不同来源的植物乳杆菌基因组和抗生素抗性
2020-11-11殷瑞敏李晓姝毛丙永崔树茂赵建新
殷瑞敏, 李晓姝, 毛丙永, 崔树茂, 赵建新
(江南大学 食品学院,江苏 无锡214122)
植物乳杆菌是一种极其多样化的乳杆菌,其分布十分广泛,包括植物、人的胃肠道、动物、蔬菜或发酵乳制品[1]。 植物乳杆菌及其发酵产品具有重要的生理功能,包括调节肠道菌群、提高机体免疫力、降胆固醇等[2-3]。 传统的研究主要集中于采用生理生化实验方法来筛选具有特定功能的菌株,但这种方法筛选效率较低。 随着分子生物学技术和高通量测序技术的发展,采用基因组学和比较基因组学等手段将有助于系统了解植物乳杆菌的基因组结构,在分子水平上分析植物乳杆菌的功能,为高效筛选具有应用潜力的菌株提供新思路[4-5]。
近年来,研究人员采用 AFLP[6]、RAPD[7]、MLST[8]、CGH[9]等生物学手段,分析了植物乳杆菌基因组上的遗传多样性,同时,也进行了基因型和表型的比较研究。Molenaar 等人[10]研究了20 株植物乳杆菌的基因组,发现参与蛋白质和脂质合成或降解的基因在很大程度上是保守的,然而参与糖转运和分解代谢的基因在菌株中差异很大。 在另一项研究中,Siezen 等人[11]评估了185 株植物乳杆菌中 24 种糖的代谢规律和生长特征。 然而,这些研究并未将基因型和表型相关联。
大量使用抗生素促进了细菌不断进化,导致耐药细菌的出现[12]。因此,食品中所用菌株的安全性越来越得到重视。 食物链是人类和动物之间传播耐药菌的主要途径,一些发酵食品会为耐药细菌提供载体[13]。 发酵食品中常用的植物乳杆菌被认为是普遍安全的,但也存在将抗生素的耐药基因转移到病原菌的风险[14]。 因此,在将菌种应用到食品生产前,评估其抗生素抗性是十分必要的。 近段时间,一些学者对植物乳杆菌等益生菌的抗性基因和抗性之间的关系进行了探索, 但其研究多集中于单个菌株,忽略了菌株耐药性与其分离源的密切关系。
作者研究了从不同环境中分离的33 株植物乳杆菌的基因组特征,包括同源基因和基于同源基因的系统发育树, 并选取抗生素耐药性这一特性,首次对不同分离源的33 株植物乳杆菌的耐药性方面进行了全面研究,进一步探讨了比较基因组揭示的进化关系、抗性基因、抗生素抗性三者之间的关系,探究分离源、 菌株进化对菌株生物学特性的影响,为今后植物乳杆菌的基因组学及生物学特性等方面的研究提供数据参考和理论基础。
1 材料与方法
1.1 材料与试剂
1.1.1 菌株 33 株植物乳杆菌, 分离自发酵酱、发酵泡菜和人的粪便,保藏于江南大学食品生物技术研究中心的菌种保藏中心。
1.1.2 试剂 胰蛋白胨、无水葡萄糖、一水合硫酸锰、牛肉浸膏、无水乙酸钠、酵母粉、磷酸氢二钾、柠檬酸氢二铵、七水合硫酸镁:均购自国药集团化学试剂有限公司;链霉素、氯霉素、四环素、氨苄西林、红霉素、克林霉素、庆大霉素、新霉素、卡那霉素、环丙沙星、甲氧苄氨嘧啶:购自生工生物工程(上海)股份有限公司;IST 标准药敏检测肉汤培养基:购自山东拓普生物工程有限公司。
1.2 仪器与设备 超净工作台:江苏苏净集团有限公司;Whitley DG250 厌氧工作站:英国Don Whitley Scientific 公司;PCR 热循环仪:美国 Bio-Rad 公司;凝胶成像系统:美国Bio-Rad 公司;酶标仪:美国Thermo Fisher Scientific 公司;UV1800 紫外可见分光光度计:日本岛津公司。
1.3 实验方法
1.3.1 基因组测序 将植物乳杆菌接种到5 mL MRS 液体培养基中,37 ℃培养 12 h。 传代培养两次后, 接种到80 mL MRS 液体培养基中,37 ℃培养16~24 h。 培养结束,8 000 r/min 离心 5 min 收集菌泥, 送至上海美吉生物医药科技有限公司, 采用Illumina HiSeq 2000 进行基因组测序。
1.3.2 基因组组装、 预测及功能注释 使用SOAP denovo 软件对基因组进行组装, 验证单个碱基,使用 GapCloser 软件填充基因组内部缺口[15];使用Glimmer 和 Genmark 软件预测ORF (开放阅读框);在NCBI NR(非冗余蛋白质文库)和COG(直系同源簇) 蛋白质组数据库中通过BLASTp (蛋白质BLAST)对ORF 进行同源性比对和功能注释。
1.3.3 同源基因韦恩图绘制及系统发育树构建使用OrthoMCL 软件对每个菌株基因组进行直系同源基因预测[17],然后使用MAFFT v7.3 软件[18]比对直系同源基因, 通过 Perl 脚本绘制韦恩图。 使用PHYLIP v3.6[19](http://evolution.genetics. washington.edu/phtlip.html)构建系统发育进化树。
1.3.4 抗生素抗性基因分析 综合抗生素抗性数据库是CARD,使用BLAST 将植物乳杆菌的所有基因和CARD 数据库中的基因[20]对比,以预测基因组中存在的抗生素抗性基因。 若基因在CARD 中的序列匹配度达到阈值20%,则认为该基因组中含有对应的抗生素抗性基因。
1.3.5 抗生素耐受性实验 根据ISO 标准(ISO10932/ IDF223), 使用肉汤微量稀释法测定11种抗生素的最低抑菌浓度(MIC),抗生素包括环丙沙星、庆大霉素、新霉素、卡那霉素、氨苄西林、四环素、甲氧苄氨嘧啶、链霉素、氯霉素、红霉素和克林霉素。
参照ISO10932 / IDF223 文件, 采用二倍数稀释法,对 LSM(IST∶MRS=9∶1)培养基配置不同质量浓度抗生素稀释液;在无菌的96 孔板中,每行第1孔添加200 μL 空白培养基做对照; 第2 孔至第11孔加100 μL 抗生素稀释液(浓度由低到高),第12孔不加抗生素。 将菌株培养至稳定期,测菌液吸光度OD625;根据OD625值预估菌液的稀释比例,使得稀释后的菌液浓度在105~106CFU/mL。 将稀释的菌悬液在第2~12 孔中加100 μL,置于厌氧工作站中37℃培养,48 h 后取出。 结合OD625吸光度值及肉眼观察结果,无可见菌株生长的抗生素质量浓度即为该抗生素对菌株的MIC 值。 同一抗生素质量浓度,做平行3 个孔;做两次独立重复试验,最终确定抗生素对菌株的MIC 值。植物乳杆菌对不同抗生素的临界值判定依据EFSA 指南和EUC 标准,在EFSA 标准中,当测定的MIC 小于或等于临界值时,判定为敏感。 EUC 标准中,当测量的MIC 小于临界值时判定为敏感。 抗生素抗性实验质量浓度范围及临界值见表1。
2 结果与分析
2.1 菌株基本信息和基因组概况
本研究共选择了33 株不同来源的植物乳杆菌,其中12 株来自于发酵大酱,10 株来自于发酵泡菜,11 株来自于人体粪便,具体信息见表2。33 株植物乳杆菌基因组的大小范围为3.00~3.31 Mb, 粪便源的植物乳杆菌平均为3.16 Mb, 要高于泡菜源植物乳杆菌的3.12 Mb 及发酵酱源的3.11 Mb。 这可能是来自粪便的植物乳杆菌其生存环境更为复杂,需要进化出更多的基因和功能来适应环境。33 株植物乳杆菌的基因数量为3 049~3 416, 植物乳杆菌GC 碱基数量占总碱基数量的45.32%~46.33%。 这与Maria 等人[24]的研究结果是一致的。

表1 抗生素实验质量浓度范围及临界值Table 1 Antibiotic experimental concentration range and critical value
2.2 植物乳杆菌的同源基因及系统发育树
为了进一步明确植物乳杆菌在基因组层面是否存在差异, 我们分析了33 株不同来源的植物乳杆菌的直系同源基因,并基于同源基因构建了系统发育树。 在基因组进化过程中,同源基因是指具有相似生化功能的基因[25],同源蛋白质的基因家族也称为“核心基因家族”,菌株特有的基因称为特定基因家族[25-26]。
基于基因组聚类结果和核心基因组群绘制韦恩图,见图1。 直系同源基因的个数显示在图中心,特异性基因数量呈现在韦恩图的花瓣外侧,总基因数标注在菌株名称下方。 33 株植物乳杆中,共有2 245 个同源基因, 特殊基因的数量因来源不同存在明显区别。来自泡菜的菌株的特殊基因的数量在7~20 个,平均12 个,来自粪便和发酵酱的菌株的特殊基因的数量明显高于泡菜来源, 从7~147 个不等,发酵酱源为平均70 个,粪便源为平均71 个,这可能与样品所处环境受到的胁迫有关。
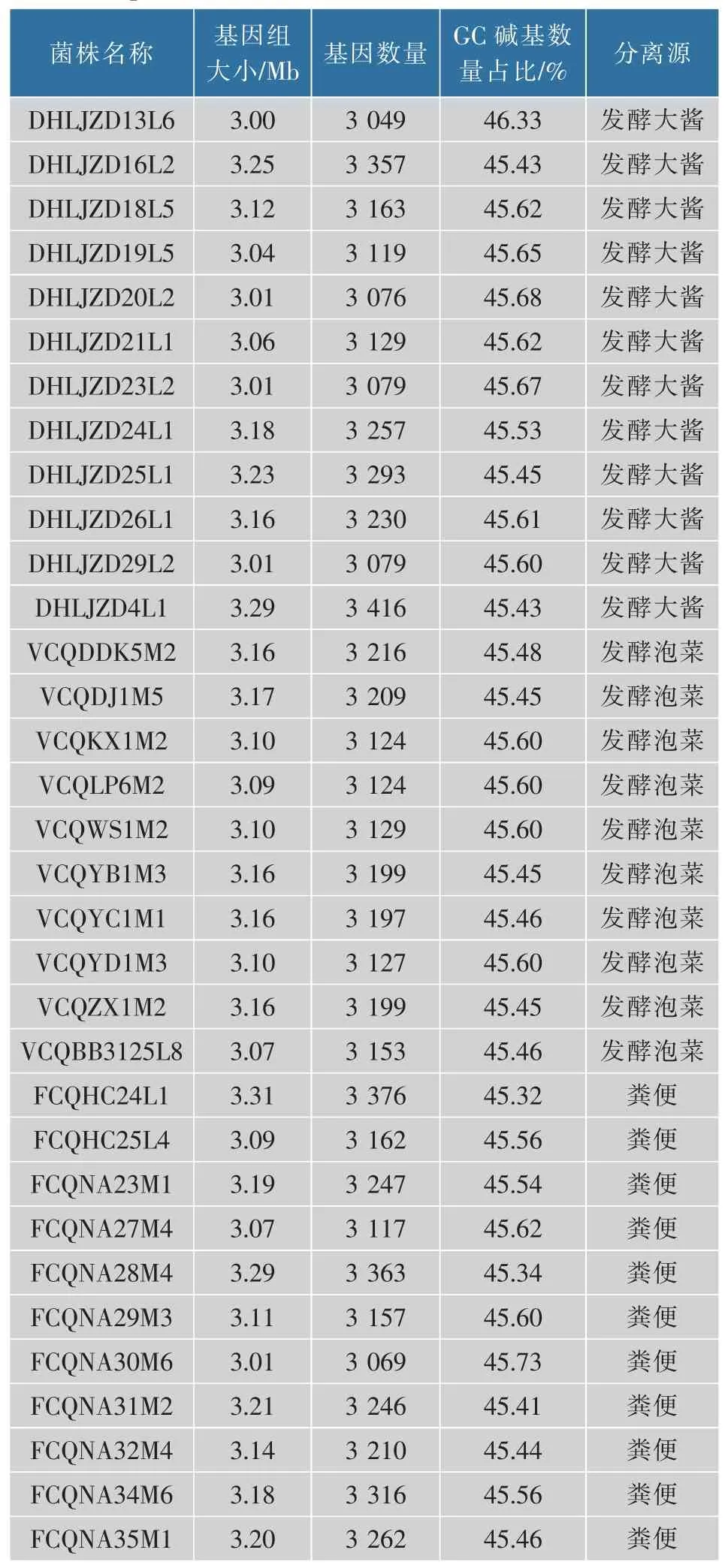
表2 33 株植物乳杆菌基本信息Table 2 Basic information of 33 strains of Lactobacillus plantarum
基于同源基因构建的系统发育树见图2。 进化树被分为3 大支, 其中来自发酵酱的DHLJZD13L6单独分为一大支,表明在基因层面上这株菌与其他植物乳杆菌有较大区别。 剩下两大分支表明来自泡菜、发酵大酱和粪便的植物乳杆菌能较好地根据来源聚成一簇,尤其是来自泡菜的植物乳杆菌,说明来源不同使得植物乳杆菌在基因层面上存在差异,生存环境赋予了菌株不同的基因型,从而使得其可能具有不同的功能。 Maria[24]等人研究了54 株来源于7 种分离源的植物乳杆菌的系统发育树,他发现分离源仅使得部分蔬菜源菌株呈现聚类趋势,整体上并没有对植物乳杆菌的进化造成显著影响,但这可能与蔬菜源菌株数多, 而其他分离源菌株数量少,导致规律不明显有关。

图1 33 株植物乳杆菌的直系同源基因Fig. 1 The orthologous gene of 33 strains of Lactobacillus plantarum
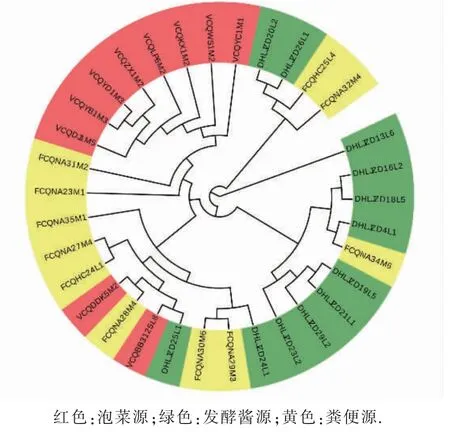
图2 基于同源基因的系统发育树Fig. 2 Phylogenetic tree based on homologous genes
2.3 抗生素抗性基因分析
依据抗生素抑菌机理的差异,可将抗生素大致分为以下5 种:抑制合成细胞壁、抑制合成核酸、抑制合成蛋白质、 改变细胞功能和抑制细胞代谢,本实验中使用的11 种抗生素类型及其常见对应基因[27]见表 3。

表3 实验中所用抗生素类型及基因Table 3 Types and genes of antibiotics used in the experiment
细菌耐受抗生素的分子和生化机制包括抗生素结合靶标的改变、耐药酶催化抗生素失活等[28]。根据基因组测序结果, 在33 株植物乳杆菌中共检测出74 种抗性基因, 其中有38 种基因属于外排泵,见图3。 36 种基因属于使抗生素失活型基因或者结合靶标改变基因,见图4。通过细菌细胞膜上的外排泵将抗生素排出胞外,是固有耐药和获得性耐药性的主要机制[29]。本实验中的33 株植物乳杆菌的外排泵机制主要属于MFS 和ATP 结合盒, 能够作用于多种类型的抗生素,例如氨基糖苷类、氟喹诺酮类等。 此外,发现红霉素外排泵基因macB 丰度最高;tetT 等四环素外排泵在33 株植物乳杆菌中均有发现;苯酚类抗生素(氯霉素)外排泵基因cmlv 仅存在于粪便源植物乳杆菌中。
其他36 抗性基因涵盖了氯霉素、环丙沙星、甲氧苄氨嘧啶、四环素、链霉素、克林霉素等,见图4。在抗生素抗性基因层面,不同来源的植物乳杆菌的抗性基因差异不大,只在个别基因上有区别,比如糖肽类抗性基因vanRF 只存在于粪便源和发酵酱源的植物乳杆菌中,而在所有泡菜源植物乳杆菌的基因组中发现了另一种糖肽类抗性基因vanRI。 在本研究中还发现,有些抗性基因是无法共存,例如莫匹罗星抗性基因mupA 和mupB。
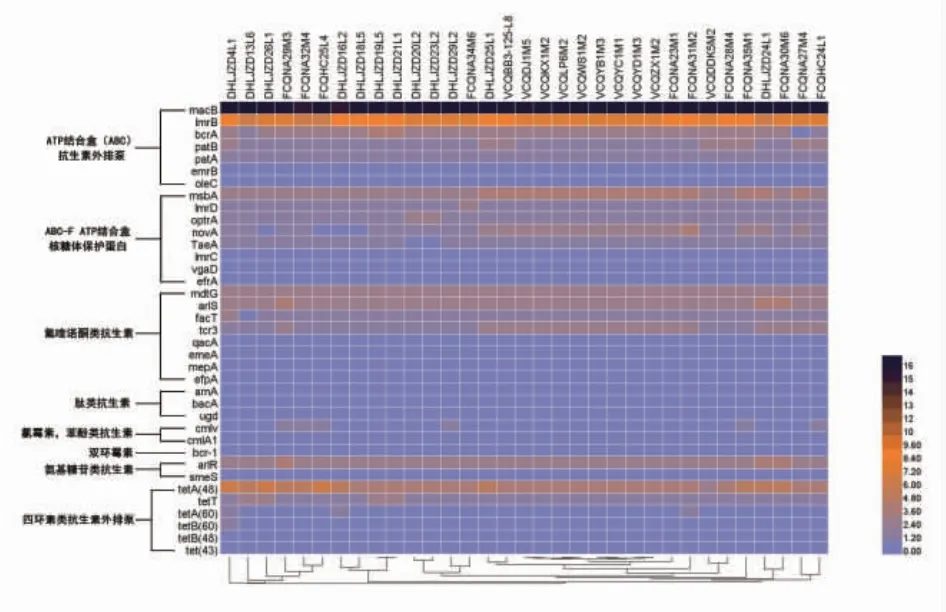
图3 33 株植物乳杆菌抗性基因中的外排泵基因Fig. 3 Efflux pump gene in 33 strains of Lactobacillus plantarum resistance gene

图4 33 株植物乳杆菌抗性基因中的其他类型基因Fig. 4 Other types of genes in the 33 strains of Lactobacillus plantarum resistance genes
2.4 植物乳杆菌抗生素抗性分析
为了明确基因型上的差异能否体现在表型上,测定了33 株植物乳杆菌对11 种抗生素的抗性,依据EFSA 和EUC 标准确定每种抗生素的临界值,根据 MIC 值将菌株划分为敏感(S)或抗性(R),并计算了不同来源的植物乳杆菌对每种抗生素的抗性菌株比例。
由表4 可知, 只有少数几株菌对庆大霉素、卡那霉素、链霉素、红霉素、新霉素、氨苄西林具有抗性,整体上表现出敏感;所有来源的菌株对四环素、环丙沙星具有抗性;对甲氧苄氨嘧啶、氯霉素,3 种来源的抗性菌株比例均达到70%以上。 不同来源的植物乳杆菌仅在克林霉素的耐受性上存在显著差异,其中泡菜源抗性最高,有80%的菌株表现为耐受,另外两种分离源的抗性菌株不足50%。

表4 3 种来源的植物乳杆菌对不同抗生素的抗性菌株比例Table 4 Proportion of resistant strains to different antibiotics from three sources
但进一步分析发现,虽然不同来源菌株对每种抗生素的MIC 范围不同,但出现的频率最高的MIC值基本是一致的,见图5。例如卡那霉素,发酵酱源菌株的 MIC 为 16~64 μg/mL, 泡菜来源菌株的 MIC 为16~128 μg/mL,粪便来源菌株的 MIC 为 8~64 μg/mL,但最高频率的MIC 值均为32 μg/mL。 其他抗生素的抗性结果也表现出了相似的规律,存在差异但不大。 这表明,分离源对植物乳杆菌的抗生素抗性影响并不显著。
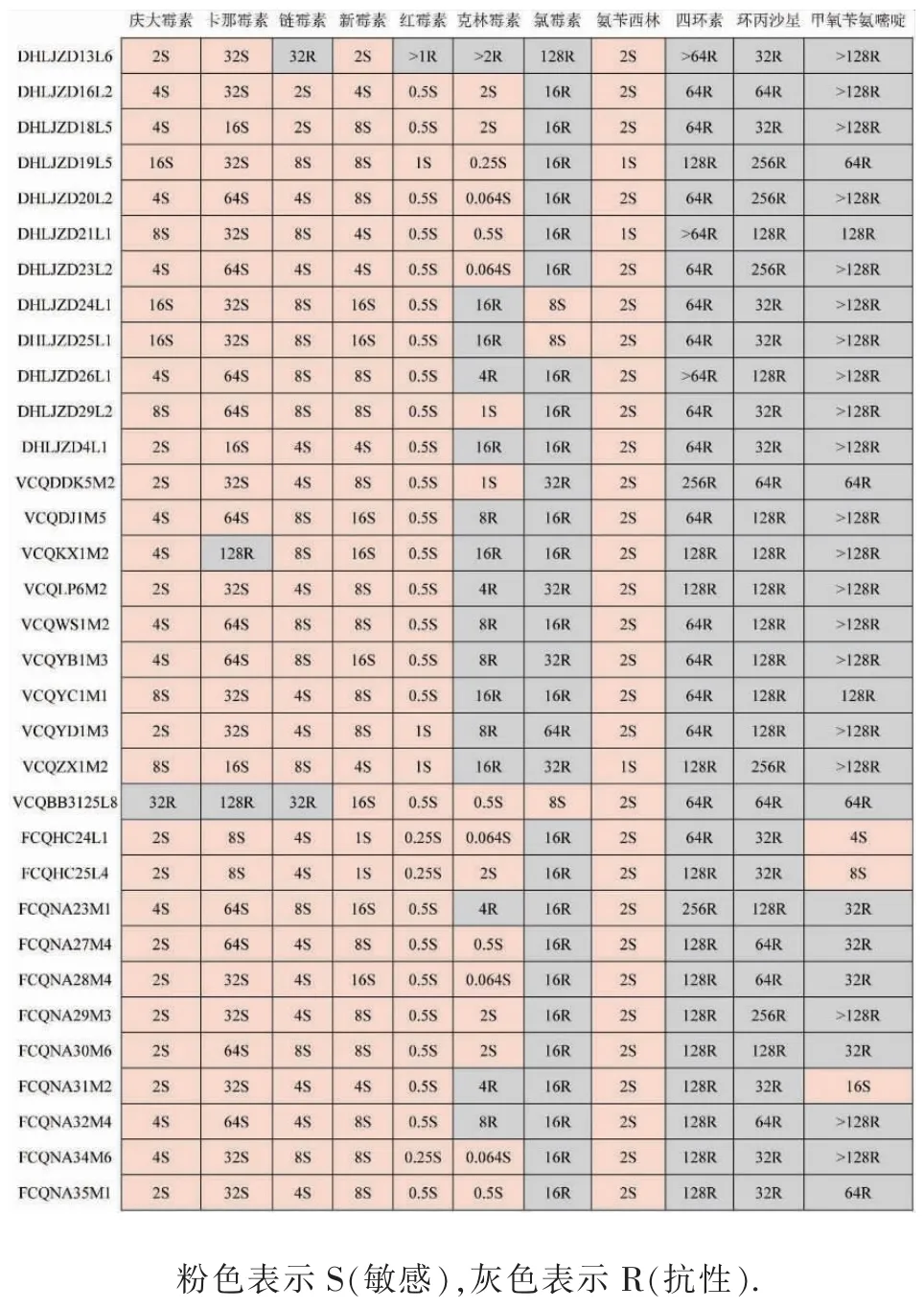
图5 33 株植物乳杆菌对不同抗生素的MIC 值Fig. 5 MIC values of 33 Lactobacillus plantarumstrains for different antibiotics
杨梅[14]研究了93 株、9 个种属的乳酸菌的耐药性, 结果表明同种乳酸菌的抗生素抗性差异不大,不同种之间差异明显,实验中所有乳杆菌属对青霉素和红霉素耐药性差,这与本研究结果一致。 周宁等[30]对分离自市售酸奶的18 株保加利亚乳杆菌的抗生素耐药性进行研究, 结果表明18 株保加利亚乳杆菌均对罗红霉素敏感, 而对卡那霉素均耐受。大量的研究表明, 乳酸菌中的乳杆菌属对万古霉素、卡那霉素、链霉素、甲硝哩、氟呱酸、环丙沙星等具有固有耐受性。
作者发现有两株菌比较特别, 泡菜源的VCQBB3125L8 和发酵酱DHLJZD13L6,其耐受抗生素的种类要略广于其他菌株。 出现这种情况的原因可能有两种,一是这两株都来自于发酵食品,制作过程中没有经过高温杀菌,导致菌株与环境中的其他微生物发生了耐药基因的转移;二是用于制作这两种发酵食品的原料在生长过程中可能接触到含有多种抗生素的农药或肥料,导致分离出的这两株菌的抗性增强。
2.5 抗生素抗性与抗性基因的关系
理论上,菌株的获得耐药性会增加耐药基因转移的风险,固有耐药性的菌株因为缺乏相关抗性基因较为安全。 因此,研究菌株的抗性基因和抗性表型的关系,对于判断菌株为获得耐药性还是固有耐药性,改造菌株应用于实际生产具有重要意义。
分析抗生素的MIC 值和抗性基因检测结果,我们发现大部分抗生素抗性可以与其抗性基因相关联。 环丙沙星是典型的氟喹诺酮类抗生素,通过抑制核酸合成达到抑菌的效果[31],所有菌株均含有氟喹诺酮类抗性基因gyrA、gyrB,MIC 结果表明菌株对环丙沙星具有抗性; 四环素抗性基因主要为tetT等,所有菌株均含有四环素类抗性基因和外排泵基因,实验也发现所有菌株对四环素具有抗性;分析氨苄西林、氯霉素和甲氧苄氨嘧啶,也得到同环丙沙星和四环素类似的结果。
万古霉素是糖肽类抗生素,主要通过抑制细胞壁合成达到抑菌效果[32],以往的研究称植物乳杆菌对万古霉素有天然抗性[33]。本研究中33 株植物乳杆菌均含有大量的万古霉素抗性基因 (van 系列),且33 株菌均筛选自含万古霉素的培养基,因此可以判定33 株菌对万古霉素均有抗性。
发酵酱源的DHLJZD13L6 对链霉素表现出抗性,在其基因组中发现了链霉素抗性基因ANT(9)-Ia;绝大部分菌株对庆大霉素、链霉素、卡那霉素、新霉素敏感,在其基因组中没有发现与这些抗生素密切相关的抗性基因。 研究发现泡菜源的VCQBB3125L8 也没有相关基因,但其对庆大霉素、卡那霉素、链霉素均表现出抗性,这可能与植物乳杆菌能对庆大霉素、链霉素、卡那霉素和新霉素等低水平氨基糖苷类抗生素表现出固有抗性[22]有关。
但红霉素和克林霉素基因和表型并不对应,可能是基因、移动元件及其细菌宿主之间发生了复杂相互作用[23]。 研究表明菌株对红霉素产生抗性主要由 mefA 和存在于质粒上的 ermB 基因介导[16,21]。 虽然所有菌株都含有红霉素外排泵基因macB,数量达到17 个,但只有DHLJZD13L6 表现出了抗性,可能是因为其质粒上含有ermB 基因。 所有菌株的基因中均检测出克林霉素外排泵基因lmrC、vgaD, 但是3 种来源的菌株其抗性并不相同,粪便源最敏感,抗性菌株只占27.27%,泡菜源抗性达到80%。 这可能是因为泡菜是发酵制品,在制作过程中没有高温杀菌,暴露的环境、多样的微生物对其产生了选择作用。
值得一提的是, 来自发酵酱的植物乳杆菌DHLJZD13L6 对红霉素、链霉素、克林霉素也都表现出了抗性,一是因为其含有相关抗性基因,二是以往的研究表明这3 种抗生素有交叉耐药性。 同时,我们发现DHLJZD13L6 含有多个独特的抗性基因,如 ANT(9)-Ia、oleC、murA,这与系统发育树表现的结果相同。 这表明,比较基因组学对于挖掘可能具有同种生理特性和功能的菌株具有指导意义。
3 结 语
对33 株不同来源的植物乳杆菌的多样性进行了初步探究, 包括基因组层面以及抗生素抗性层面。 比较基因组学结果表明,分离源对植物乳杆菌的遗传进化具有较为显著的影响。 粪便源的菌株在基因组大小和特殊基因数量上均多于泡菜源和发酵酱源的菌株,这与其生存环境复杂有关。 33 株植物乳杆菌中共检测出74 种抗性基因, 不同来源的菌株仅在vanRI 等个别抗生素基因上呈现聚类的趋势,整体上分离源对菌株的抗生素抗性基因和抗性上并没有造成明显的差异。33 株植物乳杆菌均含有环丙沙星、四环素、氯霉素、甲氧苄氨嘧啶和万古霉素的抗性基因, 其抗性菌株比例也达到90%以上。绝大多数菌株对庆大霉素、链霉素、卡那霉素、新霉素、氨苄西林敏感,基因组中也没有发现与其对应的抗性基因。 但对于红霉素和克林霉素,其基因和表型并不对应。
抗生素抗性实验结果表明,基因型和表型有很强的联系,证明了基因组学对于表型分析有指导意义。 同时,菌株的抗性结果遵循比较基因组学得出的规律,表明比较基因组学对于挖掘可能具有同种生理特性和功能的菌株具有重要意义。
