小菜蛾基因组中内源性逆转录病毒元件PxERV的发现与特征分析
2020-10-10刘天生陈韶萍尤民生
王 月,卢 丛,刘天生,陈韶萍,尤民生,*
(1.福建农林大学应用生态研究所,闽台作物有害生物生态防控国家重点实验室,福州 350002; 2.福建农林大学,教育部害虫生态防控国际合作联合实验室,福州 350002;3.福建农林大学, 农业部闽台作物有害生物综合治理重点实验室,福州 350002)
内源性逆转录病毒(endogenous retroviruses,ERVs)被认为是生物进化过程中外源逆转录病毒在宿主基因组中的残余序列,能够整合进宿主生殖系统,所以在宿主基因组中世代存留(Löweretal.,1996)。内源性逆转录病毒基因组两端有与外源逆转录病毒类似的长末端重复序列(long terminal repeat sequence,LTRs),中间有1~3个主要的开放阅读框(gag,pol和env)编码病毒复制所需的结构和功能蛋白:gag编码衣壳蛋白;pol编码蛋白酶、逆转录酶、RNA酶H和整合酶;env编码产物用于细胞受体的识别和病毒的侵入(Misserietal.,2004)。通常认为,昆虫内源性逆转录病毒的侵染能力跟env基因的表达相关(Teyssetetal.,1998)。
在多种脊椎动物和非脊椎动物基因组中都发现了内源性逆转录病毒序列,比如鼠、牛、果蝇等(Terzianetal.,2001;Garcia-Etxebarria and Jugo,2010,2016),人类基因组中此类序列约占8%(Belshawetal.,2004),家蚕基因组中也报道了256条内源性逆转录病毒序列(Fengetal.,2018)。由于结构相似,内源性逆转录病毒一度被认为是LTR转座子,后继发现果蝇中gypsy和ZAM元件具有侵染活性(Kimetal.,1994;Leblancetal.,2000),此类序列元件被划定为内源性逆转录病毒。但是国际病毒分类学委员会把脊椎动物内源性逆转录病毒归于逆转录病毒科(Retroviridae),却把昆虫内源性逆转录病毒归于转座病毒科(Metaviridae)(Fablet,2014)。经典的脊椎动物内源性逆转录病毒在宿主正常的生物学过程及疾病发生过程中可能有着重要的作用(Wangetal.,2013;Bustamante Riveraetal.,2017)。在昆虫学研究领域,黑腹果蝇Drosophilamelanogaster(gypsy,17-6,297,ZAM,Idefix,nomad,tirant)、黑果蝇Drosophilavirilis(Tv1)、嗜凤梨果蝇Drosophilaananassae(tom)、粉纹夜蛾Trichoplusiani(Ted)和地中海实蝇Ceratitiscapitata(yoyo)等物种发现了多种内源性逆转录病毒元件(Terzianetal.,2001),甚至gypsy和ZAM的活性周期都有了详细报道(Kimetal.,1994;Leblancetal.,2000),但内源性逆转录病毒对宿主的生物学意义却一无所知。
小菜蛾Plutellaxylostella是世界范围内分布非常广泛的鳞翅目害虫,已经对许多重要的十字花科经济作物造成毁灭性的破坏(Furlongetal.,2013)。在小菜蛾精巢发育的全转录组(数据未发表)研究中,我们发现了一个env基因,本研究以env基因位点为起始点,用生物信息学方法鉴定到一个小菜蛾内源性逆转录病毒元件PxERV,并分析了该内源性逆转录病毒的序列结构,进一步对env基因检测分析,揭示了该内源性逆转录病毒的复制特性,有助于研究内源性逆转录病毒对小菜蛾的影响,为探究昆虫内源性逆转录病毒对宿主的生物学意义奠定基础。
1 材料与方法
1.1 供试昆虫及饲养
供试小菜蛾为实验室人工饲料饲养的G88品系和2009年福州野外采集后在实验室用萝卜苗继代饲养的福州敏感品系(FZ),饲养条件为温度23~25℃,光周期为16L∶8D。
1.2 小菜蛾PxERV的鉴定及序列结构分析
在小菜蛾精巢全转录组(数据未公布)分析中发现了一个基因(编号为MSTRG.10684),经预测发现该基因序列有Baculo_F保守结构域,属杆状病毒融合蛋白(Baculovirus F protein)家族,是一类糖蛋白,可能是内源性逆转录病毒的env基因。在小菜蛾基因组上以PxERV的env基因(编号为MSTRG.10684,克隆序列下载网址http:∥iae.fafu.edu.cn/DBM/family/index.php)位点为起始点,上下游各以10 kb为窗口,用LTR_Finder(Xu and Wang,2007)鉴定LTR结构,确定PxERV的DNA序列;用NCBI工具CD-Search预测序列所包含的保守结构域;用ORFfinder预测env基因的开放阅读框。
1.3 PxERV的env基因克隆
分别取小菜蛾福州敏感品系(FZ)成虫以及G88品系成虫(GA)和幼虫(GL)各10头,单头置于1.5 mL离心管中,液氮冷冻后,加钢珠高速破碎;用DNeasy Bloodand Tissue Kit试剂盒(QIAGEN)按照产品说明书提取小菜蛾基因组DNA;分别用核酸蛋白测定仪和琼脂糖凝胶电泳对DNA浓度及完整性进行检测;样品于-20℃保存。用Primer Premier在小菜蛾PxERV的env基因上下游设计克隆引物S51_F/S51_R(表1),由福州博尚生物技术有限公司合成,以小菜蛾基因组DNA为模板,进行PCR扩增,反应体系(25 μL):2×Hieff Canace Gold PCR Master Mix 12.5 μL,DNA 1 μL,上下游引物(10 μmol/L)各1 μL,ddH2O 9.5 μL。反应程序:98℃ 3 min;98℃ 10 s,55℃ 20 s,72℃ 1 min,32个循环;72℃ 5 min。反应结束后,将未纯化PCR产物送样,委托福州博尚生物技术有限公司进行基因测序。

表1 引物信息Table 1 Primer information
1.4 env基因的系统发育分析
为探究小菜蛾PxERV的env与其他包膜糖蛋白的系统发育关系,我们从NCBI和果蝇数据库(Thurmondetal.,2019)下载了已知的14种昆虫内源性逆转录病毒env基因编码的氨基酸序列和9种昆虫核型多角体病毒(nucleopolyhedrovirus,NPVs)的F或gp64包膜糖蛋白序列,共29条;用MEGA6(Tamuraetal.,2013)的邻接法(Saitou and Nei,1987)对小菜蛾PxERV的env预测氨基酸序列与29条氨基酸序列进行了进化树分析。
1.5 qPCR检测env基因表达
分别在G88品系小菜蛾4龄幼虫、蛹期第0天、蛹期第1天、蛹期第2天和成虫期5个取样时间点进行取样,其中精巢样品各取样时间点每50头虫的精巢为1个重复,重复取样3次;虫体样品各取样时间点每20头剖去精巢的虫体为1个重复,重复取样3次。按照 Eastep Super总RNA提取试剂盒(Promega,北京)操作说明提取总RNA。用1.2%琼脂糖凝胶电泳和核酸蛋白测定仪检测其完整性、纯度和浓度,然后以1 μg总RNA为模板,按照GoScript Reverse Transcription System试剂盒(Promega,北京)合成cDNA第1链,-20℃保存。
以1.3节克隆得到的env基因序列为参考设计qPCR引物10684.q_F/10684.q_R(表1),以合成的cDNA为模板,以核糖体蛋白(PxL32)基因和延伸因子(PxEF1)基因为内参基因,引物序列见表1,每个样品有3个技术重复,用GoTaq qPCR Master Mix试剂盒(Promega,北京)在仪器(CFX96 TouchTMReal-time PCR Detection System,C1000 TouchTMThermal Cycler,Bio-Rad,美国)上进行PCR,测定env基因的表达情况。反应体系(20 μL):2×Go Taq qPCR Master Mix 10 μL,ddH2O 8 μL,cDNA 1 μL,上下游引物(10 μmol/L)各0.5 μL。反应程序:95℃ 2 min;95℃ 15 s,60℃ 1 min,40个循环。
取两个内参基因CT值的几何平均值为内参CT值,用2-ΔΔCT法(Livak and Schmittgen,2001)计算基因相对表达量。
1.6 qPCR检测env基因拷贝数
以1.3节提取的小菜蛾福州敏感品系(FZ)成虫以及G88品系成虫(GA)和幼虫(GL)基因组DNA为检测样本模板,以1.5节的qPCR方法测定env基因在上述样本基因组中的拷贝数。未添加DNA模板的对照的CT值为35,取35与相应样本CT值的差值为该样本env基因拷贝数的量化值,进行不同样本间env基因拷贝数的比较分析。
1.7 CRISPR/Cas9介导env基因突变
以1.3节克隆得到的小菜蛾PxERV的env基因序列为参考,在预测的ORF区设计gRNA引物10684gRNA-F/10684gRNA-R(表1),用PCR合成gRNA的DNA模板,反应体系(50 μL):DNA合成酶25 μL,上下游引物(10 μmol/L)各3 μL,ddH2O 19 μL。反应程序:95℃ 3 min;95℃ 15 s,68℃ 15 s,72℃ 20 s,29个循环;72℃ 2 min。取65 ng上述PCR产物,加2.5 μL NTP Mix和0.5 μL T7 RNA Mix(HiScribeTMT7 Quick High Yield RNA Synthesis Kit,NEB),用ddH2O补至5 μL。37℃,12 h合成gRNA。取400 ng的gRNA与250 ng的Cas9蛋白(GenCrispr Cas9-N-NLS Nuclease,金斯瑞)混匀,37℃孵育20 min;用显微注射仪对260颗G88品系小菜蛾产下30 min内的卵进行注射。G0代成虫与野生型交配,F1代开始一雌一雄自交,每代取单头成虫提DNA(方法同1.3节),用检测引物10684.d-F/10684.d-R(表1)扩增检测片段,送样委托福州博尚生物技术有限公司进行测序。
1.8 数据分析
用IBM SPSS Statistics 22采用单因素方差进行基因相对表达量和拷贝数的差异分析,并进行Tukey HSDa显著性检验。
2 结果
2.1 PxERV的序列结构分析及鉴定
以PxERV的env基因(编号为MSTRG.10684)位点为起始点,上下游各以10 kb为窗口,发现上游6 250 bp处为5′LTR,下游1 824 bp处为3′LTR,长度约268 bp。进一步的结构分析(图1)发现,env基因上游有由开放阅读框pol编码的蛋白酶(retro)、整合酶(rve)、RNA酶H(RNaseH)和逆转录酶(RT_LTR)4种逆转录病毒蛋白结构域。综合分析结果,该PxERV元件全长8 343 bp,有pol和env两个开放阅读框,不包含阅读框gag,没有完整的逆转录病毒结构,符合昆虫内源性逆转录病毒LTR-pol-env-LTR结构(Misserietal.,2004;Fengetal.,2018)。
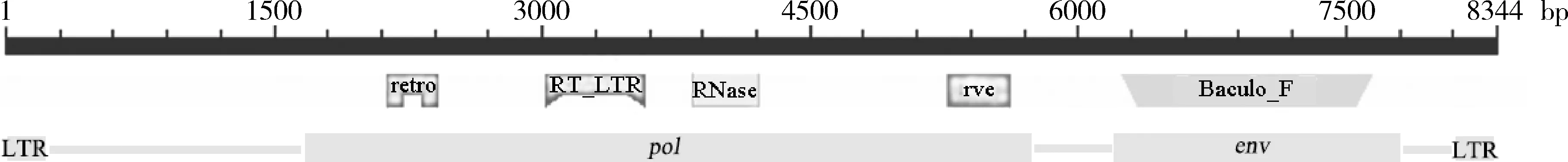
图1 小菜蛾内源性逆转录病毒元件PxERV基因组结构示意图Fig.1 Genome structure of Plutella xylostella endogenous retroviral element PxERVretro:蛋白酶 Protease;RNase:RNA酶H Ribonuclease H;rve:整合酶 Integrase;RT_LTR:逆转录酶Reverse transcriptase;Baculo_F:包膜糖蛋白F Envelope glycoprotein F.
2.2 env基因克隆及分析
克隆得到env基因序列长2 051 bp,预测最长ORF长约1 617 nt,编码538个氨基酸。系统发育分析(图2)发现,昆虫内源性逆转录病毒env基因的蛋白氨基酸序列与昆虫核型多角体病毒(NPV)的包膜糖蛋白氨基酸序列明显分为两支;小菜蛾PxERV的env基因与果蝇Drosophilabuzzatii的逆转座子osvaldo及粉纹夜蛾Trichoplusiani内源性逆转录病毒Ted的env基因进化关系最近。
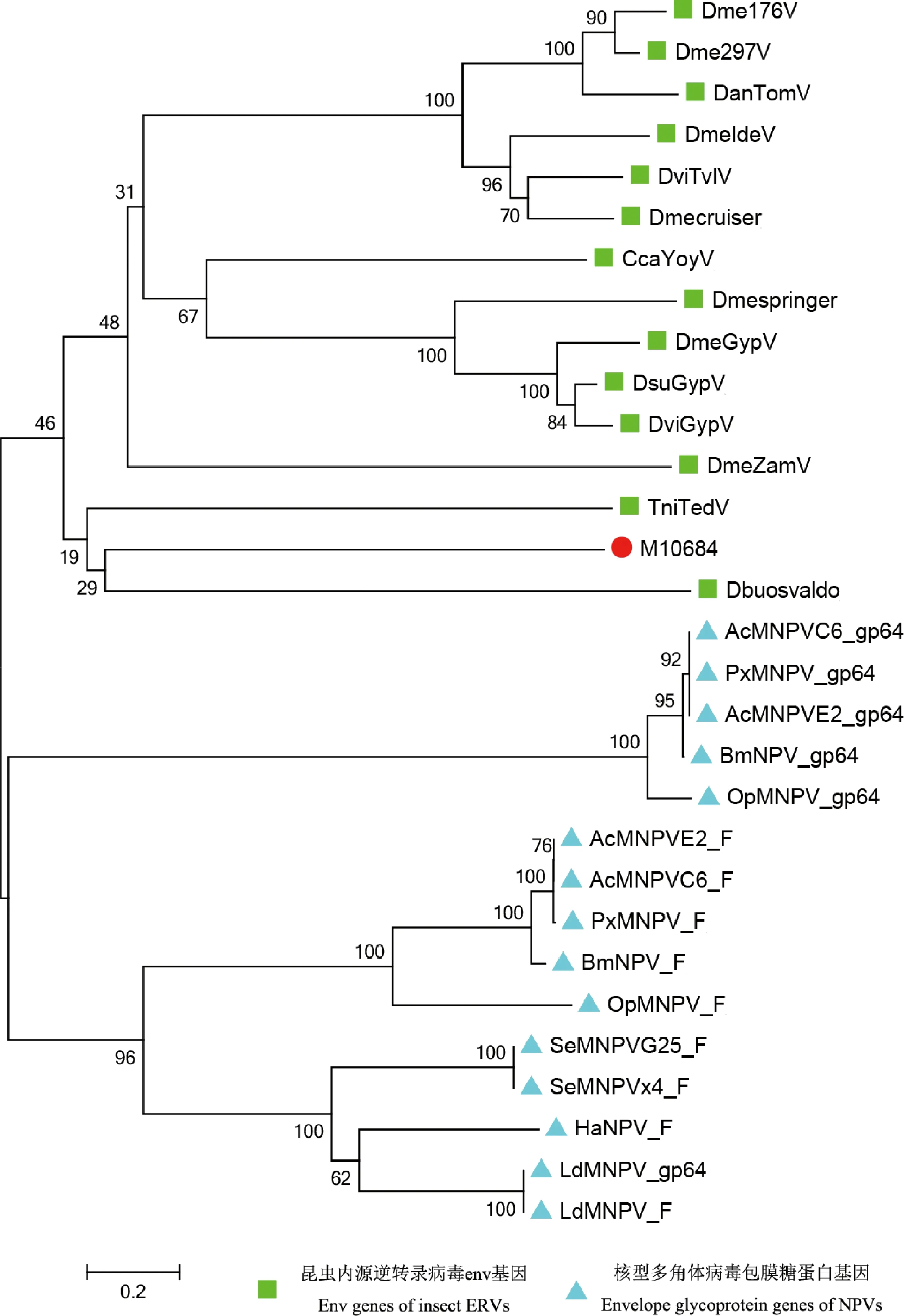
图2 基于氨基酸序列构建的昆虫内源性逆转录病毒env基因编码的氨基酸序列与核型多角体病毒包膜糖蛋白系统发育树(邻接法,1 000次重复)Fig.2 Phylogenetic tree of amino acid sequences coded by env genes of endogenous retroviruses (ERVs) and envelope glycoproteins of nucleopolyhedroviruses (NPVs) of insects constructed based on the amino acid sequence (neighbor-joining method,1 000 replicates)昆虫内源性逆转录病毒env和核型多角体病毒包膜糖蛋白序列来源物种及其GenBank登录号 Source species of env proteins of ERVs and envelope glycoproteins of NPVs of insects and their GenBank accession numbers:DmeGypV:黑腹果蝇Drosophila melanogaster,M12927;Dme176V:黑腹果蝇D.melanogaster,X01472;Dme297V:黑腹果蝇D.melanogaster,X03431;DmeZamV:黑腹果蝇D.melanogaster,AJ000387;DmeIdeV:黑腹果蝇D.melanogaster,AJ009736;DanTomV:嗜凤梨果蝇D.ananassae,Z24451;DsuGypV:果蝇D.subobscura,X72390;DviGypV:黑果蝇D.virilis,M38438;DviTvIV:黑果蝇D.virilis,AF056940;TniTedV:粉纹夜蛾Trichoplusia ni,M32662;CcaYoyV:地中海实蝇Ceratitis capitata,U60529;Dmespringer:黑腹果蝇D.melanogaster,AF364549;Dmecruiser:黑腹果蝇D.melanogaster,AF364550;Dbuosvaldo:果蝇D.buzzatii,AJ133521;AcMNPVC6_gp64:苜蓿银纹夜蛾Autographa californica,L22858;AcMNPVC6_F:苜蓿银纹夜蛾A.californica,L22858;AcMNPVE2_gp64:苜蓿银纹夜蛾E2核型多角体病毒A.californica,KM667940;AcMNPVE2_F:苜蓿银纹夜蛾A.californica,KM667940;BmNPV_gp64:家蚕Bombyx mori,KJ186100;BmNPV_F:家蚕B.mori,KJ186100;OpMNPV_gp64:黄杉合毒蛾Orgyia pseudotsugata,U75930;OpMNPV_F:黄杉合毒蛾O.pseudotsugata,U75930;LdMNPV_gp64:舞毒蛾Lymantria dispar,KX618634;LdMNPV_F:舞毒蛾L.dispar,KX618634;SeMNPVx4_F:甜菜夜蛾Spodoptera exigua,HG425345;SeMNPVG25_F:甜菜夜蛾S.exigua,HG425347;HaNPV_F:棉铃虫Helicoverpa armigera,NC_003094;PxMNPV_gp64:小菜蛾Plutella xylostella,DQ457003;PxMNPV_F:小菜蛾P.xylostella,DQ457003;M10684:小菜蛾P.xylostella,MSTRG.10684.
2.3 env基因的表达
我们对5个发育时期的小菜蛾G88品系虫体和精巢中env基因的表达情况进行了qPCR检测,结果(图3)显示该env基因在成虫精巢中特异性高表达(P<0.05),在其他发育时期的虫体和精巢之间表达量都没有显著差异(P>0.05),但在成虫时期的虫体中比其他时期虫体中表达量稍高;在精巢中的表达量总体比在虫体中表达量稍高。
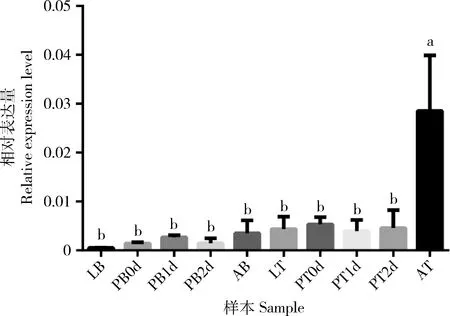
图3 env基因在小菜蛾G88品系不同发育阶段虫体和精巢中的相对表达量Fig.3 Relative expression levels of env gene in the body and testis of Plutella xylostella strain G88 at different developmental stagesLB:4龄幼虫虫体Body of the 4th instar larva;PB0d:蛹期第0天虫体Body of the 0-d-old pupa;PB1d:蛹期第1天虫体Body of the 1-d-old pupa;PB2d:蛹期第2天虫体Body of the 2-d-old pupa;AB:成虫期虫体Adult body;LT:4龄幼虫精巢Testis of the 4th instar larva;PT0d:蛹期第0天精巢Testis of the 0-d-old pupa;PT1d:蛹期第1天精巢Testis of the 1-d-old pupa;PT2d:蛹期第2天精巢Testis of the 2-d-old pupa;AT:成虫期精巢Adult testis.图中数据为平均值±标准误;柱上不同小写字母表示基因表达量在不同发育时期的虫体和精巢间差异显著(P<0.05,Tukey HSDa检验)。Data in the figure are mean±SE.Different lowercase letters above bars indicate significant differences in the gene expression level among insect bodies and testes at different developmental stages (P<0.05,Tukey HSDa test).
2.4 env基因在小菜蛾不同品系中的拷贝数
qPCR检测env基因表达情况时,G88品系成虫期精巢的生物学重复不太稳定,我们猜测可能是由于内源性逆转录病毒元件的拷贝数不同引起的。经检测(图4),env基因的拷贝数在小菜蛾不同品系个体间确实存在差异。另外,在福州敏感品系(FZ)个体中env基因的拷贝数显著少于G88品系个体中的拷贝数(P<0.05),G88品系的幼虫期个体与成虫期个体中env基因拷贝数没有显著差异(P>0.05)。
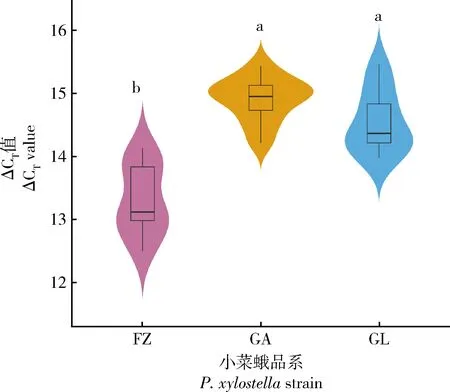
图4 env基因在小菜蛾不同品系和不同龄期的拷贝数Fig.4 Copy number of env gene in different stains and developmental stages of Plutella xylostella未添加DNA模板的对照的CT值为35,取35与相应样本CT值的差值(ΔCT值)为该样本env基因拷贝数的量化值。The CT value of the control without DNA template was 35,and the difference between 35 and the CT value (ΔCT value) was taken to be the quantized value of the env gene copy number in this sample.FZ:福州敏感品系成虫Adult of the susceptible strain from Fuzhou;GA:G88品系成虫Adult of strain G88;GL:G88品系幼虫Larva of strain G88.不同小写字母表示拷贝数在不同样本间差异显著(P<0.05,Tukey HSDa检验)。Different lowercase letters indicate significant differences in the gene copy number among different samples (P<0.05,Tukey HSDa test).
2.5 PxERV的独立复制活性
为探究内源性逆转录病毒的独立复制活性,我们利用CRISPR/Cas9技术共注射小菜蛾260颗卵,成活120头,送测G0代100头,突变率达60%,绝大多数突变虫自交后代的检测序列在突变区的同一位点有3种以上的碱基信号(图5:A),说明虫体中含多种突变类型;在送测小菜蛾中筛得1头只含在312和313位点有两碱基突变(图5:A)的雌虫,其后代连续自交、测序发现,F2代(图5:C)突变型碱基信号的峰高相对F1代(图5:B)变小,自交多代后无法测到突变型碱基信号,说明自交多代,突变型env基因在小菜蛾基因组上不断减少,直至消失,纯合野生型env基因恢复。
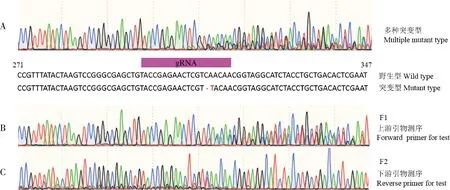
图5 env基因检测片段的碱基信号图(局部)Fig.5 The base signal of partial env gene sequence碱基信号Base signal:C:蓝色Blue;G:黑色Black;T:红色Red;A:绿色Green
3 讨论
本研究中,我们在小菜蛾基因组中发现了一个内源性逆转录病毒元件,长8 343 bp,具有LTR-pol-env-LTR结构(图1)。通常认为昆虫内源性逆转录病毒的侵染能力与env基因的表达相关(Teyssetetal.,1998),所以对内源性逆转录病毒env基因的特征分析,有利于全面了解该内源性逆转录病毒。
缺乏env基因的LTR转座子可以通过整合到杆状病毒的双链DNA基因组中从杆状病毒中“捕获”env基因(Maliketal.,2000)。并且,还有报道发现与杆状病毒GroupⅡ的融合蛋白具有显著同源性的昆虫内源性逆转录病毒的env蛋白有融合活性(Rohrmann and Karplus,2001;Misserietal.,2003,2004)。杆状病毒家族有两类包膜糖蛋白,分别为gp64和F。通常,杆状病毒GroupⅠ含有gp64和F蛋白,而杆状病毒GroupⅡ只含有F蛋白(Pearson and Rohrmann,2002)。通过对昆虫内源性逆转录病毒env蛋白序列和几种昆虫核型多角体病毒的包膜糖蛋白序列比对及系统发育分析发现,昆虫内源性逆转录病毒env基因的蛋白序列与昆虫核型多角体病毒的包膜糖蛋白序列明显分为两支,符合内源性逆转录病毒是古外源逆转录病毒在宿主基因组中的残余这一猜想(Löweretal.,1996)。本研究中的PxERV与粉纹夜蛾内源性逆转录病毒Ted和果蝇逆转座子osvaldo的env基因进化关系最近(图2),我们猜测PxERV可能在早期已整合到昆虫某祖先基因组中,世代留存并不断分化,或者在很早之前,PxERV与Ted和osvaldo分别从某一种寄生在昆虫上的杆状病毒中“捕获”了同一段env基因。
昆虫内源性逆转录病毒ZAM属于gypsy家族,在果蝇的生殖系中可通过RNA中间体实现世代间的转移,其侵染性在果蝇的不同品系中是不一样的,造成拷贝数和插入位点有显著差异(Leblancetal.,1997)。本研究env基因的突变检测发现同一虫体内可包含多种突变类型(图5:A),说明PxERV在小菜蛾基因组上是多拷贝存在的;突变后的env基因多代自交过程中在基因组上不断减少,最终恢复为野生型,说明PxERV在小菜蛾世代间有独立复制活性;env基因拷贝数的量化检测发现PxERV在福州敏感品系(FZ)中的基因拷贝数低于在G88品系中的拷贝数,但G88品系不同发育龄期间的拷贝数没有显著差异(图4),说明PxERV在小菜蛾个体发育过程中不发生独立复制,并且独立复制能力在不同品系间有差异。另有研究发现ZAM的全长RNA与gag和env基因在卵母细胞周围的泡细胞中协同表达,组装成颗粒进入卵母细胞,从而实现ZAM在果蝇中的世代转染(Leblancetal.,2000)。我们研究发现PxERV的env基因在成虫精巢中特异性高表达(图3),我们猜测PxERV可能是在成虫精巢中进行独立复制,通过小菜蛾雄性生殖系统完成世代侵染。这一猜测还需进一步的验证,其世代侵染的分子机制也需进一步的研究。
本研究发现了一个小菜蛾内源性逆转录病毒元件,揭示了它有独立的复制活性,为进一步研究内源性逆转录病毒对小菜蛾的影响奠定了基础,也促进了昆虫内源性逆转录病毒领域的研究,有利于研究内源性逆转录病毒对宿主的生物学意义。
