一个大麦矮秆突变体m1062 的表型鉴定和遗传分析
2020-03-30罗忠跃谢思宇贾巧君
罗忠跃,谢思宇,贾巧君
(浙江理工大学生命科学与医药学院/浙江省植物次生代谢调控重点实验室,浙江杭州310018)
大麦是全球栽培的第四大禾谷类作物,集啤用、饲用和食用于一体。2002 年以来,我国成为世界第一啤酒生产和消费大国,大麦需求量逐年攀升。与其他作物相比,大麦由于茎秆的机械组织不发达,纤维化程度低,在高水肥栽培条件下,大麦易出现倒伏现象[1]。倒伏不但造成大麦减产,而且间接影响麦芽品质[2]。20 世纪60—70 年代以来,大麦育种家先后选育出矮秆品种,减少了大麦倒伏,提高了大麦产量。
大麦矮化育种的基因主要是sdw1/denso、ari-e.GP、uzu1以及它们的等位基因。在美国、加拿大和澳大利亚,矮秆品种的选育主要依赖sdw1基因[3-4];大部分欧洲矮秆啤酒大麦选育依赖于sdw1的等位基因denso[5-6]。目前,sdw1/denso基因已克隆,其与水稻绿色革命基因sd1同源,编码赤霉素合成途径关键基因GA20 氧化酶,参与赤霉素合成[6-7]。通过开发sdw1/denso分子标记并对我国大麦矮秆品种检测发现,我国甘肃、云南等地选育的大麦品种也携带sdw1/denso基因[8]。此外,矮秆品种Golden Promise携带的矮秆基因命名为ari-e.GP[9],Golden Promise及其衍生系也是欧洲尤其是英国啤酒大麦育种的重要矮源之一[10]。基因克隆发现该基因编码G 蛋白γ亚基,其在第1508、1509 位点间插入1 个碱基A 导致翻译提前终止,造成该基因功能缺失,植株表现矮秆[11]。
在我国,矮秆育种使大麦株高降低20~25 cm,产量增长4.7 倍[1],显示矮秆品种在大麦增产上的巨大作用。但我国大麦育种中矮源单一,主要是尺八大麦、萧山立夏黄、沧州裸大麦和矮秆齐,且这些品种的矮秆基因都是uzu1基因[1]。单一的矮源容易导致品种遗传多样性的丧失[12],因此有必要筛选、诱变创制大麦矮秆资源,丰富我国大麦矮秆资源的多样性。
本研究从辐照诱变啤酒大麦品种秀麦3 号的M2 代中筛选到1 份矮秆突变体m1062,并对该突变体的表型特征、色素含量、产量和遗传特点进行分析,以期为该矮秆突变体的育种应用、基因克隆和功能研究奠定基础。
1 材料与方法
1.1 试验材料
矮秆突变体m1062来源于啤酒大麦品种秀麦3 号的60Co-γ 射线辐射诱变。以突变体m1062为母本,秀麦3 号为父本杂交,获得F1、F2和F3代。以中高秆品种浙大9 号为母本,m1062为父本获得杂交后代F1和F2。
1.2 田间试验和农艺性状测定
将秀麦3 号和突变体m1062播种于田间,2015、2016 年连续2 年测定其抽穗期、株高、茎节长、穗长、芒长、旗叶长等性状。将m1062/秀麦3 号的杂交种子F1点播于试验田,成熟后种子混收获得F2。m1062/秀麦3 号组合的F2种子及其亲本播种于试验田,分别在苗期、抽穗期和成熟期调查其生长特性、株型和株高,成熟期收获高秆和矮秆株系各100 株。对F2代高秆株系进行表型鉴定,确定获得纯合的高秆株系。秀麦3 号、m1062突变体和m1062/秀麦3 号组合的F3代高秆纯合单株和矮秆纯合单株点播于田间,调查群体及亲本的每穗粒数、穗数、百粒质量、单株质量等重要农艺性状,分析矮秆基因的多效性。采用t- 检验分析各性状的统计数据,检验野生型、突变体及群体后代间的显著性差异。
1.3 叶绿体色素含量测定
大麦播种6 周后,分别取秀麦3 号和突变体m1062植株的最新全展开叶,参照赵绍路等提取叶片叶绿素和类胡萝卜素的方法[13],用分光光度计测定665、649 和470 nm 下的吸光值,计算叶绿素a、叶绿素b 和类胡萝卜素的含量,重复3 次。采用t-检验分析野生型和突变体间的显著性差异。
1.4 遗传分析
将浙大9 号/m1062组合的F1播种于田间,观察其株型、株高、芒长等特征,F1表现为中高秆,没有出现矮秆表型。F1混合收获后,F2点播于田间,随机调查其株高分离情况,利用卡方检验分析其遗传方式。
2 结果与分析
2.1 突变体的表型特征
田间试验结果(表1、图1)表明,m1062矮秆突变体的株高较野生型秀麦3 号显著降低,2 年试验株高分别降低26.77、28.11 cm,降低幅度分别为34.2%、36.4%。但矮化效应并不在于减少茎节数,而是通过减少各茎节长度来降低株高。矮秆突变体m1062第1 茎节长度较秀麦3 号分别降低1.19、0.67 cm,降低幅度分别为26.6%、21.1%;第2 茎节长度分别降低2.56、2.98 cm,降低幅度分别为35.4%、38.5%;第3 茎节长度分别降低3.58、3.94 cm,降低幅度分别为33.9%、35.4%;第4 茎节长度分别降低6.54、6.59 cm,降低幅度分别为36.8%、37.2%;穗下茎长度分别降低12.81、11.60 cm,降低幅度分别为38.0%、37.2%。各茎节长度都降幅明显,穗下茎长度和第4 茎节长度降低幅度较大,是植株矮化的主要原因。矮秆突变体在降低株高的同时,抽穗期比野生型晚5~6 d。此外,矮秆突变体m1062的穗长、芒长和旗叶长都显著降低,但旗叶宽的变化相对较小。
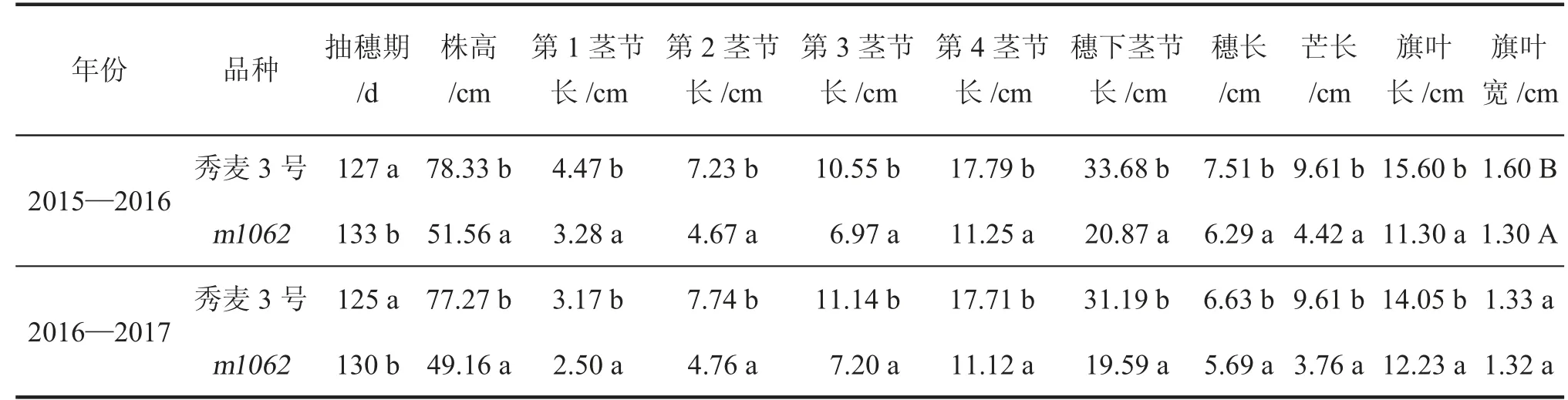
表1 野生型秀麦3 号和矮秆突变体m1062 的株高及相关性状
2.2 野生型与突变体叶绿体色素含量变化
由图1 表型观察发现,突变体m1062叶色较秀麦3 号深,而叶色一般与色素含量密切相关。由图2可知,突变体的叶绿素a、叶绿素b、叶绿素总量和类胡萝卜素含量均显著高于秀麦3 号,分别比秀麦3 号增加21.9%、12.2%、27.1%、18.8%。叶绿素a/叶绿素b 比值和类胡萝卜素/叶绿素比值变化不大。可见,秀麦3 号矮秆突变体中叶绿素和类胡萝卜素的含量变化与叶色存在一定关系。

图1 野生型秀麦3 号与矮秆突变体m1062 表型
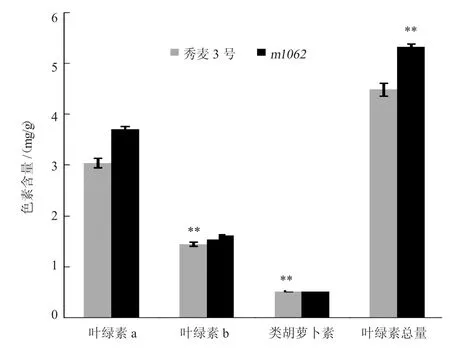
图2 野生型秀麦3 号和突变体m1062 叶片色素含量
2.3 突变体的遗传分析
浙大9 号/m1062组合的F2代株高出现高秆和矮秆分离,随机调查299 个植株,其中72 个植株表现为矮秆、短芒特征。对基因型分析比(227︰72)进行卡方检验表明,符合3︰1 的孟德尔单基因分离比(χ2=0.09<χ20.05=3.84),因此m1062突变体的矮秆性状受1 对隐性单基因控制。
2.4 突变体对大麦产量性状的影响
由表2 可知,与秀麦3 号相比,m1062矮秆突变体的穗粒数和穗数分别增加2.48 粒/穗、1.39 穗/株,增幅分别为8.3%、12.0%,百粒质量降低0.69 g,下降幅度为13.3%,且差异显著(P<0.05);而m1062矮秆突变体的单株质量没有显著变化。m1062/秀麦3 号组合的纯合F3代株中,矮秆株系的每穗粒数、穗数显著大于高秆株系,百粒质量低于高秆株系,但两者单株产量差异不显著。可能是由于单株穗数和粒数的增加弥补了百粒质量,使得单株产量两者基本保持稳定。
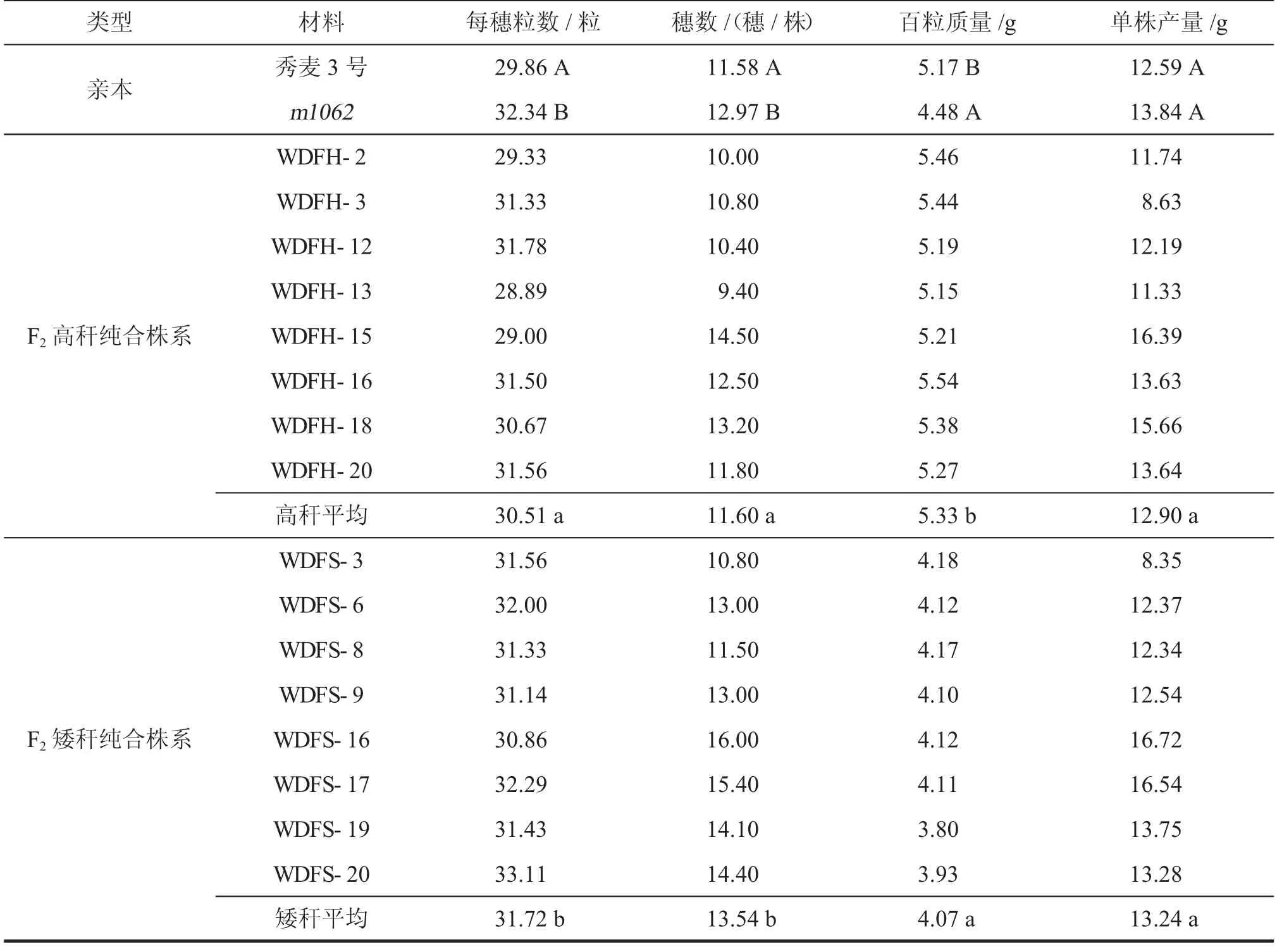
表2 秀麦3 号、矮秆突变体m1062 及其F2 纯合株系产量组成性状
3 讨论与结论
m1062矮秆突变体株高降低27 cm 左右,但旗叶宽和产量没有变化。有研究表明,sdw1/denso使大麦株高降低10~20 cm[5],uzu1使大麦株高降低38.0~46.5 cm[14],而ari-e.GP使大麦株高降低6.2~9.2 cm[11],说明不同的矮秆基因降秆效应差异较大。此外,m1062突变体对穗下茎和第2~4 茎节长度的抑制程度相当,都超过34%,而有研究表明,sdw1/denso对茎节的矮化作用主要体现在第1、2 茎节,对穗下茎相对较小[15]。m1062突变体的穗长、芒长和旗叶长明显变短,这与uzu1和ari-e.GP相似[11,16],说明矮秆突变体除使株高受影响外,其他器官也都存在不同程度的变短。此外,突变体m1062的叶绿素含量和类胡萝卜素含量明显高于秀麦3 号,因此其叶色较秀麦3 号深,这与uzu1突变体相似[16]。
本研究中,m1062突变体抽穗期比秀麦3 号晚5~6 d,可能会减少长江中下游地区“倒春寒”的危害,但这种晚熟特性也可能缩短最佳灌浆时间,从而导致其百粒质量降低。矮秆突变体sdw1/denso为赤霉素敏感型突变体,其抽穗期也推迟,可能是赤霉素合成途径关键基因HvGA20ox2缺失或表达降低,使植株体内赤霉素含量降低[7]。本研究材料m1062在拔节期喷施赤霉素并未能恢复株高,因此该突变体不同于sdw1/denso突变体,表现为赤霉素不敏感。
虽然m1062突变体的百粒质量低于秀麦3 号,但其穗粒数和穗数高于秀麦3 号,最终两者的产量相当。这可能是突变体中叶绿素含量的增加显著提高了光合速率,同时该矮秆突变体m1062分蘖能力增强,表现为穗数增加,因而与产量密切相关的性状穗数和总粒数增加,弥补了百粒质量的降低,使得单株产量基本保持不变。在生产上,矮秆材料可通过增加基本苗数达到增产目的,如sdw1/denso矮秆突变体[15]。另外,由于矮秆突变体m1062受隐性单基因控制,并影响抽穗期和百粒质量,因此在育种中可通过与其他优良品种的杂交或回交,改良该矮秆突变体的抽穗期和百粒质量等农艺性状,进一步达到增产目的。总体来说,m1062矮秆突变体可用于在生产上培育系列矮秆大麦材料。
