内含子编码蛋白Mg2+结合位点功能分析及验证
2020-11-02崔古贞陈相好洪伟张峥嵘綦廷娜陈峥宏
崔古贞 陈相好 洪伟 张峥嵘 綦廷娜 陈峥宏
(1. 贵州医科大学基础医学院,贵阳 550025;2. 遵义医科大学第三附属医院(遵义市第一人民医院),遵义 563002;3. 贵州省普通高等学
校病原生物学特色重点实验室,贵阳 550025;4. 贵州省分子生物学重点实验室,贵阳 550004)
II型内含子(Group II intron)是一类反转录转座子(Retrotransposon),在细菌、古细菌以及真核生物叶绿体和线粒体基因组中广泛存在,尤其以细菌中最为普遍[1-4]。目前,通过现代基因组测序已鉴定了数千种细菌II型内含子,在鉴定的细菌II型内含子中,研究最为详细的主要包括两类,第一类是来源于乳酸乳球菌(Lactococcus lactis,Ll.LtrB)的嗜中温II型内含子[5-9],另一类是来源于Thermosynechococcus elongatus及Geobacillus stearothermophilus的嗜高温II型内含子[2,10-12]。以此两类II型内含子为基础,利用其“归巢”机制,已开发出了高效基因打靶技术——Targetron技术(中温型)和Thermotargetron技术(高温型),并在多种微生物基因工程中获得广泛应用[13-19]。
II型内含子由内含子RNA和内含子编码蛋白(Intron encoded protein,IEP)两部分组成,其中内含子编码蛋白IEP具有DNA聚合酶、反转录酶、核酸内切酶等多种酶活性,在II型内含子“归巢”过程中发挥重要功能[20-21]。其“归巢”过程主要包括:首先,内含子RNA与内含子编码蛋白相互识别,组装为有活性的核糖核蛋白复合体(Ribonucleoprotein)。然后,内含子RNA通过碱基互补配对原则识别双链DNA靶位点。IEP蛋白在内含子RNA的辅助下,利用其自身的核酸内切酶活性反向剪切双链DNA,形成双链DNA缺口。然后,IEP蛋白的反转录酶活性,以切口处的3'-OH断端为引物,以内含子RNA为模板,反转录合成互补cDNA,再以新合成的cDNA为模板,通过DNA聚合酶活性合成双链DNA,最后通过DNA修复机制,实现内含子在靶位点的“归巢”[9,19,21](图1)。
在整个内含子“归巢”过程中,高浓度Mg2+是保证其功能的重要条件,这也是II型内含子在真核生物细胞内效率极低的主要原因(真核生物细胞核内Mg2+浓度较低)[7-8,22-23]。IEP蛋白反转录酶活性在II型内含子的“归巢”过程中发挥重要功能,其Mg2+结合位点是影响其反转录功能的关键催化位点,如果能克服其依赖高浓度Mg2+的缺点,便有可能将II型内含子广泛应用于真核细胞的遗传改造。因此,为了研究Mg2+结合位点对II型内含子“归巢”效率的影响,本研究利用生物信息学手段筛选影响IEP与Mg2+结合的关键氨基酸催化位点,并利用基因工程手段对其进行定点突变,最后,构建突变型Targetron载体,在大肠杆菌中验证其功能,旨为深入研究II型内含子“归巢”机理奠定基础。
1 材料与方法
1.1 材料
1.1.1 试剂 高保真DNA聚合酶、T4 DNA连接酶、

图1 II型内含子“归巢”示意图
DNA Marker、DNA loading buffer购自北京全式金公司,DNA纯化及回收试剂盒、质粒提取试剂盒、快速定点突变试剂盒购自天根生化科技有限公司,蛋白胨、酵母膏、氯化钠,琼脂糖、异丙基硫代半乳糖苷(IPTG)、5-溴-4-氯-3-吲哚-β-D-半乳糖苷(X-gal)、氨苄青霉素购自北京索莱宝科技有限公司,限制性内切酶购自赛默飞公司,其他常用生化试剂均购自国药集团化学试剂公司。
1.1.2 仪器 WD-9413B成像仪、DYY-7C型电泳仪、Legend Micro 17R冷冻高速离心机、K960 PCR仪。
1.1.3 引物 本研究所用引物均由上海生物工程有限公司合成,引物序列见表1。
1.1.4 菌株及质粒 本研究所用菌株及质粒见表2。
1.2 方法
1.2.1 菌株的培养 本研究所用的大肠杆菌DH5α和BL21(DE3)均在37℃、LB液体或固体培养基中培养,氨苄青霉素使用浓度为100 μg/mL,蓝白斑筛选时加入X-gal(40 μg/mL)和IPTG(0.1 mmol/L)进行筛选。

表1 本研究用到的引物

表2 本研究用到的菌株及质粒
1.2.2 生物信息学分析 利用NCBI中的保守结构域数据库(Conserved Domain Databse,CDD),以来源于乳酸乳球菌Ll.LtrB的内含子编码蛋白序列为基础进行序列比对,分析保守的关键氨基酸位点,筛选与Mg2+结合的可能氨基酸残基。
1.2.3 定点突变 以IEP-F/IEP-R为引物扩增野生型IEP获得其DNA产物,并与pMD19T连接得到载体pMD19T-IEP,以载体pMD19T-IEP为模板,利用定点突变引物进行定点突变PCR,扩增体系为:模板pMD19T-IEP 1 μL,正向及反向突变引物均为2 μL,5×buffer 10 μL,DNA Polymerase 1.5 μL,加ddH2O至总体积为50 μL,PCR扩增条件为:95℃ 30 s,(95℃ 30 s,55℃ 20 s,72℃ 4 min)15个循环,72℃5 min。反应结束后加入1 μL的DpnI限制性内切酶,37℃条件下反应1 h去除模板质粒,随后转化大肠杆菌DH5α,菌落PCR结合测序进行验证,最终获得含有IEP突变位点的载体pMD19T-M-IEP(图2)。
1.2.4 Targetron载体构建 利用NheI和BglII双酶切质粒pMD19T-M-IEP,与经同样酶切的载体pSY7-lacZ-635s、pSY7-lacZ-1063分别连接,连接体系为:载体1 μL,目的片段4.5 μL,T4 DNA ligase buffer 2 μL,T4 DNA ligase 0.5 μL,加ddH2O至总体积10 μL,25℃反应1 h后转化感受态细胞E. coliDH5α,菌落PCR结合DNA测序验证突变型Targetron载体pSY7-M-lacZ-635s/1063a(图2)。下文所用到的突变型Targetron载 体pSY7-M3-lacZ-635s/1063a、pSY7-M4-lacZ-635s/1063a、pSY7-M5-lacZ-635s/1063a,其中M3表示IEP D308A单点突变,M4表示IEP D309A单点突变,M5表示IEP D308A/D309A双点突变。
1.2.5 打靶效率检测 将突变型Targetron载体pSY7-M-lacZ-635s/1063a与及野生型Targetron载体pSY7 -lacZ-635s/1063a分别转化大肠杆菌BL21(DE3)感受态细胞,复苏后加入5 mL的LB液体培养基(含100 μg/mL 氨苄青霉素),37℃、180 r/min培养12 h。取50 μL过夜培养的菌液加入5 mL含100 μg/mL 氨苄青霉素的LB液体培养基中,37℃、200 r/min条件下培养1 h,随后加入IPTG使其终浓度为0.5 mmol/L,诱导45 min,以不同浓度的稀释度涂布LB固体培养基(含氨苄青霉素100 μg/mL、X-gal 40 μg/mL、IPTG 0.1 mmol/L),37℃过夜培养,菌落PCR结合蓝白斑筛选计数计算其“归巢”效率(归巢效率=白斑/白斑+蓝斑×100%)。
1.2.6 菌落PCR验证 以lacZ基因打靶位点外侧引物进行菌落PCR验证,分别检测平板上的蓝色菌落及白色菌落,进一步验证IEP蛋白反转录结构域突变后对打靶效率的影响。
2 结果
2.1 IEP蛋白生物信息学分析
在NCBI数据库中,以乳酸乳球菌Ll.ltrB的IEP蛋白序列(HQ263410.1)为模板进行序列比对及分析,发现细菌II型内含子反转录酶具有极其保守的YADD序列(图3),结构预测表明其两个毗邻的天冬氨酸残基(DD)与Mg2+结合,可能是负责反转录功能的关键催化位点。因此,本研究选择这两个天冬氨酸残基为靶点,即D308和D309两个位点,构建突变型IEP蛋白突变体。
2.2 Mg2+结合位点突变对Ⅱ型内含子“归巢”效率的影响
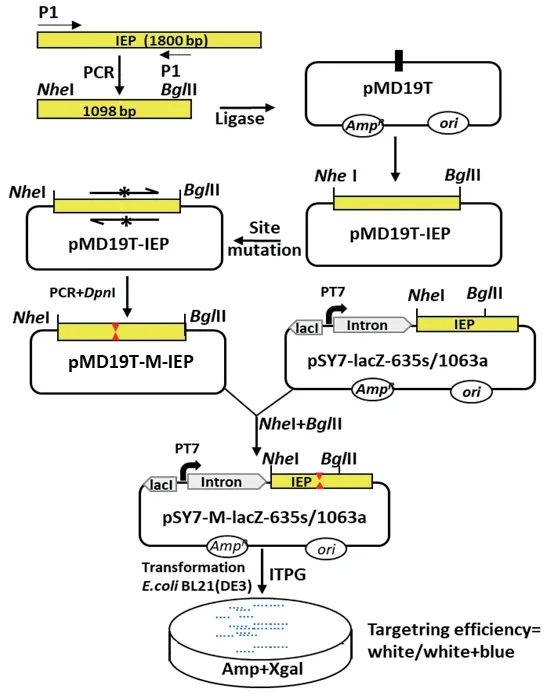
图2 突变型IEP蛋白构建及打靶效率检测
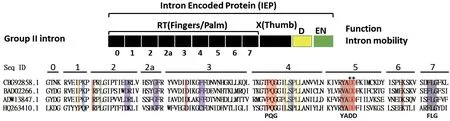
图3 IEP蛋白生物信息学分析
为了验证D308A、D309A、D308A/D309A突变对II型内含子“归巢”效率的影响,将突变型IEP蛋白取代Targetron载体上的野生型IEP蛋白,构建Mg2+结合位点突变的Targetron载体,同时以野生型Targetron载体作为对照,分别转化大肠杆菌BL21(DE3),IPTG诱导后涂布含有Xgal和IPTG的平板,过夜培养后通过菌落PCR(图4)及蓝白斑筛选(图5-A),分析IEP蛋白Mg2+结合位点突变对Ⅱ型内含子“归巢”效率的影响。研究结果表明,野生型Ⅱ型内含子在大肠杆菌中针对lacZ-635s位点的“归巢”效率为90.845%±6.792%,针对lacZ-1063a位点的“归巢”效率为92.582%±2.898%[24];然而,将IEP蛋白反转录结构域Mg2+结合位点(D308A、D309A、D308A/D309A)突变后,平板均为蓝斑,菌落PCR验证其靶位点也没有任何内含子插入,其“归巢”效率均降低为0%(图4-5)。此结果表明,IEP蛋白反转录结构域Mg2+结合位点突变,失活了IEP蛋白的反转录功能,使Ⅱ型内含子不能以内含子RNA为模板合成cDNA而插入到DNA靶位点,即II型内含子完全失去其“归巢”功能。
3 讨论
细菌Ⅱ型内含子因其极高的“归巢”效率,目前已被设计为高效基因打靶工具,在革兰阴性细菌、阳性细菌中获得广泛应用。尽管在真核生物甚至人类细胞中也能够实现基因打靶功能[26-28],但由于真核生物细胞核中Mg2+浓度较低,极大影响了其打靶效率,限制了其广泛应用。
内含子编码蛋白IEP在内含子“归巢 ”过程中发挥着重要的功能。IEP蛋白是由4个相对独立的结构域组成,包括:反转录结构域(RT)、成熟酶结构域(X)、DNA结构域(D)、核酸酶结构域(EN),此4个结构在功能上和空间上相对独立[1,8,29]。因此,突变其中一个结构域对其他结构域的影响相对较小。本研究通过突变RT结构域的Mg2+结合位点,获得了失活反转录功能的IEP蛋白,理论上对其他结构域的功能影响可能不大,这为深入研究II型内含子的功能提供了前提。
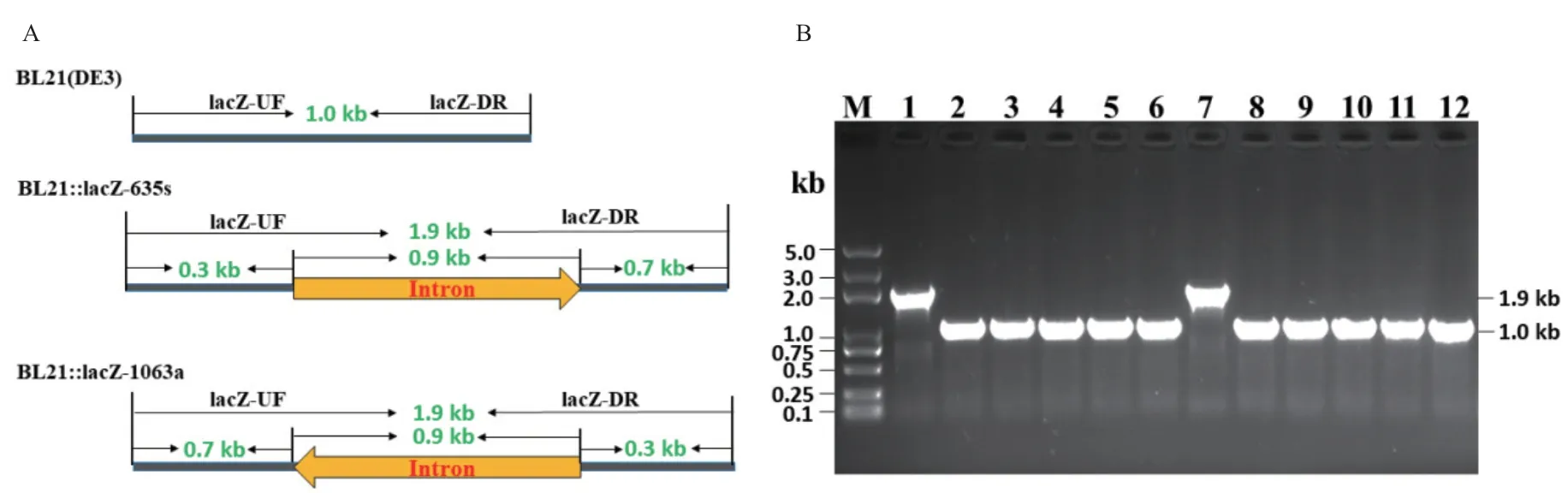
图4 突变型II型内含子功能验证
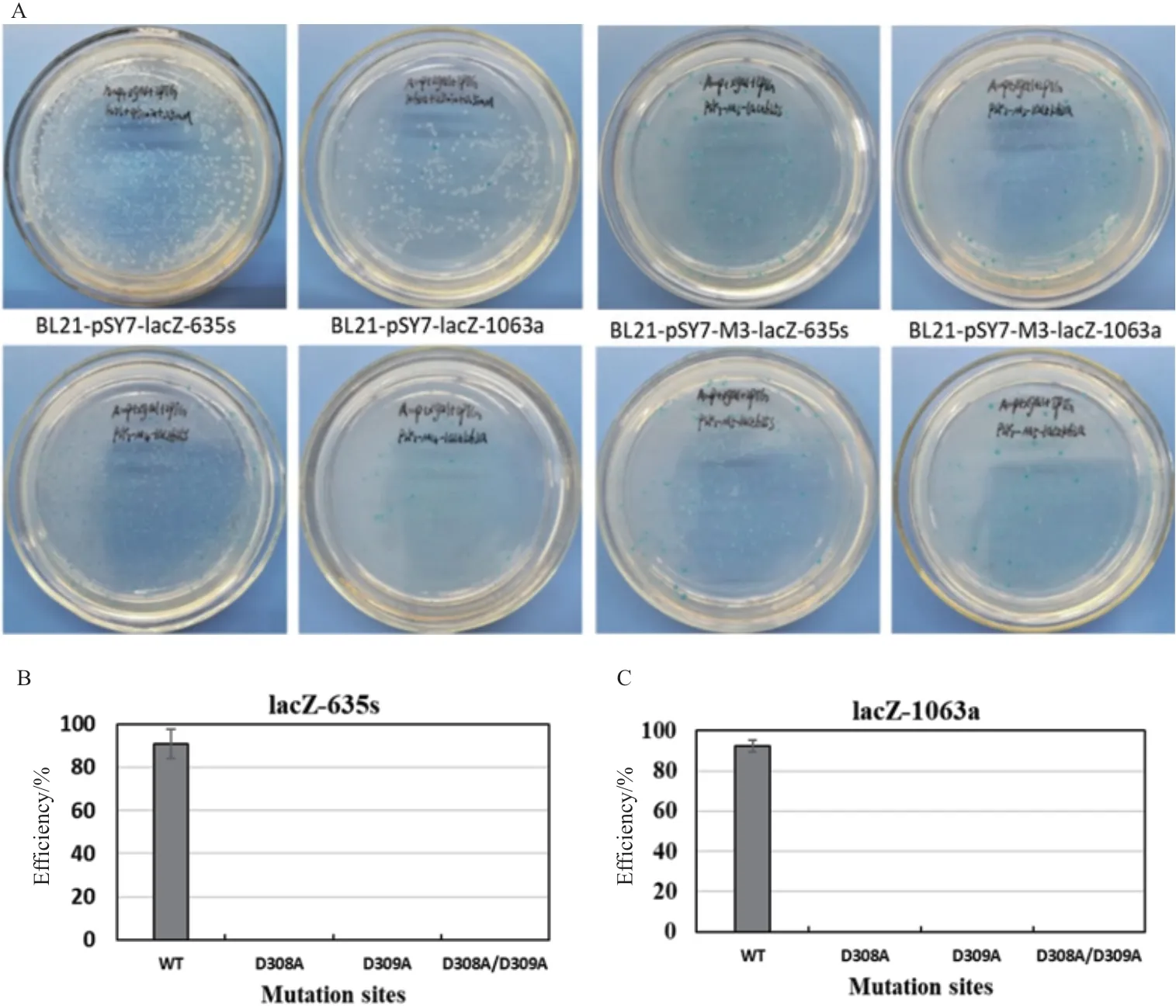
图5 蓝白斑筛选及“归巢”效率比较突变
此外,在以前工作中,我们已筛选到C164和Y208两个位点也是影响II型内含子“归巢”的核心催化位点,该位点突变后其“归巢”功能也完全丧失[24],与本研究筛选的D308和D309位点具有相似的功能。然而,在IEP反转录过程中,C164和Y208两个位点分别与底物NTP和模板DNA结合,而D308和D309位点是与Mg2+结合,因此,D308和D309位点突变对其他结构域功能的影响可能比C164和Y208位点小。在后续研究中,我们以构建的Mg2+结合位点突变体为基础,发现该位点不但能失活II型内含子的“归巢”功能,而且还能在靶位点引入DNA损伤(数据尚未发表),表明D308和D309位点突变对II型内含子识别、切割DNA靶位点的影响较小,与C164和Y208位点突变相比,可能更有利用于II型内含子结构的稳定[29]。众所周知,靶向DNA损伤恰是基因编辑的前提[30],预示本实验构建的突变型II型内含子可能具有基因编辑的潜能,因此,本实验构建的Mg2+结合位点突变体将为深入研究Ⅱ型内含子的功能及应用奠定良好的基础。
4 结论
本实验利用生物信息方法及定点突变技术,筛选并构建了IEP蛋白反转录结构域Mg2+结合位点突变体(D308A和D309A单点突变,D308A/D309A双点突变),并以大肠杆菌为基础,体内分析了IEP蛋白Mg2+结合位点突变对II型内含子“归巢”效率的影响,结果表明,Mg2+结合位点突变完全失活了II型内含子的“归巢”功能。
