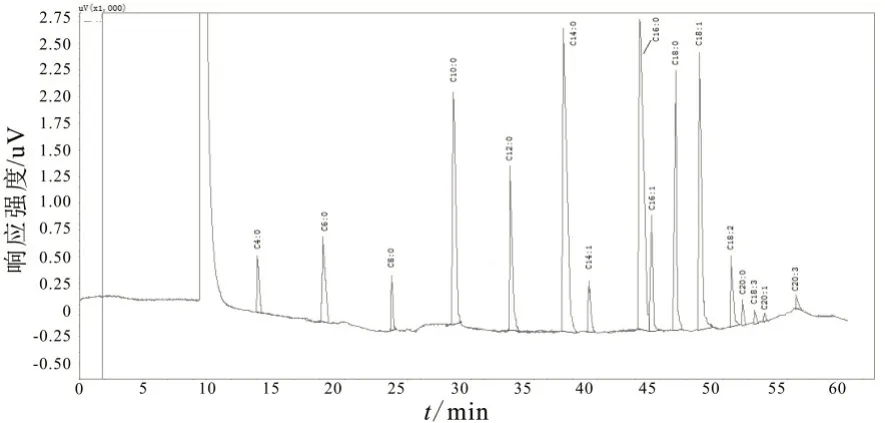
图1 热应激条件下奶牛乳脂脂肪酸气相色谱图(阳性)
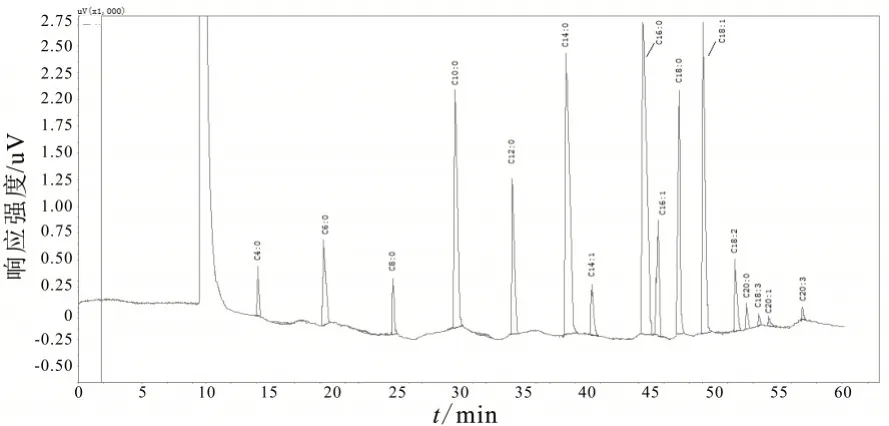
图2 非热应激条件下奶牛乳脂脂肪酸气相色谱图(阴性)
3 讨 论
3.1 热应激对产奶量和乳成分的影响
奶牛对热应激的反应首先表现为食欲降低,采食量减少。热应激引起奶牛采食量的减少必然导致奶牛产奶量的降低。当环境温度在4~21 ℃之间,奶牛的产奶量较高[7],当环境温度超过24 ℃时,奶牛的产奶量开始下降。王前等[8]报道,温度和产奶量之间呈强负相关。李建国等[9]研究表明,热应激能显著降低奶牛各泌乳阶段及泌乳期的平均泌乳量。温度达到40 ℃时,耐热性差的奶牛拒绝采食[10]。本试验中,热应激奶牛的产奶量极显著低于非热应激奶牛,与上述文献报道结果一致。热应激引起奶牛产奶量的降低主要有两个原因,一方面是采食量的下降减少了营养成分的消化和代谢,从而用于乳汁合成的营养成分也相应减少;另一方面是热应激造成奶牛生殖内分泌出现抑制反应,使孕激素、雌激素和促乳素等分泌量下降,抑制了乳腺泡、乳腺管的形成与发育,抑制了排乳反应,从而造成产奶量的下降。
热应激除了降低奶牛的产奶量,还会影响牛奶中乳成分的组成。乳脂、乳蛋白和乳糖均可因高温而下降[7],其中乳脂率在一年四季中的变化较大,以夏季最低。McDowell 等研究[11]表明当气温由18 ℃上升到30 ℃时,乳脂率和乳蛋白分别下降了39.7%和16.9% 。本试验中,奶牛的乳脂率和乳蛋白率在热应激时均显著降低。热应激使乳脂率降低主要是因为奶牛在热应激时喜食精料,从而使精料的采食量相对增多,而纤维摄取量下降,以致瘤胃内低级脂肪酸如乙酸、丙酸等的比例缩少,而乙酸和丙酸是合成乳脂的前体物质,于是奶牛的乳脂率降低。本研究中热应激奶牛的乳蛋白率显著下降可能是因为奶牛的采食量下降造成蛋白质的摄取量下降,而热应激时代谢率增加,加速了蛋白质的降解,同时皮肤蒸发使氮的排出增加,从而使机体内的蛋白质减少,引起乳蛋白率的下降。
3.2 热应激对乳脂脂肪酸的影响
乳脂大约含有质量分数为70%的饱和脂肪酸(SFA),27%的单不饱和脂肪酸(MUFA)和3%的多不饱和脂肪酸(PUFA)。由于不饱和脂肪酸,尤其是多不饱和脂肪酸结构中的双键,所以容易被氧化。研究表明动物处于应激状态时,代谢机能紊乱,交感-肾上腺系统反应增强,导致肾上腺髓质部大量分泌儿茶酚胺类激素而使氧自由基增加。在正常情况下,体内氧自由基的产生、利用和清除三者之间处于动态平衡。而在热应激状态下,机体产生过多的氧自由基不能及时清除,会导致体内氧自由基过剩,从而引起脂质过氧化作用,破坏生物膜的结构[12]。因此在热应激时期,乳脂中的不饱和脂肪酸可能被氧化,从而减少乳脂中它们的质量分数,而相应的增加饱和脂肪酸的质量分数。本研究中,总饱和脂肪酸质量分数在热应激时期显著升高,总不饱和脂肪酸质量分数显著降低。而在动物日粮中添加抗氧化剂也可以保护不饱和脂肪酸不被氧化,从而增加组织或牛奶中不饱和脂肪酸的质量分数,而减少饱和脂肪酸的质量分数。Barja 等[13]报道在猪日粮中添加抗氧化剂维生素E,由于维生素E对多不饱和脂肪酸的过氧化反应的保护作用,从而增加了猪肝脏中多不饱和脂肪酸的质量分数。在奶牛日粮中添加高剂量的抗氧化剂硒和维生素E 也可以增加牛奶中总不饱和脂肪酸的质量分数,而降低总饱和脂肪酸的质量分数[14]。另外在人类中,Dodge 等[15]报道硒可以提高母乳中多不饱和脂肪酸的质量分数(41%),而降低饱和脂肪酸的质量分数(11%)。
4 结 论
热应激使奶牛的产奶量、乳脂率和乳蛋白率降低,但对乳糖率没有影响。热应激使奶牛乳脂中的总饱和脂肪酸质量分数升高,而总不饱和脂肪酸质量分数降低。
