白菜型冬油菜萌动种子低温春化的生理生化特征
2020-03-30徐春梅邹娅刘自刚米文博徐明霞董小云曹小东郑国强方新玲
徐春梅,邹娅,刘自刚,米文博,徐明霞,董小云,曹小东,郑国强,方新玲
(甘肃农业大学农学院/甘肃省油菜工程与技术研究中心/甘肃省干旱生境作物学重点实验室/甘肃省作物遗传改良与种质创新重点实验室,兰州 730070)
0 引言
【研究意义】中国北方冬季严寒漫长,属于传统一年一熟制区,白菜型冬油菜是该区主要越冬作物。白菜型冬油菜具有优异抗寒性和较强的感温性,强冬性冬油菜幼苗完成春化所需时间长,南繁或室内加代存在较大困难,使冬油菜育种进程极为缓慢,不适应高效育种的需求[1-2]。本研究探讨白菜型冬油菜萌动种子低温春化的可能性,萌动种子如能感受低温完成春化,即可简便地对大规模育种材料等进行春化处理,对缩短冬油菜育种周期、提高育种效率具有重要意义。【前人研究进展】春化作用是冬性植物感受环境低温机体由营养生长转向生殖生长,最终诱导植株成花的过程[3]。植物春化分为绿体春化和种芽春化2种方式,绿体春化需要植株生长至一定大小,此时幼苗才可感受低温通过春化,缺点是占用空间大,费时费力;种芽春化是在种子萌动状态下进行春化处理,可同时对大量种子进行处理,要求设备简单,省时省力[4],能满足对育种材料繁育加代的需求。研究表明冷藏柜和生长室4℃低温对油菜萌动种子进行春化处理,可实现冬性和半冬性品种的加代;感温性不同的大白菜品种通过春化所需低温处理时间不同,冬性越强低温处理时间越长;随低温春化时间的延长,冬性材料现蕾的时间逐渐提早,开花株率增加;半冬性甘蓝型油菜试管苗置于6—8℃光照培养箱处理32 d,所有植株均可通过春化[5-10]。青花菜[11]和小麦[12]萌动种子低温处理后,青花菜促进其花芽分化;小麦抽穗提前,粒重增加。通过在高纬度高海拔地区早春播种不同感温性白菜型冬油菜育种材料,发现显蕾不抽薹、抽薹不开花、结实不正常等明显不完全春化现象,其生育期进程、植高、结籽率等经济性状与春化率密切相关。另外,激素、硝酸还原酶、胞内生理状态等均与植物低温春化状态和程度有关[13-15]。【本研究切入点】白菜型冬油菜[16]的春化研究中,采用5—6叶龄的幼苗进行自然越冬和低温培养,易受空间限制,繁育加代困难。而萌动种子低温春化能够加快繁育速度,但目前对不同感温性冬油菜品种萌动种子低温春化的研究较少,且具体春化时间不明确。【拟解决的关键问题】本研究以不同感温性白菜型冬油菜品种为材料,萌动后低温(4℃)春化处理不同时间(20—60 d),研究萌动种子能否感受低温完成春化,对其生理生化响应、激素水平、萌动种子播种后株高、结实性差异进行研究,明确不同感温性白菜型冬油菜春化所需低温时长,探讨北方白菜型冬油菜利用人工低温春化加代的方法,为加快育种进程提供依据。
1 材料与方法
1.1 试验材料
材料为白菜型冬油菜陇油7号、陇油9号和天油4号的萌动种子,均由甘肃农业大学提供;陇油7号、陇油9号和天油4号在兰州新区气候条件下的越冬率分别为97.6%、89.3%和80.2%,冬性强弱顺序为陇油7号>陇油9号>天油4号。
1.2 种子的处理方法
试验于2018年5月在甘肃农业大学干旱生境作物学重点实验室进行,选取饱满均匀、大小一致的陇油7号、陇油9号和天油4号白菜型冬油菜种子,用70%酒精消毒1 min,无菌水冲洗3遍,再用0.01%氯化汞消毒5 min,无菌水冲洗3遍,播于铺有两层滤纸的玻璃培养皿中,在光照培养箱(25℃)中萌动,待种子露白后,4℃低温处理。5月10日开始,处理20 d后取出一批萌动种子;之后,每隔10 d取出一批萌动种子,直至低温春化处理60 d,共5个处理时间(20、30、40、50和60 d),0 d为对照。每处理的萌动种子均分成2份,一份播种于装满营养土的盆钵中,用于观察统计春化率、测定表型性状等;另一份萌动种子液氮速冻,用于测定激素等生理指标。试验均重复3次。
1.3 测定指标和方法
1.3.1 表型统计 播种于营养钵中的萌动种子,待幼苗生长15 d后,需每天观察生育期进程,现蕾即看作通过春化,计算春化率,公式为:春化率=(现蕾株数+抽薹株数+开花株数)/总株数×100%[17];初花期、成熟期各测量一次株高,成熟期测定结实率等经济性状。
1.3.2 生理指标的测定 按照邹琦[18]的方法测定叶片生理生化指标,采用离体法测定硝酸还原酶(nitrate reductase,NR)活性,NBT光还原法检测超氧化物歧化酶(superoxide dismutase,SOD)活性,愈创木酚法检测过氧化物酶(peroxidase,POD)活性,紫外吸收法检测过氧化氢酶(catalase,CAT)活性,考马斯亮蓝G-250法检测可溶性蛋白(soluble protein,SP)含量,蒽酮法检测可溶性糖(soluble sugar,SS)含量,2-硫代巴比妥酸法测定丙二醛(malondialdehyde,MDA)含量。
1.3.3 内源激素的测定 采用液相色谱质谱联用仪Agilent LC-MS(美国安捷伦公司)测定赤霉素(gibberellin,GA3)、生长素(auxin,IAA)和脱落酸(abscisic acid,ABA)[19-20]。
色谱参数:色谱柱:Agilent SB-C18(50 mm×2.1 mm,1.8 μm);流动相:A泵溶液:0.1%甲酸水溶液,B泵溶液:甲醇;流速:0.3 mL·min-1;柱温:40℃;进样量:2 μL。质谱参数:在MRM模式下,用负离子模式,毛细管电压为4 kV,干燥器流量为11 L·min-1,干燥器温度为350℃,MS参数见表1。
1.4 数据处理
运用Excel 2013和SPSS19统计软件进行数据整理和分析。
2 结果
2.1 春化时间对白菜型冬油菜现蕾开花的影响
随低温春化处理天数的增加,白菜型冬油菜的春化率基本呈上升趋势;未进行低温春化处理的萌动种子形成的植株(CK),未见现蕾开花单株,各品种春化率均为0。陇油7号萌动种子低温春化处理20 d所形成的植株,其春化率仅为25.81%,当春化时间增加至60 d时,其春化率达到86.96%;陇油9号、天油4号均呈相似趋势(图1和表2)。
不同感温性白菜型冬油菜品种萌动种子通过春化(春化率>95%)所需低温处理时间明显不同。陇油7号所需低温处理时间最长,为76.9 d,陇油9号为54.0 d,天油4号最短为39.4 d。天油4号萌动种子低温处理30 d形成植株春化率达到93.10%;而陇油9号和陇油7号春化率仅为56.52%和19.23%(表2)。此外,白菜型冬油菜感温性(春化所需低温天数)与抗寒性(越冬率)呈显著正相关(r=0.988),表明白菜型冬油菜冬性越强的品种其抗寒性也越强。

表1 不同内源植物激素在MRM模式下的MS参数Table 1 MS/MS conditions for different endogenous plant hormones

图1 不同春化时间对白菜型冬油菜表型的影响Fig.1 Effect of vernalization time on phenotype of B.rapa

表2 不同春化时间对白菜型冬油菜春化率的影响Table 2 Effect of vernalization time on the number of flowering plants of B.rapa (%)
2.2 春化时间对白菜型冬油菜株高的影响
随春化处理时间的增加,白菜型冬油菜初花期和成熟期株高均呈升高趋势(表3)。陇油7号萌动种子低温春化处理20 d时初花期、成熟期平均株高仅为5.68和15.16 cm;60 d时初花期、成熟期平均株高可达29.80和68.26 cm。低温春化处理相同时间时,不同品种初花期和成熟期株高总体表现为冬性越弱,株高越高;低温春化处理20—60 d,天油4号株高均明显高于陇油9号和陇油7号。

表3 春化时间对白菜型冬油菜株高的影响Table 3 Effect of vernalization time on plant height of B.rapa
2.3 春化时间对白菜型冬油菜结实性能的影响
随着低温春化处理时间增加,白菜型冬油菜植株的一次分枝数、单株角果数、角果长度、角果粒数、单株产量等性状均呈升高趋势(表4),表明随着低温春化时间增加,可明显提高植株结实能力。如陇油7号萌动种子低温春化20 d,角果数37个,角粒数12.33粒,平均单株产量0.74 g;当低温春化时间增至60 d时,植株角果数增加54.9%,角粒数、单株产量均增加73.0%;萌动种子低温春化时间增加后,陇油7号植株结实能力大量提高。
各品种在低温春化处理前期,通过春化植株的结实能力在品种间差异表现明显;随春化处理时间延长,品种间结实能力的差异逐渐减小。低温春化20—30 d,陇油9号植株分枝数显著高于陇油7号和天油4号,延长至40 d以上,陇油9号分枝数仍略高于其他2个品种,但品种间差异均未达到显著水平。表明春化不完全的植株结实能力受其春化程度的影响较大,完全春化植株结实能力主要取决于品种特性。
2.4 白菜型冬油菜不同感温性品种表型特征与春化率的相关性分析
不同感温性白菜型冬油菜低温春化处理后,其春化率与平均初花株高、平均成熟株高、相对株高、一次分枝数、单株角果数、角果长度、角果粒数、单株产量呈极显著正相关(P<0.01);表明低温春化处理的白菜型冬油菜萌动种子春化程度,与之后其所形成植株的生长发育、结实能力等密切相关;且各表型性状之间也呈极显著正相关(P<0.01),表明春化率能够显著的影响表型特征(表5)。其中,春化率与初花期和成熟期株高的相关系数最大,分别为0.947和0.985,说明春化率与株高关联密切;单株产量与单株角果数、角果长度、角果粒数相关系数达0.980,说明单株产量受单株角果数、角果长度、角果粒数影响。
2.5 春化处理对白菜型冬油菜萌发幼苗硝酸还原酶活性的影响
随春化处理时间的增加,白菜型冬油菜硝酸还原酶活性总体呈降低趋势(图2)。低温春化处理20 d,陇油7号、陇油9号和天油4号NR活性显著升高,相比对照分别增加86.48%、51.98%、95.02%;之后随低温春化时间延长(30—50 d),各品种NR活性明显下降,并与对照始终保持在同一水平;春化时间增至60 d时,各品种NR活性相比对照均显著降低。

表4 不同春化时间对白菜型冬油菜结实性的影响Table 4 Effect of vernalization time on seed set of B.rapa
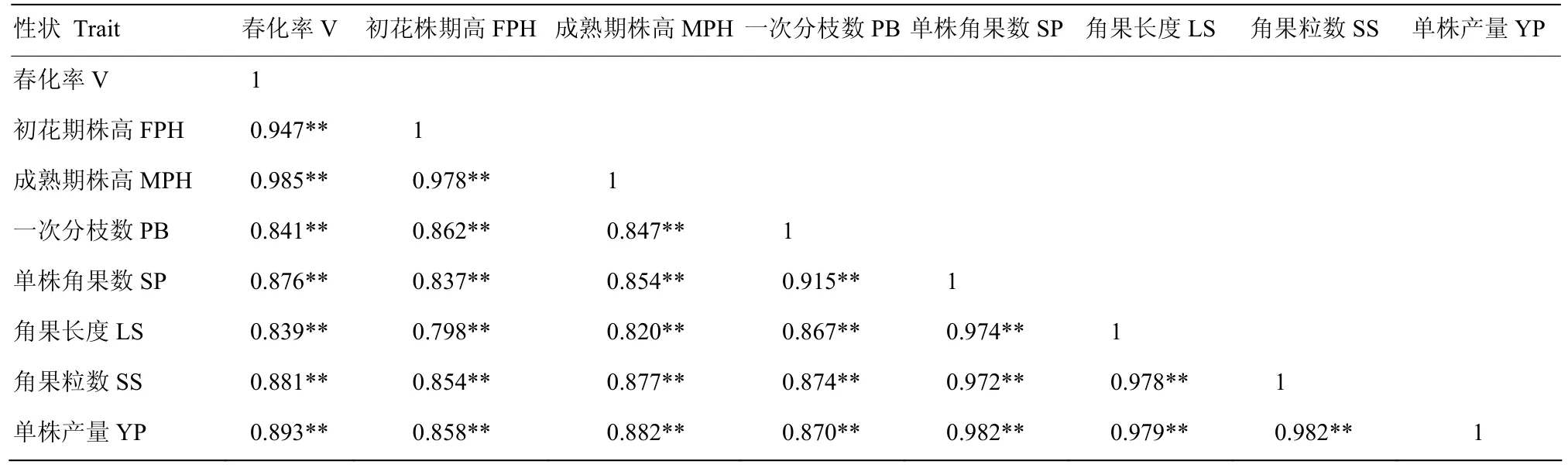
表5 不同冬性白菜型油菜春化率、表型特征之间的相关性Table 5 Correlation between vernalization rate and phenotypic traits of B.rapa
2.6 春化处理对白菜型冬油菜萌动种子抗氧化酶活性的影响
SOD可以清除植物体内积累的O2-,与POD、CAT共同作用生成H2O,减轻逆境对植物的伤害。随低温春化时间的延长,陇油7号、陇油9号、天油4号SOD活性呈先逐渐升高后降低的趋势(图3-a);春化处理0 d(CK)时,供试品种SOD均处于较低水平,春化处理40 d时,各品种SOD活性达到最大值,与对照相比,分别增加53.6%、52.1%和41.8%,强冬性品种增加幅度高于冬性品种,说明强冬性品种对低温春化处理的耐受性更强;春化处理至60 d时,天油4号SOD活性较陇油7号、陇油9号最低,春化处理时间过长导致天油4号O2-积累,萌动种子受害。

图2 春化处理对白菜型冬油菜萌动种子硝酸还原酶活性的影响Fig.2 Effects of low temperature vernalization on nitrate reductase activity of B.rapa
随低温春化时间的延长,陇油7号、陇油9号、天油4号POD酶活性呈先逐渐增加后降低的趋势(图3-b);春化处理0 d(CK)时,各品种POD均处于较低水平,春化处理40 d时POD活性达到最大值,相比对照分别增加110.02%、78.29%和100.2%,达显著水平(P<0.05),春化处理50—60 d,各品种POD酶活性下降趋势明显。POD活性与SOD活性变化一致。
随春化时间的增加,各白菜型油菜CAT活性随春化时间增加而逐渐下降(图3-c);在春化处理40 d时稍有波动,但整体呈下降趋势,与对照相比,达显著水平(P<0.05)。MDA含量高低可反映细胞膜脂过氧化的水平和膜受伤害的程度[21]。随低温春化时间的增加,各白菜型冬油菜MDA 含量逐渐升高,春化处理后期稍有下降,但相比对照均增加显著(图3-d);陇油7号MDA含量在春化处理50 d达到最大,相比对照增加52.12%,陇油9号和天油4号均在春化处理40 d达到最大值,相比对照分别增加81.98%和173.93%,达显著水平(P<0.05),MDA含量增量陇油7号<陇油9号<天油4号,说明天油4号对低温春化处理更敏感。
2.7 低温春化处理对白菜型冬油菜萌发幼苗渗透调节物质及质膜透性的影响
低温春化过程中,随春化时间的增加,白菜型冬油菜各品种SP、SS含量呈先增加后降低的趋势(图4-a)。陇油7号和陇油9号SP含量在春化处理30 d达到峰值,较对照分别增加24.65%和35.92%,春化处理中期SP含量达到相对平衡的状态,60 d SP含量下降;天油4号在春化处理20 d达到峰值,较对照增加13.34%,40—60 d呈下降趋势。
陇油7号萌动种子SS含量在春化处理30 d时达到峰值,之后呈波动变化(图4-b),与对照相比,各处理时间SS含量升高,达显著水平(P<0.05);陇油9号SS含量在春化处理20 d达到最大值,天油4号SS含量在春化处理30 d时达到最大,40—60 d时SS含量下降显著。

图3 春化处理对白菜型冬油菜萌动种子抗氧化酶活性的影响Fig.3 Effect of low temperature vernalization on antioxidant enzyme activity of B.rapa
2.8 低温春化处理对白菜型冬油菜萌发种子植物激素的影响
随春化时间的增加,白菜型冬油菜不同感温品种GA3含量的变化趋势不同;低温春化处理后,与对照相比,陇油7号、陇油9号萌动种子中GA3含量明显下降;而天油4号萌动种子在春化处理20—30 d后GA3含量明显升高,30 d时相比对照增加14.51%;随后萌动种子中GA3含量有所下降(图5-a)。
低温春化处理白菜型冬油菜萌动种子后,与对照相比,不同感温品种的IAA含量均有所增加。陇油7号春化处理后IAA含量相比对照增量基本一致;陇油9号和天油4号IAA含量呈逐渐升高后稍有下降的趋势,在春化50 d 时IAA增幅最大,分别增加39.9%和197.0%(图5-b)。
随低温春化时间的增加,白菜型冬油菜各品种萌动种子中ABA含量呈先增加后降低的趋势,与对照相比,陇油7号的ABA含量在春化20 d达到最大,增加94.23%,之后逐渐下降;陇油9号、天油4号在春化40 d时ABA含量达到最大,分别增加34.87%和61.24%,之后迅速下降至对照水平(图5-c)。
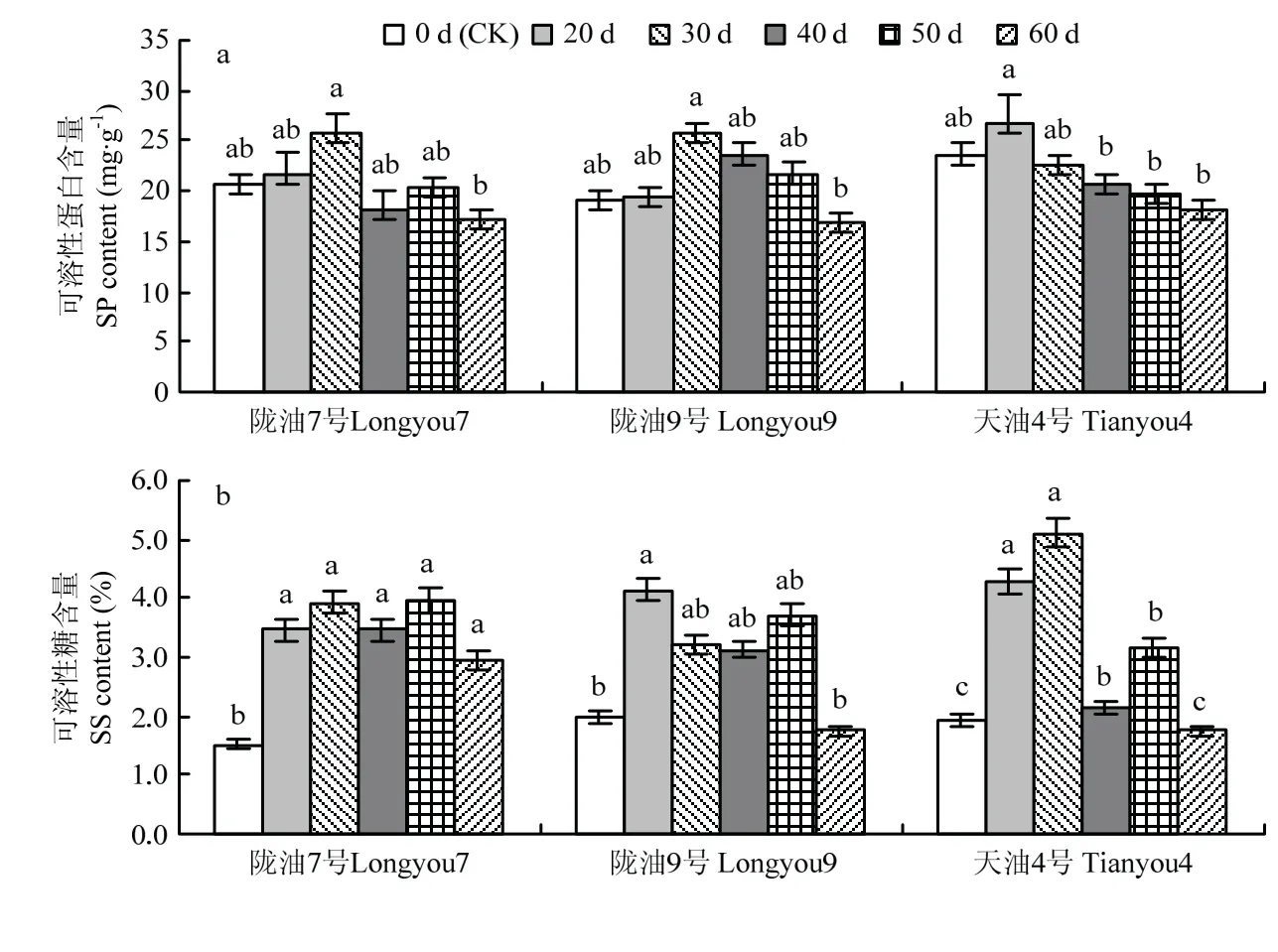
图4 低温春化处理对白菜型冬油菜萌动种子渗透调节物质及质膜透性的影响Fig.4 Effects of low temperature vernalization on osmotic adjustment substances and plasma membrane permeability of B.rapa

图5 低温春化处理对白菜型冬油菜萌动种子植物激素的影响Fig.5 Effects of low temperature vernalization on plant hormones of B.rapa
2.9 生理生化指标与春化率的相关性分析
采用相关分析计算各生理指标(NR、SOD、POD、CAT、MDA、SP、SS、GA3、IAA、ABA)与春化率的相关系数(表6),春化率与CAT呈极显著负相关(P<0.01),与MDA呈极显著正相关(P<0.01)。NR与SOD显著正相关(P<0.05),与SP呈极显著正相关;SOD与POD、ABA呈显著正相关,POD与MDA呈极显著正相关,CAT与MDA呈极显著负相关,SP与GA3呈显著正相关。

表6 不同冬性白菜型油菜春化率、生理指标之间的相关性Table 6 Correlation between vernalization rate and physiological indexes of B.rapa
3 讨论
3.1 白菜型冬油菜萌动种子低温春化的表型特征
成花过程是植物发育的重要环节,是繁衍后代群体、维持种群稳定的重要生存策略。多数温带植物都需感受一定时间的低温,才可完成茎尖分生组织花芽的分化转化,形成花芽原基实现开花结实的过程,称为春化现象。一般而言,植物低温春化对温度的敏感期是种子萌动至幼苗发育的某一时期。依据低温春化作用敏感时期,可将春化作用分为萌动种子春化型和绿体春化型[22]。萌动种子春化型植物在种子萌动期即可感受低温,完成春化作用,若需要人工春化处理时,即可很方便的处理大量种子材料,这对具有感温特性,特别是强冬性作物开展繁育加代、缩短育种周期非常有效。中国北方白菜型冬油菜属于强冬性作物类型,本研究以3个不同感温品种为材料,探讨了萌动种子感受低温春化的生理过程,及春化后植株表型发育特性,结果表明,白菜型冬油菜种子萌动期可以感受低温春化作用,依据冬性强弱,通过春化作用所需的低温时间不同,强冬性陇油7号、陇油9号、冬性品种天油4号完全通过(春化率>95%)春化需要时间分别为76.9、54.0和39.4 d。随春化处理时间的增加,陇油7号、天油4号、陇油9号初花株高、成熟期株高、一次分枝数、单株角果数、角果长度、角果粒数、单株产量逐渐增加,各指标与春化率极显著相关,且各表型之间也极显著相关,表明春化时长对株型、结实等表型有较大影响,这与杨国浪等的研究结果一致[23]。
3.2 白菜型冬油菜萌动种子低温春化的生理生化特征
硝酸还原酶是一个对温度敏感的诱导酶[24]。研究表明低温诱导可使植物体内NR酶活性升高,不同感温性白菜型冬油菜在春化处理过程中NR活性呈先升高后逐渐下降趋势,这与以玉米为材料的研究结果一致[25]。逆境下植物大量积累活性氧等自由基,加剧膜脂过氧化从而导致膜系统受损。抗氧化酶系可有效清除这类自由基,改善胞内生理状态,提升植物对逆境的适应能力。本研究中白菜型冬油菜萌动种子低温春化处理初期SOD、POD活性均有显著提高,这很可能是种子对春化低温做出的应激反应;同时,种子内MDA含量呈升高的趋势,表明在低温逆境下种子胞内氧自由基产生量,超过抗氧化酶的清除能力,积累活性氧等自由基氧化攻击膜脂等还原物质,其产物MDA在胞内积累,导致膜系统受损。低温春化40 d时,SOD和POD活性出现峰值,之后酶活性均有所降低,可能是由于在春化过程中,低温明显抑制了萌动种子代谢活动,其胞内SOD和POD等抗氧化酶系活性随之下降,这与闫利恒等[26]的研究一致。在低温春化期间,CAT活性呈逐渐降低的趋势,这与KAR等[27]、李琦瑶等[28]研究结果吻合。
渗透调节物质能增加细胞质浓度,降低渗透势,维持细胞膨压,有利于保持水分和细胞各种生理过程的正常进行[29]。SP、SS等物质在稳定细胞内生物大分子结构等方面具有重要作用。本研究结果表明,在春化过程中SP和SS含量呈先升高后降低的趋势,处理初期(20—30 d),其萌动种子SP和SS含量比对照明显升高;春化处理中期(40—60 d),其含量明显下降;表明长时间的低温春化使萌动种子渗透调节能力减弱;与菜用豌豆和花生的研究结果一致[30-31]。
激素是植物通过次生代谢途径产生的,并能从产生部位转运至作用部位,在低浓度下就具有明显生物学效应的内源微量物质。GA3是最早被认为是植物春化激素,在低温条件下GA3、ABA等激素含量增加[32]。冬油菜萌动种子低温春化处理后,内源激素水平发生显著变化,随春化时间延长,陇油7号和陇油9号GA3含量呈下降趋势;春化30 d,天油4号GA3含量显著增加,其他处理GA3含量与对照在同一水平。IAA含量略微升高,但变化不显著;ABA含量升高,且强冬性陇油7号ABA含量明显高于陇油9号和天油4号,这与周碧燕等[33]和王兴等[34]研究结果基本一致。ABA被认为是一种逆境激素,在春化处理过程中,萌动种子ABA含量变化幅度较大,这可能与ABA在抵御冷胁迫发面发挥的作用有关。GA3被认为在在冬性植物春化过程中发挥正调控作用,本研究结果显示,冬油菜萌动种子春化过程中GA3含量明显下降,表明GA3在萌动种子春化过程中可能并未发挥调控作用,萌动种子春化途径是否有别于绿体春化途径,值得进一步研究。
4 结论
白菜型冬油菜萌动种子可感受环境低温完成春化作用,冬性越强,完成春化所需的低温(4℃)时间越长。陇油7号萌动种子完成春化(春化率>95%)需低温处理76.9 d,陇油9号、天油4号分别为54.0和39.4 d。低温春化过程中,白菜型冬油菜萌动种子生理状态变化与其春化程度密切相关,并显著影响其后植株生长发育与结实性能。低温春化处理萌动种子可用于白菜型冬油菜的加代繁育等育种实践工作。
