玉米BPC 家族基因的生物信息学与表达规律分析
2020-10-30米淑玲张晓燕藏金萍邢继红董金皋
周 帆,米淑玲,张晓燕,藏金萍,张 康,邢继红,董金皋
(1. 河北省植物生理与分子病理学重点实验室/河北农业大学真菌毒素与植物分子病理学实验室,河北 保定 071000; 2. 河北省沧州市运河区农业局,河北 沧州 061000)
BASIC PENTACYSTEINE(BPC)基因家族是植物特异性的转录因子家族,在基因转录调控过程中发挥重要作用[1]。拟南芥有7 个BPC 基因,分为BPC1-BPC3、BPC4-BPC6 和BPC7 3 种类型。除了BPC5 被认为是假基因外,其他6 个BPC 基因均是编码转录的激活因子或阻遏因子[1-2],与3 000多个包含GAGA 重复序列的基因结合。研究发现,BPC 家族基因存在功能冗余,单个的bpc 基因突变表型不明显,但多个bpc 基因同时突变会导致明显的发育缺陷[1-2]。BPC 可与Polycomb-Repressive Complex 1(PRC1)蛋白相互作用形成蛋白复合物,通过修饰组蛋白的尾部抑制基因的转录,进而进行染色质的重塑[3-4]。此外,BPC6 与PRC1 复合体的成员LHP1 相互作用,将LHP1 招募至GAGA结合基序上,并且显示PRC2 成员Vernalization 2(VRN2)在体内通过LHP1 与BPC6 关联[5],也能通过PRC1/2 复合物独立机制调节转录[5-6]。
BPC 的功能并非对一种发育过程或组织具有特异性。BPC 是胚珠发育基因INNER NO OUTER (INO)的调节因子[1],同时也参与种子的发育,是B3 结构域LEAFY COTYLEDON LEC2(LEC2)基因的调节因子[7]。此外,体外试验表明BPC1 的结合导致STK 启动子区的DNA 弯曲,这可能是由于BPC1 在STK 启动子中寡聚并结合多个GA 片段的能力所促进的[6,8]。
玉米中转录因子家族众多且结构复杂,Plant Transcription Factor Database(Plant TFDB)数据库显示玉米中共含有3 376 个转录因子,分别隶属于56 个转录因子家族[9]。其中大部分转录因子的研究都已取得进展,而玉米BPC 家族的研究还未及深入。本研究利用生物信息学技术,对玉米BPC 家族基因进行系统发育、基因结构、保守结构域、组织表达特性以及在玉米抵抗非生物胁迫过程中的表达规律分析;利用Real-time PCR 技术,检测玉米BPC 家族基因在不同激素处理后的表达水平,为阐明玉米BPC 家族基因的功能及其调控机制奠定基础。
1 材料和方法
1.1 试验材料
玉米自交系‘郑单958’,由河北农业大学真菌毒素与植物分子病理学实验室提供和保存。
1.2 玉米BPC 家族基因的系统发育分析
从Plant TFDB 数据库中下载拟南芥、水稻、高粱、玉米的BPC 蛋白序列,利用MEGA 7.0 构建BPC 基因的系统发育进化树[2],比对方法选取邻接法,bootstrap 值设为1 000。
1.3 玉米BPC 家族基因的基因结构分析
从玉米基因组数据库MaizeGDB(https://www.maizegdb.org/)中查询得到玉米BPC 家族中各基因的分析图,对比分析玉米BPC 家族基因的相关信息。
1.4 玉米BPC 家族基因的结构域分析
利用已知的玉米BPC 基因序列,使用Pfam(http://pfam.xfam.org/)数据库工具对其结构域进行分析,获得玉米BPC 家族基因的结构域分析图。
1.5 玉米BPC 家族基因的组织特异性表达分析
从玉米基因数据库MaizeGDB(https://www.maizegdb.org/)中下载玉米BPC 家族基因各自的gene model 表达图谱,其中MaizeGDB 数据库使用高密度NimbleGen 微阵列[10]和RNA 测序[11]可确定玉米BPC 家族的组织特异性表达情况,从eFP 浏览器可获得NimbleGen 阵列数据的表达图像[12],蛋白质在组织中不同的表达程度用不同的颜色表示,黄色表达程度低,红色表达程度高,黄色到红色表示蛋白在组织中表达量逐渐增高。
1.6 玉米BPC 家族基因在不同胁迫处理下的表达分析
从玉米数据公共平台上获得14 日龄的B73 玉米幼苗中玉米BPC 家族在热胁迫(50 ℃处理4 h)、冷胁迫(5 ℃处理4 h)、盐胁迫(300 mmol/L NaCl 浸泡20 h)、紫外线胁迫(2 h)和干旱胁迫(幼苗置于滤纸上4 h)处理下的数据,通过Heml 软件制作热图并分析。
1.7 玉米BPC 家族基因在不同激素处理下的表达分析
选取14 日龄的生长状态良好且一致的玉米自交系郑单958 幼苗,分别进行水杨酸(SA)、茉莉酸(JA)、乙烯利(ACC)处理,并于处理后的0、12、24 h 截取玉米幼茎,并用液氮速冻,试验重复3 次。利用OMEGA 植物RNA 提取试剂盒,提取玉米幼茎的总RNA,使用TaKaRa 反转录合成试剂盒,进行玉米cDNA 的合成。以反转录得到的cDNA 为模板,使用玉米BPC 家族基因的特异性引物(表1),进行荧光实时定量PCR 检测玉米BPC 家族基因的表达水平,试验重复3 次。
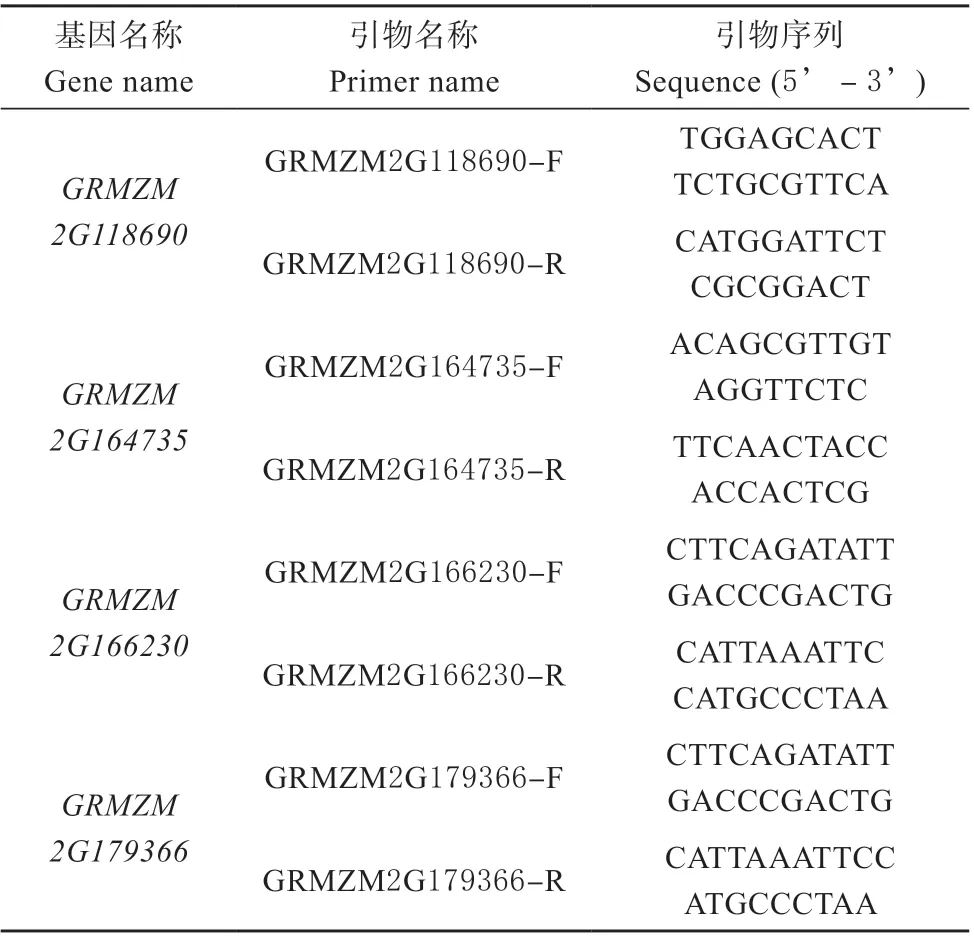
表1 试验中所用引物Table 1 Primers used in this study
2 结果与分析
2.1 玉米BPC 家族基因的系统发育分析
为了阐明玉米BPC 家族基因的系统发育关系,本研究从Plant TFDB 中获取了7 个拟南芥、4 个水稻、5 个高粱和4 个玉米的BPC 家族蛋白的相关数据,通过MEGA 7.0 使用邻接法对比得出BPC 蛋白序列进化树(图1)。进化树形成3 个亲缘关系较近的分支,分别命名I、II、III;分支I 包含GRMZM2G118690、Sobic.010G021200、LOC_Os06g04010、AT5G42520、AT4G38910、AT2G21240,分支II 包含AT2G35550,分支III 包含AT2G01930、AT1G14685、AT1G68120、LOC_Os10g02620、LOC_Os10g02509、LOC_Os10g02584、GRMZM2G164735、GRMZM2G166230、GRMZM2G179366、Sobic.002G114000、Sobic.002G114200、Sobic.002G114400、Sobic.010G056000。其 中,4 个玉米的BPC 家族蛋白中,GRMZM2G118690 属于分 支I,GRMZM2G164735、GRMZM2G166230、GRMZM2G179366 亲缘关系较近,均属于分支III 中。
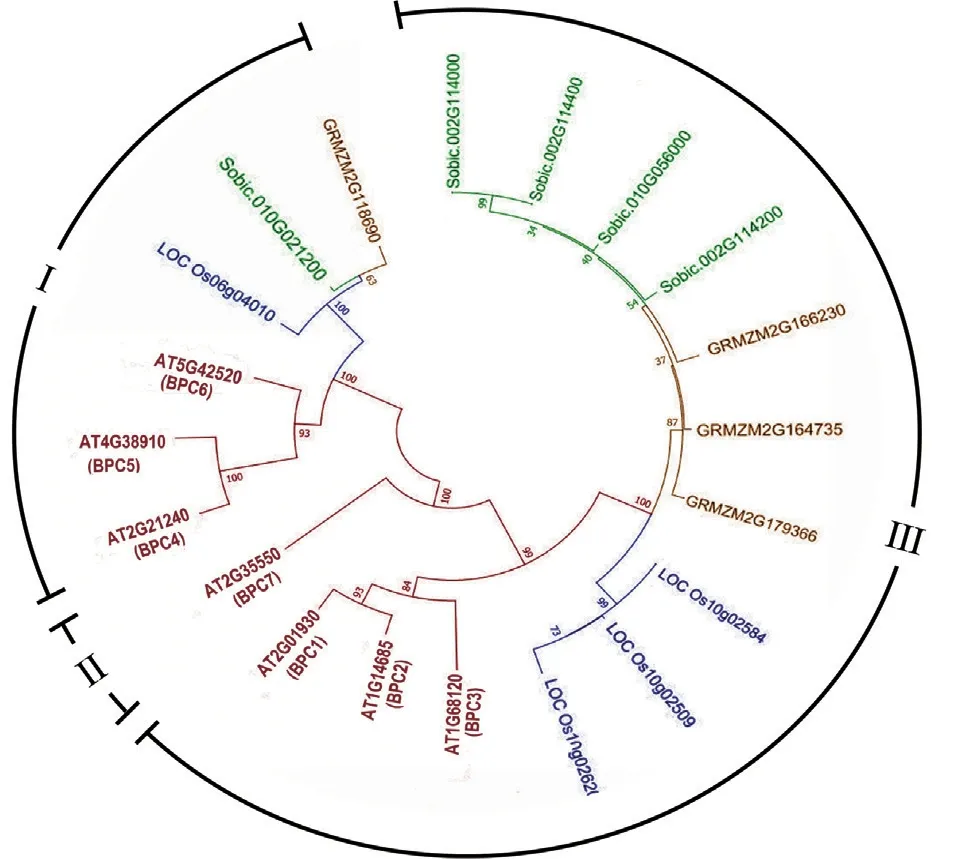
图1 玉米BPC 家族蛋白的系统发育树Fig.1 Phylogenetic trees of BPC family proteins in maize
2.2 玉米BPC 家族基因的基因结构分析
对玉米BPC 家族基因的基因结构进行分析,结果发现,玉米BPC 家族基因的内含子和外显子数量及其分布存在明显的差异。其中,在GRMZM2G118690的3 种转录本T01、T02、T03 中都含有2 个外显子和1 个内含子,3 个转录本的编码区长度一致;GRMZM2G164735 和GRMZM2G166230 的2 种 转 录本T01、T02 中均无内含子,且2 个转录本的编码区长度一致;GRMZM2G179366 的转录本T01 中含4 个外显子和3 个内含子,T02 中只含有1 个外显子,T01 编码区的长度明显长于T02(图2)。以内含子的数量为参考依据,可将玉米BPC 基因分为三类:GRMZM2G179366-T01 含 有3 个 内 含 子,为I 类 基因;GRMZM2G118690 的3 种转录本T01、T02、T03都含有1 个内含子,为II 类基因;III 类没有内含子,包 括GRMZM2G164735 和GRMZM2G166230 的2 种转录本T01、T02 以及GRMZM2G179366-T02。从基因长度上比较,GRMZM2G179366-T01 的序列最长,其次是GRMZM2G118690 的3 种转录本和GRMZM2G164735 的2 个转录本,GRMZM2G166230的2 种转录本的序列最短。
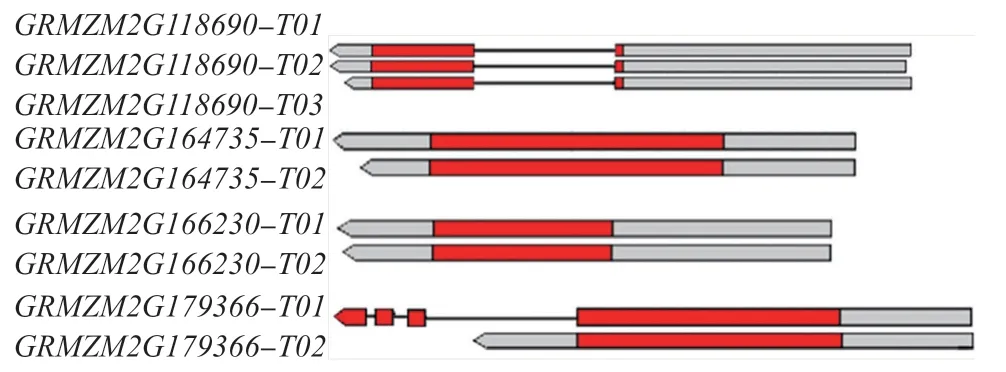
图 2 玉米BPC 家族基因的基因结构Fig.2 Gene structure of BPC family genes in maize
2.3 玉米BPC 家族基因的结构域分析
对玉米BPC 家族基因的结构域进行分析,结果发现,玉米BPC 家族基因均包含有保守的GAGA 结合结构域(GAGA_bind),但保守结构域的长度存在明显差别(图3)。其中,GRMZM2G118690 的3种转录本T01、T02、T03 均含有一段由330 个氨基酸组成的GAGA_bind 结构域,GRMZM2G164735的2 个转录本T01、T02 均含有348 个氨基酸组成的GAGA_bind 结 构 域,GRMZM2G166230 的2 个转录本T01、T02 含有184 个氨基酸组成的GAGA_bind 结构域,GRMZM2G179366 的2 个转录本T01和T02 的保守结构域GAGA_bind 由350 个氨基酸组成。GAGA 结合结构域在GRMZM2G118690、GRMZM2G164735 和GRMZM2G179366 中所占长度比例基本一致,GRMZM2G179366 中GAGA 结合结构域所占长度比例最短。

图 3 玉米BPC 家族蛋白的保守结构域Fig.3 Conserved domain of the BPC family proteins in maize
2.4 玉米BPC 家族基因的组织特异性表达分析
从MaizeGDB 数据库获得玉米BPC 家族基因的组织特异性表达数据,发现玉米BPC 家族基因呈现出相同的分布和表达模式(见图4)。
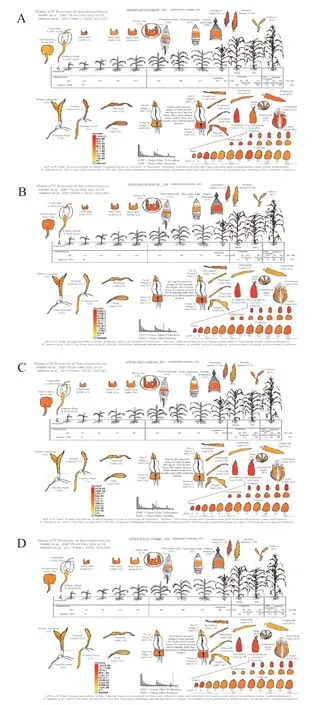
图4 玉米BPC 家族基因的组织表达特性Fig.4 Tissue-specific expression profile of BPC family genes in maize
在玉米未成熟阶段,BPC 家族的4 种蛋白GRM ZM2G118690、GRMZM2G164735、GRMZM2G 166230 和GRMZM2G179366 均在茎尖、未成熟的雄穗、叶片的叶尖、叶基以及未成熟的穗轴中的表达量较高,这些部位都含有大量的分生组织,雄穗中则含有大量减数分裂产生的雄配子,因此推测BPC 蛋白可能与细胞的有丝分裂与减数分裂有关。在成熟玉米中,BPC 家族蛋白在玉米种子的胚、胚乳和种皮中表达量较高。植物的胚与植物的遗传息息相关,胚乳则为种子的发育提供营养,因此BPC家族可能在种子发育方面发挥着重要作用。
2.5 玉米BPC 家族基因在胁迫处理下的表达水平
对玉米BPC 家族基因在热胁迫、冷胁迫、盐胁迫、紫外线胁迫和干旱胁迫处理下的表达水平进行分析,结果发现,热胁迫下,GRMZM2G118690、G R M Z M 2 G 1 6 4 7 3 5、G R M Z M 2 G 1 6 6 2 3 0、GRMZM2G179366 的表达量均明显的降低,其中GRMZM2G166230 在玉米中的表达量最低,其次是GRMZM2G164735。冷胁迫下,4 个基因的表达量均有一定程度的下降,其中GRMZM2G164735 下降的幅度最小,GRMZM2G166230 下降程度最为明显。盐胁迫下,GRMZM2G164735、GRMZM2G166230和GRMZM2G179366 的 表 达 明 显 下 调,GRMZM2G118690 的表达量没有明显变化。紫外线 胁 迫 下,GRMZM2G118690、GRMZM2G166230和GRMZM2G179366 的表达量受到明显的抑制,GRMZM2G164735 的表达量没有明显变化。干旱胁迫下,GRMZM2G166230 的表达量受到明显的抑制,其它3 个基因的表达量没有明显的变化(图5)。因此推测BPC 家族基因参与玉米对热胁迫、冷胁迫、盐胁迫、紫外线胁迫和干旱胁迫的响应过程。
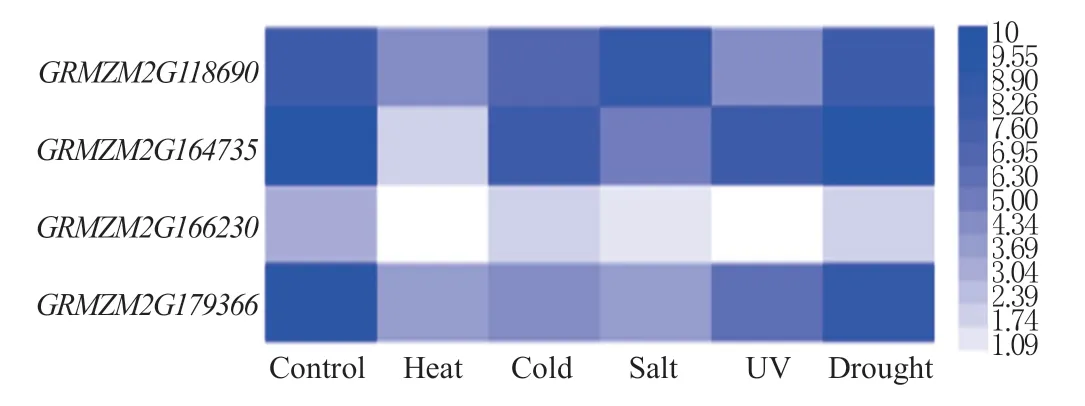
图5 玉米BPC 家族基因在胁迫处理下的表达Fig.5 Expression levels of BPC family genes in maize under stresses
2.6 玉米BPC 家族基因在激素处理后的表达水平
水杨酸、茉莉酸、乙烯利处理14 日龄的玉米自交系郑单958幼苗,对玉米BPC家族基因GRMZM2G118690、 GRMZM2G164735、GRMZM2G166230、GRMZM2G179366 的表达量进行qRT-PCR 检测(见图6)。
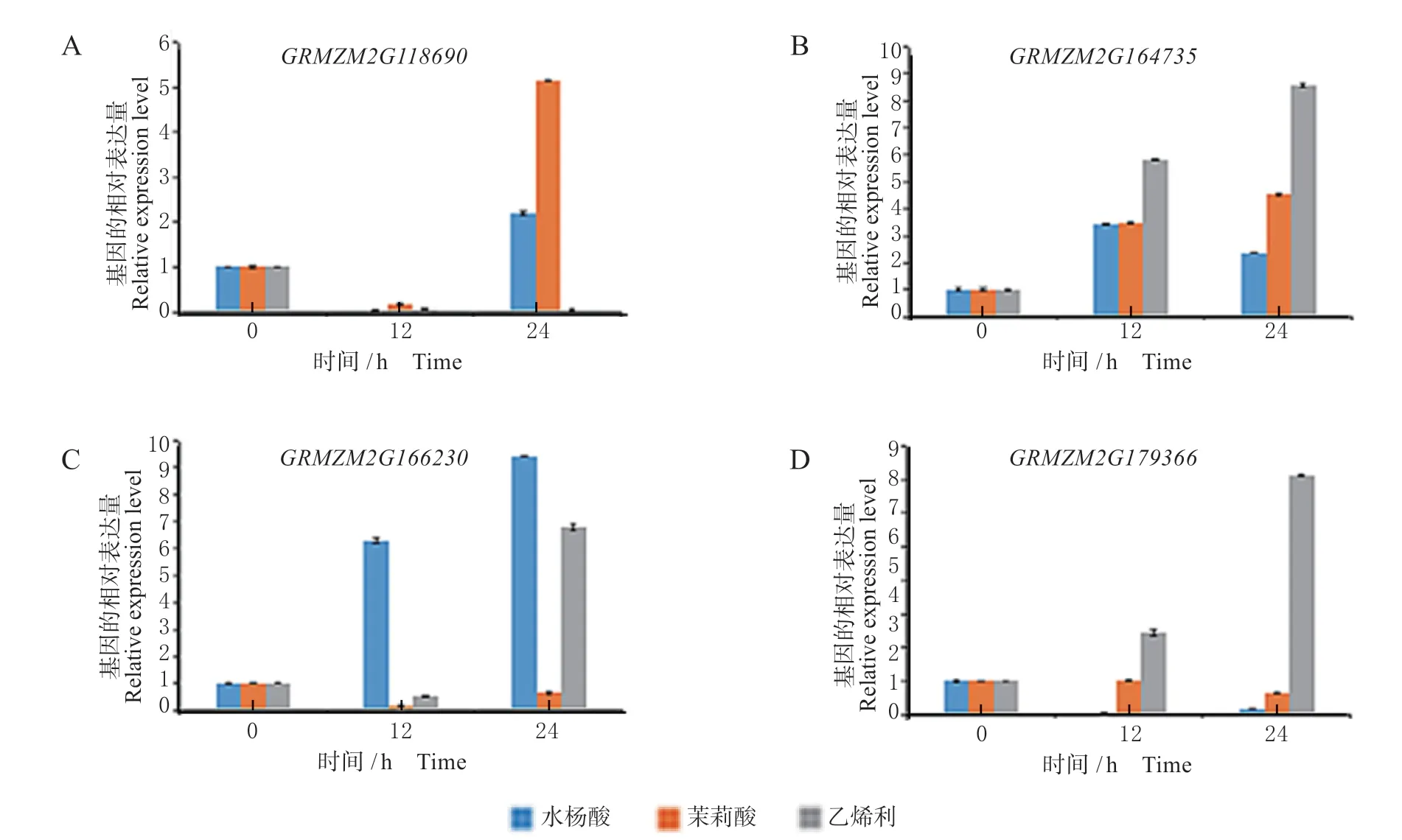
图6 SA、JA 和ACC 处理下玉米BPC 家族基因的表达水平Fig.6 Expression levels of BPC family genes in maize treated with SA, JA and ACC
结果发现,GRMZM2G118690 在水杨酸、茉莉酸、乙烯利处理12 h 后,表达量均明显下降,处理24 h 后,在水杨酸和茉莉酸处理的玉米样品中表达量则出现增高,在茉莉酸处理下,增幅更为明显。GRMZM2G164735 在水杨酸、茉莉酸、乙烯利处理的样品中,在处理后的12 h 和24 h 表达量均增高,在乙烯利处理24 h 内玉米样品中的表达量最高。GRMZM2G166230 表达量在水杨酸处理后24 h 内持续增高;茉莉酸处理后,GRMZM2G166230 的表达量在处理24 h 内有所下降,其中处理的前12 h 内,下降趋势最为明显。GRMZM2G179366 在乙烯利处理的样品中,表达量变化明显上升,而水杨酸和茉莉酸处理后,GRMZM2G179366 的表达量则显示出下降的趋势。
3 讨论与结论
BPC 家族基因参与调控植物的多种生理过 程[1-2],尽管BPC 家族是研究时间较短的植物特异性转录因子家族[13],但目前研究已经发现,其可以充当基因表达的双向调节因子发挥作用[1,6,8,13]。在真核生物中,多种转录因子通常共同作用以调节基因表达[14-16]。鉴于BPC 调控多种发育过程,它们很可能依赖于其他转录因子的共同作用调控靶基因的特异性表达。BPC6 蛋白的靶基因与响应细胞分裂素而富集的B 型ARR10 的靶基因之间存在明显的重叠[17]。另外,BPC 可能与B 型ARR 和/或CRF共同作用,以响应细胞分裂素信号传导来调节特定基因[17-19]。ARR10 可与BPC1 和BPC2 的启动子结合,并且BPC1 响应细胞分裂素而被诱导,因此BPC 和B 型ARR 之间可能存在正反馈回路。
本研究发现,玉米BPC 家族蛋白与水稻的BPC家族蛋白的亲缘关系更为密切,因此,以水稻的同源转录因子为参照,进行玉米BPC 家族基因的研究,分析进化过程中的变异,推测玉米的物种起源或进化途径。同时基因结构和蛋白保守结构域与基因的功能有着紧密的联系,以水稻和玉米的亲缘关系为基础,以基因结构和蛋白保守结构域的分析为辅助,可以更直接地对玉米BPC 家族基因进行功能分析。
研究表明,BPC 调节细胞分裂素调节基因的一个子集[18],这与BPC 家族蛋白的分布相一致。从MaizeGDB 中得到的玉米BPC 家族在不同组织当中表达量的分析发现:4 种BPC 蛋白均在茎尖、穗尖和未成熟玉米籽粒中的表达量较高,推测其功能可能与细胞分裂和促进生长发育有关。非生物胁迫中,GRMAM2G179366 在热处理、冷处理、盐处理、紫外线处理中,表达量均有明显的降低趋势,推测其参与植物抵抗逆境的过程。水杨酸、茉莉酸和乙烯利处理后,4 种基因表达量的不同变化表明,玉米BPC 家族的不同基因可能参与不同激素介导的免疫过程。
本研究运用生物信息学技术结合Real-time PCR技术,对玉米BPC 家族进行了系统发育、基因结构、蛋白保守结构域、组织表达特异性、非生物胁迫及激素处理下的表达水平进行了分析,推测玉米BPC家族可能与有丝分裂、减数分裂、种子发育以及植物体抗病抗逆过程有关,为玉米BPC 家族基因的功能及其调控机制的研究指明了方向。
