二系杂交小麦亲本光周期、春化和矮秆 等位基因的组成和分布
2020-10-30陈现朝张立平秦志列廖祥政张风廷赵昌平
陈现朝,张立平,秦志列,廖祥政,孙 辉,张风廷,赵昌平
(北京市农林科学院 北京杂交小麦工程技术研究中心/杂交小麦分子遗传北京市重点实验室,北京 100097)
基于小麦光温敏不育系建立的二系杂交小麦技术,已成为小麦杂种优势利用的主要途径。由于二系杂交小麦亲本材料来源广泛,需要对材料的光温敏的特性进行田间鉴定和评价工作,以获得生产应用价值更高的骨干亲本。小麦光温敏特性主要由光周期、春化、开花期基因控制,此外光温特性和株高往往决定小麦品种适于种植的生态区。控制光周期特性的基因包括Ppd-D1 (Ppd1)、Ppd-B1 (Ppd2) 和Ppd-A1 (Ppd3),分别位于染色体2D、2B 和2A。Beals 等根据大麦Ppd-H1 基因,同源克隆到小麦Ppd 基因并开发了Ppd-D1 的功能标记[1],该标记已用于青海和西藏小麦品种等位变异检测[2]。春化基因决定生长习性,影响开花时间。依据通过春化阶段所需的温度和时间将小麦划分为冬性和春性。研究发现控制小麦春化特性的基因有Vrn-A1、Vrn-B1、Vrn-D1、Vrn-B3、Vrn-D3 和Vrn-D4,分别位于5A、5B、5D、6B、7D 和5D 上,其中Vrn-A1、Vrn-B1、Vrn-D1 和Vrn-B3 已被克隆[3-5],开发的功能标记已用于小麦品种检测[6-7]。世界上已发现的Rht (Reduced height)矮秆基因有24 个, 但在推广小麦品种中包含的Rht 基因主要是Rht-1B、Rht-1D、Rht8[8],其中Rht-1B、Rht-1D 开发了功能标记并用于小麦品种检测[9]。
光周期、春化和株高等性状的田间鉴定受环境因素影响较大。近年来,随着高通量测序技术的发展,由LGC 公司研发的竞争性等位基因特异性PCR 技术具有高度稳定性、准确性和低成本优势,逐渐被用于重要功能基因的SNP (Single nucleotide polymorphism, SNP) 和InDels (Insertion-deletion, InDels)检测[10]。本团队前期研究表明,双亲为地理远缘、生态远缘时,二系杂交小麦的增产优势最为显著[11]。但是目前主要通过多年多点的田间表型测定来鉴定二系杂交小麦育种亲本的光温生态及株高特性,对于其基因组成和分布等研究仍是空白,这已成为限制杂交小麦亲本选育、组合配置以及组合筛选效率提高的重要瓶颈。本研究利用光周期、春化和矮秆性状的6 对KASP 功能标记,对245 份二系杂交小麦亲本材料进行检测,分析上述基因的等位变异类型和分布频率,初步了解杂交小麦亲本材料中光温生态及株高性状的基因等位变异信息,为进一步开展二系杂交小麦分子育种奠定基础。
1 材料和方法
1.1 供试材料
以245 份二系杂交小麦亲本为供试材料,其中恢复系233 份,不育系12 份,于2017—2018 年度种植于邓州杂交小麦产业化基地(北纬N32°40′7.41″ 东经E112°08′54.46″)。
1.2 KASP 标记引物设计
参照Rasheed A 研究结果[12]设计本研究所需的6 对KASP 标记引物,根据KASP 技术的引物设计要求,FAM 引物和HEX 引物5′端分别含有特异性接头,GAAGGTGACCAAGTTCATGCT和GAAGGTCGGAGTCAACGGATT, 可 与 荧光标记结合用于荧光检测。具体引物序列信息如 表1 所示。
1.3 DNA 提取
小麦返青期,田间剪取供试恢复系和不育系叶片。利用植物基因组DNA 提取试剂盒(天根生化科技有限公司,北京)分别提取245 份供试小麦材料基因组DNA,供试基因组DNA 在10 ~30 ng/μL。紫外分光光度计Nanodrop2000(Thermo)检测A260/A280比值在1.8 左右,A260/A230比值大于1.8。

表1 试验所需引物Table 1 Primers in the test
1.4 KASP 标记检测及数据统计
供试小麦材料基因组DNA 为模板,分别采用6 对引物进行PCR 扩增,得到PCR 扩增产物。反应程序为:94 ℃预变性,15 min;94 ℃变性20 s,61 ~55 ℃(选用Touch down 程序,每循环降低0.6 ℃) 1 min,扩增10 个循环;94 ℃变性20 s,55 ℃延伸1 min,继续扩增26 个循环。待PCR 扩增产物温度降至40 ℃以下时通过酶标仪FAM、HEX 光束扫描读取荧光值,并做基因型分析。试验结果剔除基因型杂合和缺失样本,用Excel 统计恢复系和不育系材料中不同基因型的频率并计算基因型的百分比。
2 结果与分析
2.1 KASP 标记分型
选择光周期、春化和矮秆性状的6 对KASP 功能标记,对245 份二系杂交小麦亲本材料进行鉴定。KASP 标记对上述基因进行分型,其中光周期基因Ppd-B1、春化基因Vrn-A1 和矮秆基因Rht-D1 的KASP 分型效果如图1 所示。

图1 KASP 标记在供试材料光周期Ppd-B1(A)、春化Vrn-A1(B)和矮秆Rht-D1(C)基因分型效果Fig. 1 Result of the tested materials by selected KASP assays of photoperiod gene Ppd-B1 (A), vernalization gene Vrn-A1 (B) and dwarf gene Rht-D1 (C)
2.2 光周期基因、春化基因和矮秆基因组成
本研究检测的光周期基因有Ppd-A1、Ppd-B1和Ppd-D1(表2)。检测结果显示二系杂交小麦亲本主要以光周期敏感的类型为主。在245 份供试材料中均发现光周期敏感基因型Ppd-A1b、Ppd-D1b,216 份材料中检测到光周期敏感基因型Ppd-B1b;只有29 份材料含有光周期不敏感基因型Ppd-B1a,其中恢复系28 份,不育系1 份,其比率分别为12.02%和8.33%。
本研究检测的春化基因为Vrn-A1,检测结果显示二系杂交小麦亲本主要以冬性为主。242 份材料含有冬性基因型vrn-A1,占全部供试材料的98.78%,其中恢复系230 份,不育系12 份;其比率分别为98.71%和100%。只有3 份恢复系材料含有春性基因型Vrn-A1,其比率为1.29%(见表2)。
本研究检测小麦矮秆基因为Rht-B1 和Rht-D1。检测结果显示96.73%的材料含有Rht-B1a基因型,其中恢复系229 份,不育系8 份;在恢复系和不育系的比率分别98.28%和66.67%。89.8%的材料含有Rht-D1a 基因型,其中恢复系220 份,其比率为94.42%;10.2%的材料含有Rht-D1b 基因型,其中恢复系13 份,不育系12 份;在恢复系和不育系的比率分别5.58%和100%(见表2)。

表2 光周期、春化和矮秆等位基因类型及频率Table 2 Allelic gene type and frequency of photoperiod, vernalization and dwarf genes
2.3 恢复系和不育系基因型及频率
检测结果表明,供试材料中光周期和春化基因型Ppd-A1b+Ppd-B1b+Ppd-D1b 和vrnA1a 占 主 导地位,在恢复系的比率分别91.67%和98.71%,在不育系的比率分别87.98%和100%。这表明二系杂交小麦亲本主要为光周期敏感和冬性类型材料。
检测结果表明,不育系中矮秆基因型Rht-D1b+Rht-B1a-160 bp 和Rht-D1b+Rht-B1a 比 例 为1∶2。恢复系中矮秆基因型Rht-D1a+Rht-B1a 高达92.70%。这表明供试恢复系和不育系材料中矮秆基因型存在明显的差异。
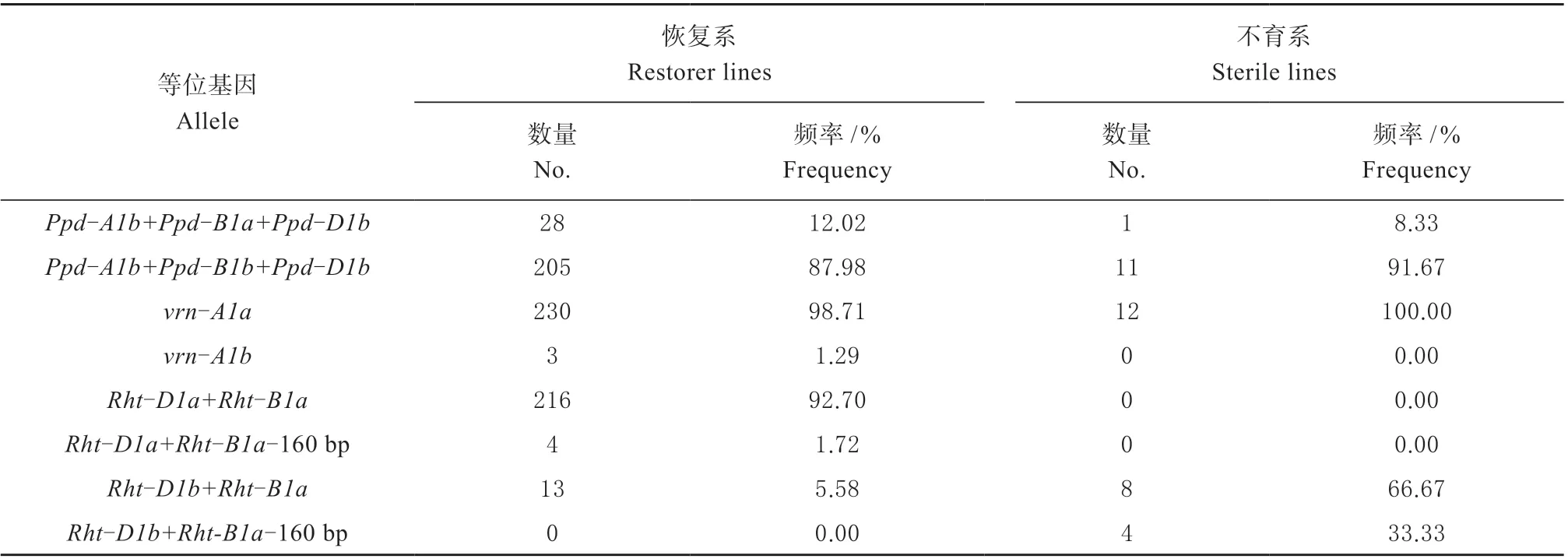
表3 恢复系和不育系基因型数量和频率Table 3 Number and frequency of gene type in restorer lines and sterile lines
3 讨论
3.1 光周期和春化基因型、光温特性和地域分布的关系
光周期基因型可以反映品种的光周期特性。研究表明光周期基因Ppd-A1、Ppd-B1 和Ppd-D1 是调控小麦光周期敏感性的重要遗传因素之一,对光周期不敏感作用强弱顺序分别为Ppd-D1、Ppd-B1和Ppd-A1[1,13]。此外,Ppd-D1a 等位基因检测可以作为小麦品种光周期的鉴定方法[14]。在不同的种植区选择适宜的小麦品种,可以充分利用光照资源,能够最大限度发挥品种的潜力,从而提高产量。本研究发现在二系杂交小麦亲本中,88%的不育系和91.7%的恢复系是光周期敏感型Ppd-A1b+Ppd-B1b+Ppd-D1b。这可能与选育或筛选出的亲本材料地处北部冬麦区有关,光周期敏感型基因有利于推迟小麦拔节,防止小麦过早形成花器官,避免遭受早春霜冻的伤害。
控制小麦春化特性的基因有Vrn-A1、Vrn-B1、Vrn-D1、Vrn-B3、Vrn-D3 和Vrn-D。Vrn-A1 的春化作用最强,且对Vrn-B1 和Vrn-D1 有上位性效应[15]。由于Vrn-1 基因分为显性和隐性两种类型,显性基因对应春性,隐性基因对应冬性[16]。本研究发现供试材料中98.7%的恢复系和全部不育系含有vrn-A1,1.3%的恢复系含有Vrn-A1,说明二系杂交小麦大部分恢复系和不育系是含vrn-A1 基因型为主的材料。由于本研究尚未检测春化基因有Vrn-B1、Vrn-D1、Vrn-B3,二系杂交小麦亲本中是否含有其他春化基因类型有待进一步检测。
3.2 矮秆基因型和株高的关系
Rht 基因具有降低小麦植株高度,增强小麦抗倒伏能力,甚至增加小麦粮食收获指数的功能,但是不同的Rht 基因具有不同的特征,对于小麦的影响也各不相同[8]。携带Rht1(Rht-B1b)植株的平均高度大约是69.9 cm,降秆24%,Rht2(Rht-D1b)植株的平均高度大约是67.9 cm,降秆24%,当Rht1 和Rht2 矮秆基因共同作用时株高降秆60%[17]。本研究发现二系杂交小麦骨干不育系利用的主要携带矮秆基因Rht-D1b,占不育系材料的100%,二系杂交小麦不育系材料平均株高67 cm(未发表数据)。在恢复系中主要携带矮秆野生型基因Rht-B1a,均株高92 cm(未发表数据)。上述结果可能是二系杂交小麦亲本定向选择形成的。由于目前还未开发出Rht8 基因的KASP 功能标记,因此本研究尚未对Rht8 基因进行检测。
3.3 KASP 标记鉴定杂交小麦亲本的科学意义
小麦光周期不敏感品种,长日照和短日照条件下均可正常开花,其种植适宜区域广[14]。而本团队多年的研究发现,冬春杂交选育出杂交小麦品种,具有较广泛的生态适应性,其种植适宜区域较广[11],本研究发现,杂交小麦亲本主要集中于光周期敏感和冬性基因型,不利于筛选出光周期不敏感组合,因此今后在种质资源引进和利用方面,需要增加光周期不敏感型材料,而且进一步发展冬春杂交组合配置模式,以增强二系杂交小麦的生态适应性。此外,需要进一步在亲本株高性状的选育中,聚合降秆作用较强的基因,同时考虑携带不同株高基因的不育系和恢复系配制的杂交组合的性状表现,以期将二系杂交小麦株高控制在更合理的范围。
本研究结果表明可利用KASP 功能标记对二系杂交小麦亲本光周期、春化和矮秆基因的基因型进行检测,结合田间农艺性状调查,从而建立二系杂交小麦生态适应性评价体系。这为鉴定并筛选优异基因型材料,并充分利用优异种质资源,丰富二系杂交小麦亲本的遗传基础,以及创制强优势、广适性杂交小麦组合提供重要支撑。本研究进一步验证了KASP 标记可以作为一种重要的分子育种手段,提高二系杂交小麦育种效率。
