牛乳中嗜冷菌及其耐热酶研究进展
2020-12-09张树飞薛玉玲崔玥韩俊华朱宏王世杰
张树飞,薛玉玲,崔玥,韩俊华,朱宏,王世杰,
(1.河北科技大学生物科学与工程学院,石家庄 050018;2.石家庄君乐宝乳业有限公司,石家庄 050221)
0 引 言
在20 ℃以下能生长繁殖,最适生长温度为10~15 ℃的微生物定义为嗜冷菌[1]。嗜冷菌是引起牛乳质量问题的主要微生物之一。通常,在挤奶、储存和运输过程中容易发生嗜冷菌污染[2],从而导致产品质量问题。在低温存储过程中,嗜冷菌仍然能够生长繁殖[3],并且能够产生蛋白酶和脂肪酶等多种水解酶类[4],释放到牛乳中。当采用加热杀菌时,嗜冷菌菌体较容易杀灭[5],而其产生的酶类具有热稳定性[6],仍然能够保持一定的活性。脂肪酶和蛋白酶是影响牛乳品质的主要腐败酶[7],这些酶在牛乳中长时间作用,会破坏其中的营养物质,影响牛乳的品质。
1 牛乳中的嗜冷菌
1.1 种类
牛乳中嗜冷菌的主要菌属有假单胞菌属(Pseudomonas)、气单胞菌属(Aeromonas)、不动杆菌属(Acinetobacter)、沙雷氏菌属(Serratia)、金黄杆菌属(Chryseobacterium)、产碱杆菌属(Alcaligenes)、黄杆菌属(Flavobacterium)、乳杆菌属(Lactobacillus)、乳球菌属(Lactococcus)、芽孢杆菌属(Bacillus)、链球菌属(Streptococcus)等[8]。有研究报道,来自某农场和奶罐的20 个不同的原奶样品中共鉴定出2 906 株嗜冷菌,其属于169 个种和61 个属。其中,假单胞菌属(Pseudomonas)、乳球菌属(Lactococcus)和不动杆菌属(Acinetobacter)是最丰富的属,占所有菌株的62%。在所有分离株中,有18%属于迄今未知的物种,这表明部分牛乳微生物区系尚未开发[9]。
1.2 产生的蛋白酶和脂肪酶
牛乳中的嗜冷菌具有产生腐败酶的能力,主要有蛋白酶和脂肪酶[10]。其产生的蛋白酶残留在牛乳中,缓慢作用于牛乳蛋白,分解α-酪蛋白和κ-酪蛋白[11],释放出带苦味的氨基酸和短肽,导致产品出现苦味、涩味等非正常风味[12];也会导致酪蛋白胶束的ζ电势降低[13],使UHT 乳在贮存过程中出现蛋白质凝胶和乳清析出现象[11]。嗜冷菌也是牛乳中微生物脂肪酶的主要来源,其产生的脂肪酶主要是胞外酶[14]。脂肪酶能水解脂肪为甘油和游离脂肪酸[15],导致产品出现脂肪氧化味、腐臭等异味[16],同时会导致脂肪上浮速度加快,进而影响牛乳的品质。
不同的嗜冷菌产生蛋白酶和脂肪酶的能力有所差异。假单胞菌属(Pseudomonas)、沙雷氏菌属(Serratia)和金黄杆菌属(Chyseobacterium)的一些菌种产生蛋白酶的能力较强,而一些不动杆菌属(Acinetobacter)的菌种具有较强的脂肪酶产生能力[17]。牛乳中产生蛋白酶和脂肪酶的主要嗜冷菌菌属和菌种见表1。

表1 产生蛋白酶和脂肪酶的主要菌属和菌种
2 嗜冷菌产生的酶的热稳定性
2.1 蛋白酶
嗜冷菌产生的蛋白酶具有热稳定性,很难通过巴氏杀菌、高温瞬时灭菌等常规热处理技术使其失活[23-24]。研究表明,荧光假单胞菌07A(Pseudomonas fluorescens 07A)分泌的胞外蛋白酶具有477 个氨基酸以及高度保守的Ca2+和Zn2+结合域,属于金属蛋白酶,在72 °C 下处理15 s 后保留31.7%±0.37%的活性,而经过65 °C、30 min 处理后保留14.3%±0.37%的活性,即使分别在 80 °C、90 °C 和100 °C 下处理 5 min,该蛋白酶仍保留其初始活性的44.1%±1.10%、43.6%±0.58%和40.5%±1.73%[25]。在 UHT 脱脂牛乳、半脱脂牛乳、全脂牛乳3 种产品中,液化沙雷氏菌L53(Serratia liquefaciens L53)分泌的蛋白酶 Ser2 在 140 °C 下处理 4 s 的残留活性在 90%~95%之间,65 °C 处理 30 min 后的残留活性在87%~95%之间,在72 °C 热处理20 s 后,未观察到对蛋白酶活性的影响[11]。近几年研究报道的牛乳中嗜冷菌分泌的蛋白酶的热稳定性见表2。
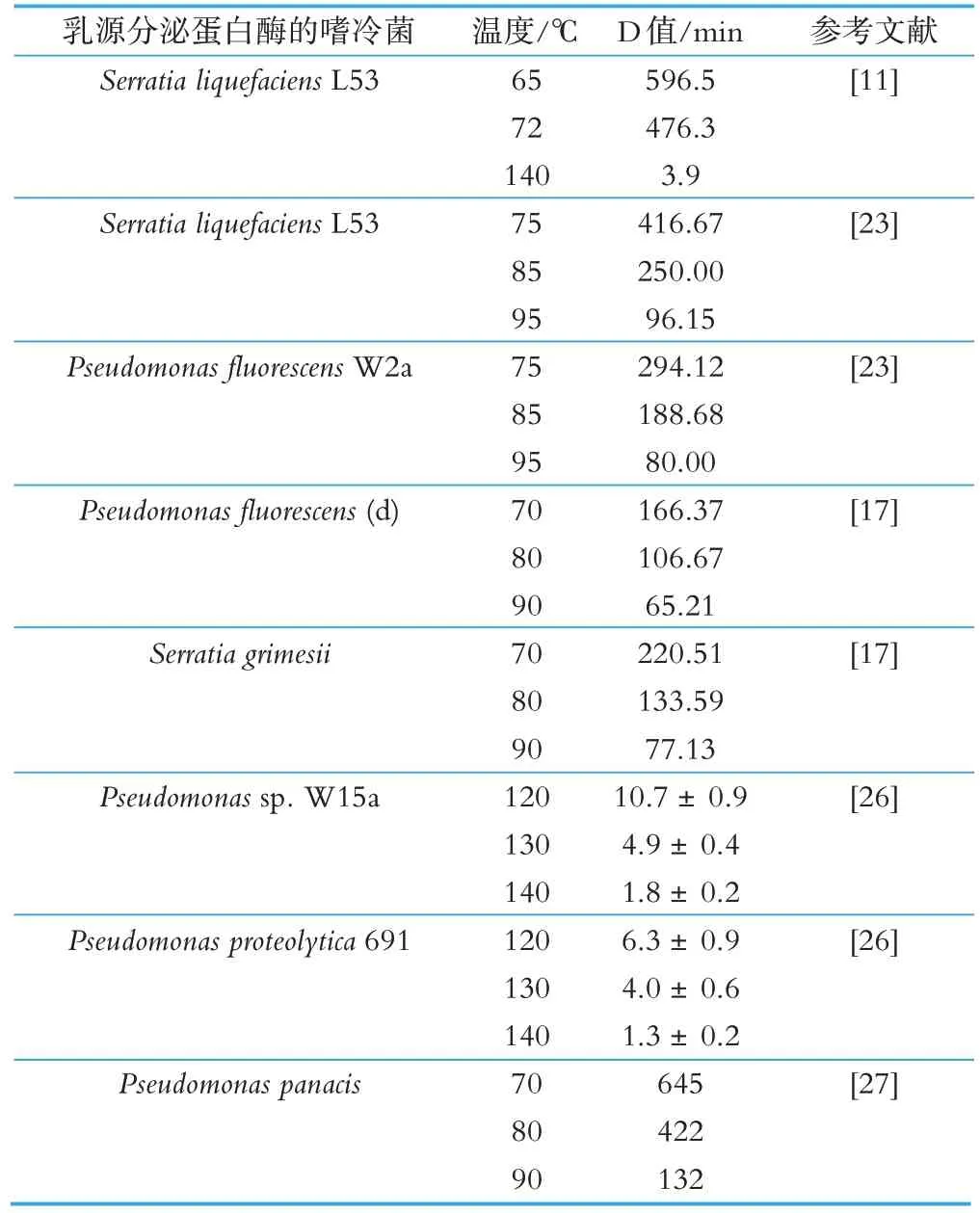
表2 乳源嗜冷菌分泌的蛋白酶的热稳定性
2.2 脂肪酶
脂肪酶具有较强的耐热性,在经过较高温度处理后依然能够保持一定的活性[28]。石璞洁等[29]报道了不同的温度时间处理对荧光假单胞菌脂肪酶活性的影响。研究发现,经50 ℃、30 min,63 ℃、30 min,72 ℃、20 s处理后,残留的脂肪酶相对酶活力分别为75.02%、86.21%、73.25%;90 ℃ 、10 min,121 ℃ 、0.1 MPa、20 min 处理后,残留的脂肪酶相对酶活力分别为17.62%和13.63%。近几年报道的牛乳中嗜冷菌分泌的脂肪酶的热稳定性见表3。
3 嗜冷菌及其酶的控制措施
避免由嗜冷菌及其酶引起的牛乳质量问题,主要还是通过对嗜冷菌进行控制。当前策略主要包括保持奶站和牧场良好的卫生环境[34-37]、使用生物防腐剂抑制嗜冷菌的生长[38]、CO2和 N2处理[39-43]、高压均质和微滤处理[44-46]。
3.1 保持奶站和牧场良好的卫生环境
嗜冷菌通常散布在卫生环境较差的奶牛场中。牛乳中嗜冷菌的数量和类型受设备清洁度、饲料、动物健康状况以及周边环境等因素的影响[34]。奶站及牧场应为奶牛提供整洁的场所,并对奶牛的生产环境进行严格的控制,对于挤奶设备需要及时的消毒,最大限度的减小嗜冷菌感染的风险[35-36]。褚立群[36]提出了通过及时清理牛床上的粪便、增添干净垫料、每天刷洗牛体、保持牛身清洁等方法来控制奶牛的生产环境,以降低嗜冷菌的感染风险。Bradley[37]等人报道了挤奶前对乳头进行消毒并干燥处理,可减少牛乳中的链球菌、肠球菌的数量;当不同奶牛进行挤奶时,需要进行集体消毒,这与牛乳中嗜冷菌数量的减少有关。

表3 乳源嗜冷菌分泌的脂肪酶的热稳定性
3.2 添加生物防腐剂
目前对于生物防腐剂的研究主要是噬菌体对于嗜冷菌的抑制作用。噬菌体能够感染嗜冷菌细胞,裂解嗜冷菌细胞以释放它们的后代,然后重复感染和裂解这个循环,直到嗜冷菌种群严重减少[5],随着嗜冷菌数量的降低,其分泌到牛乳中的耐热性蛋白酶和脂肪酶也随之减少,对于牛乳的危害减小。Hu 等[38]人在2016 年的研究中报道了,在含有嗜冷假单胞菌的UHT 牛乳中添加噬菌体混合物可使假单胞菌的细胞数显著降低2 log CFU/mL;当在原料乳中添加噬菌体混合物时,无论是在4 ℃下还是在25 ℃下储存,嗜冷菌的数量均减少了1 log CFU/mL;原料乳在4 ℃下冷藏5 d 后,经过噬菌体处理的原料乳比未经噬菌体处理的样品中的嗜冷菌数量低了4 log CFU/mL。该研究表明了噬菌体对于牛乳中嗜冷菌的显著抑制作用。Tanaka 等[5]报道了用噬菌体HU1 处理的脱脂牛乳和全脂牛乳比未处理样品中的假单胞菌活细胞数降低了 1 000 倍,同时,在 63 ℃ 加热 30 min 后,噬菌体的活性降低到不可检测的水平,显著提高了产品的安全性。因此,使用噬菌体处理牛乳可能是减少牛乳中嗜冷菌数量、降低耐热酶对于牛乳的危害的有效方法。
3.3 CO2和 N2处理
CO2已经成为一种有潜力的原料乳的防腐剂,它可以抑制嗜冷菌的生长,降低嗜冷菌合成耐热性蛋白酶和脂肪酶的速率,且其方法为非热处理,对牛乳的营养成分不会产生不利影响,因此被公认为是安全的[39]。向牛乳中充入适量的CO2,CO2与嗜冷菌细胞质中的水发生水合反应会生成碳酸使pH 降低,且CO2会取代O2,会影响嗜冷菌正常的代谢[40]。研究表明,经过CO2处理的原料乳出现变质的时间至少可以推迟7 d[39],且使用经CO2处理的原料乳生产的UHT牛乳在储存120 d 期间呈现较低的蛋白水解和脂解作用[41]。因此,用CO2处理原料乳将有利于改善UHT 乳的质量,降低牛乳中蛋白酶的蛋白水解和脂肪酶的脂解作用。
也有研究表明,用纯净的N2连续冲洗原料乳能够显著抑制嗜冷菌的生长,减少蛋白酶和脂肪酶的产生,降低冷藏期间蛋白水解和脂解作用,延长原料乳的冷藏时间[42-43]。
3.4 高压均质和微滤处理
高压均质是乳制品加工中常用的处理技术。有研究表明,高压均质在控制蛋白酶引起的蛋白质水解方面效果显著。Oliveira 等[44]研究发现,脂肪含量3%的牛乳经100 MPa 和150 MPa 压力处理后,蛋白水解分别显著降低了29%和51%,产品出现凝胶的时间分别延迟了23 d 和40 d。在UHT 处理前和处理后,分别经250 MPa 和400 MPa 双均质化处理,也成功地防止了UHT 牛乳由于脂肪酶造成的脂肪分解,显著改善了其贮存期[45]。
微滤可延长原料乳的保质期和改善UHT 牛乳质量。在高体细胞数和低体细胞数的原料乳中,微滤处理使牛乳菌落总数显著降低了3.56 和3.57 log CFU/mL,且分别加工成UHT 乳后,其货架期有效延长了63 d 和21 d[46]。微滤还可以延迟UHT 牛乳中凝胶和沉淀物的形成,并减少蛋白水解[45]。
4 结 语
牛乳中嗜冷菌的种类繁多,目前已有部分确定了其菌属,还有一部分尚未发掘,且对于嗜冷菌的嗜冷机制鲜有报道;另外,嗜冷菌产生的蛋白酶和脂肪酶具有热稳定性,会对牛乳造成破坏,然而对于其产生的酶类的耐热机制和酶的具体种类很少有研究,还需要进一步的探索。对于嗜冷菌及其酶的控制,最主要的还是从源头对嗜冷菌进行控制,以减少耐热酶的产生。因此,发掘天然、无毒害的抑菌物质也是当今的重要任务,还需要进一步的探索研究。
