转飞蝗羧酸酯酶基因果蝇品系的构建及其在有机磷农药代谢解毒中的作用
2020-10-10张徐波张建珍张建琴
尹 菲,杨 洋,张徐波,张建珍,张建琴,*
(1.山西大学中医药现代研究中心,太原 030006;2.山西大学应用生物学研究所,太原030006)
飞蝗Locustamigratoria属于植食性昆虫,主要以禾本科植物为生,危害的农作物涉及玉米、小麦、谷子和高粱,在极端环境下也可危害大豆和白菜等(尤其儆等,1958)。羽化后的成虫在环境温度达到 31~34℃时,开始聚集迁飞,所到之处农作物大面积破坏,造成农业经济损失(尤其儆等,2003;吴瑞芬等,2006)。化学防治法是防治蝗灾的一种有效方法,杀虫剂的长期施用导致飞蝗对马拉硫磷等杀虫剂产生了抗性(Maetal.,2004)。羧酸酯酶是昆虫体内主要的一类代谢解毒酶系,其功能主要是参与包含酯键的杀虫剂等外源性化合物的解毒,信息素的分解和神经发育及调控(Heikinheimoetal.,1999)。一般情况下当羧酸酯酶与有机磷杀虫剂相互反应时,去磷酸化反应的速率较慢,昆虫体内羧酸酯酶基因无法与底物快速分离,此时酶活性被抑制。但是在一些具有有机磷农药抗性的昆虫体内,存在着一种特殊的酶类,这种酶可以快速将马拉硫磷分解为无毒的产物,从而使得昆虫对马拉硫磷的耐受性增强,进而使得昆虫对该农药具有抗性(Heidarietal.,2004;Cuietal.,2007)。Yang等(2008)的研究表明野生型飞蝗马拉硫磷抗性品系的羧酸酯酶活性提高。由此得知,羧酸酯酶在飞蝗对有机磷的抗性中发挥重要作用。
本课题组前期采用RNA干扰(RNA interference,RNAi)结合杀虫剂生物学测定实验结果显示处于A簇的直翅目昆虫α酯酶基因LmCesA1和LmCesA2与有机磷农药的代谢解毒相关(Zhangetal.,2013),但这两个基因在飞蝗体内的超表达是否会导致飞蝗对马拉硫磷的抗性还不清楚。目前已有研究利用果蝇模型超表达非模式昆虫的基因,借助转基因果蝇技术优势研究非模式生物基因的功能(Riveronetal.,2013,2014)。因此,本研究利用果蝇模型,分析飞蝗羧酸酯酶基因LmCesA1和LmCesA2的超表达及其与对马拉硫磷的抗性的关系,以期为飞蝗抗性的监测和治理提供理论依据。
1 材料与方法
1.1 供试昆虫
果蝇品系:RB0006亲本黑腹果蝇Drosophilamelanogaster品系,基因型为{y v;attP40,y+},购买于北京清华大学医学院果蝇实验中心。tub-Gal4品系可启动靶标基因的全身表达,该品系由中国科学院遗传与发育生物学研究所姚爱玉老师友情提供;act-Gal4品系可启动靶标基因的全身表达;c601-Gal4品系是肠道组织特异表达的果蝇品系,act-Gal4与c601-Gal4由山西大学应用生物学研究所友情提供。
果蝇饲养条件:温度25±2℃,相对湿度60%~70%,光周期14L∶10D。
1.2 供试试剂
DEPC水购自Sangon公司;Trizol,DNA Marker,6×Loading Buffer,Olige(dT)12-18 Primer,5×M-MLV Buffer,dNTP Mixture,RNase Inhibitor,Ex Taq和RTase M-MLV均购自TaKaRa公司;2×Taq PCR Master购自Tiangen公司;SYBR Green Realtime PCR Master Mix购自Toyobo公司;马拉硫磷购自Sigma公司;其他化学试剂均为国产试剂。
1.3 转基因果蝇的构建
本研究转飞蝗羧酸酯酶基因果蝇构建委托清华大学医学院果蝇模式中心完成,具体的研究方法如下:根据飞蝗羧酸酯酶基因LmCesA1和LmCesA2的核苷酸序列,采用在线软件NEBcutter V2.0 (http:∥tools.neb.com/NEBcutter2/)分析这两个基因的酶切位点,结合载体pVALIUM20多克隆位点内切酶类型,设计构建载体引物(表1)。后经PCR扩增,克隆测序确定将目的基因克隆到pVALIUM20载体中。将一定浓度的重组载体注射到亲本果蝇受精卵中,筛选到的转基因果蝇再与balancer果蝇杂交,得到纯合的、稳定的转基因果蝇品系UAS-LmCesA1和UAS-LmCesA2,该转基因技术采用了定点转座技术,目的基因以单拷贝的方式插入到果蝇2号染色体上。
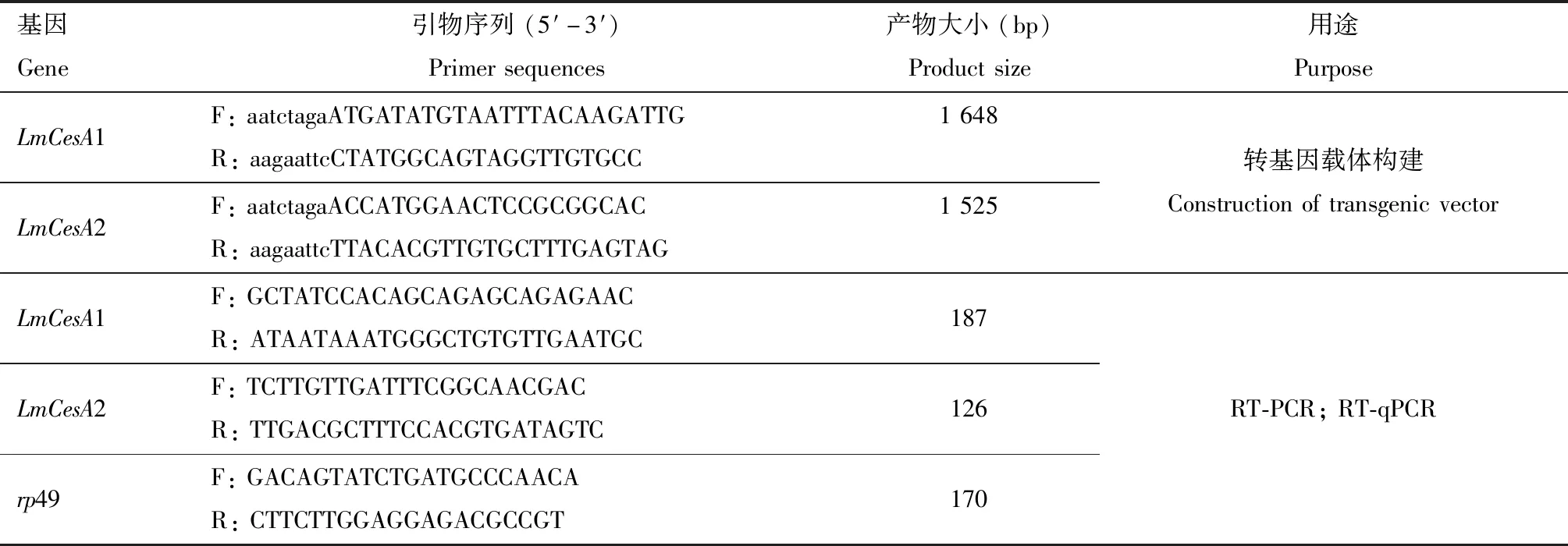
表1 用于本研究的引物Table 1 Primers used in this study
1.4 转基因果蝇与3品系Gal4果蝇杂交试验
分别挑取不同Gal4果蝇中的处女蝇,与转基因果蝇品系UAS-LmCesA1和UAS-LmCesA2的父本果蝇以雌雄2∶1的比例进行杂交,按照标准挑取所需要的子一代果蝇,RB0006品系作为对照果蝇,具体步骤如下:
(1)act-Gal4品系的处女蝇与UAS-LmCesA1品系的雄蝇杂交,子一代基因型为act>LmCesA1;act-Gal4品系的处女蝇与UAS-LmCesA2品系的雄蝇杂交,子一代基因型为act>LmCesA2;act-Gal4品系的处女蝇与RB0006品系的雄蝇杂交,子一代基因型为act>attP40。act-Gal4果蝇带有卷翅平衡子,因此,挑取子一代的直翅果蝇进行后续研究。
(2)tub-Gal4品系的处女蝇与UAS-LmCesA1品系的雄蝇杂交,子一代基因型为tub>LmCesA1;tub-Gal4品系的处女蝇与UAS-LmCesA2品系的雄蝇杂交,子一代基因型为tub>LmCesA2;tub-Gal4品系的处女蝇与RB0006品系的雄蝇杂交,子一代基因型为tub>attP40。tub-Gal4果蝇带有TM6B平衡子,因此,挑取子一代带有两根簇刚毛的果蝇进行后续研究。
(3)c601-Gal4品系的处女蝇与UAS-LmCesA1品系的雄蝇杂交,子一代基因型为c601>LmCesA1;c601-Gal4品系的处女蝇与UAS-LmCesA2品系的雄蝇杂交,子一代基因型为c601>LmCesA2;c601-Gal4品系的处女蝇与RB0006品系的雄蝇杂交,子一代基因型为c601>attP40,c601-Gal4品系是肠道组织特异表达的果蝇品系,子一代全部为靶标基因超表达果蝇。
1.5 转基因果蝇的分子鉴定
1.5.1转基因果蝇品系DNA水平的鉴定:转基因果蝇品系与tub-Gal4果蝇品系杂交,分别选择4-6 d杂交子一代tub>LmCesA1,tub>LmCesA2和tub>attP40的雌性果蝇进行DNA提取,采用转基因载体构建引物(表1)进行PCR扩增。25 μL PCR反应体系:DNA(10×) 2 μL,TaKaRa Ex Taq (5 U/μL) 0.1 μL,10×Ex Taq Buffer 2.5 μL,dNTPs(2.5 mmol/L) 2 μL,上下游引物各 0.5 μL,ddH2O 17.4 μL。反应条件:95℃ 1 min;95℃ 30 s,56℃ 30 s,72℃ 2 min,35个循环;72℃ 10 min,4℃保存,用2%的琼脂糖凝胶电泳检测PCR产物,采用天根公司的胶回收试剂盒进行PCR产物回收,T-A克隆测序。
1.5.2转基因果蝇品系RNA水平的鉴定:分别选择各组羽化后第4-6天的子一代雌蝇提取总RNA,体外反转录合成cDNA第1链,以cDNA为模板,以表1设计的引物进行RT-PCR反应,rp49作为内参基因。RT-PCR 反应体系:cDNA(10×)4 μL,上下游引物各0.8 μL,2×Taq Master Mix 12.5 μL,ddH2O 6.9 μL。反应条件:94℃ 3 min;94℃ 30 s,60℃ 30 s,72℃ 1 min (LmCesA1和LmCesA2基因28个循环,内参rp49基因25个循环);72℃ 5 min。RT-PCR实验分组按照1.4节杂交分组,每个组别共设置3个生物学重复,5头果蝇为一个生物学重复。
采用RT-qPCR法分别检测靶标基因在c601-Gal4果蝇品系与UAS-LmCesA1品系和UAS-LmCesA2品系杂交子一代成虫不同组织脑、肠和表皮等中的mRNA表达情况,rp49作为内参基因。引物如表1。RT-qPCR反应体系:SYBR Green Real-time PCR Master Mix 10 μL,上下游引物各 0.8 μL,cDNA (10×) 4.0 μL,灭菌水4.4 μL。反应条件:95℃ 1 min;95℃ 5 s,55℃ 10 s,72℃ 15 s,40个循环;95℃ 1 min,55℃ 1 min,97℃ 1 min,37℃ 30 s。至少50头果蝇为一个生物学重复,共设置4个生物学重复。
1.6 转基因果蝇对马拉硫磷的耐受性测定
生物学测定采用的果蝇为c601>LmCesA1,c601>LmCesA2和c601>attP40。用纯丙酮将马拉硫磷配制成一系列浓度梯度。挑取羽化后4-6 d的子一代果蝇,并用CO2麻痹,将0.25 μL的农药点滴于果蝇的腹部,每组选用15头雌性果蝇进行处理,每个组设置3个生物学重复。将果蝇置于培养箱中,温度设置为25±1℃,相对湿度为60%,24 h后记录果蝇的死亡率,当果蝇轻触不动则视为死亡。
1.7 数据分析
RT-qPCR数据利用ABI Prism 7500 SDS 1.1软件进行分析,基因的相对表达量采用2-ΔΔCt法,使用SPSS 19.0软件进行统计分析,采用Duncan氏多样本间差异比较目标基因在各品系中的表达量的差异。生物测定数据用SPSS软件的Probit方法计算LC50。
2 结果
2.1 转基因果蝇品系DNA水平的鉴定结果
以DNA为模板进行PCR扩增,LmCesA1和LmCesA2的PCR产物的长度分别为1 648 bp和1 525 bp,由图1可知,转基因果蝇tub>LmCesA1和tub>LmCesA2中均能扩增到目的基因,而对照组果蝇tub>attP40中未扩增到目的基因。测序结果显示插入的目的基因序列正确无误。

图1 转基因黑腹果蝇DNA水平的鉴定结果Fig.1 Identification results of transgenic Drosophila melanogaster at the DNA levelM:DL5000 DNA Marker;tub>attP40:对照组果蝇Control Drosophila;tub>LmCesA1:过表达LmCesA1的转基因果蝇Transgenic Drosophila overexpressing LmCesA1;tub>LmCesA2:过表达LmCesA2的转基因果蝇Transgenic Drosophila overexpressing LmCesA2.
2.2 转基因果蝇品系RNA水平的鉴定结果
将转基因果蝇UAS-LmCesA1,UAS-LmCesA2和亲本果蝇RB0006分别与3品系Gal4果蝇杂交。选择其第4-6天的子一代提取总RNA,并体外反转录获得cDNA用于RNA水平的基因鉴定,结果显示:靶标基因LmCesA1和LmCesA2在对照组(act>attP40;tub>attP40;c601>attP40)中均没有表达;LmCesA1在不同Gal4品系与UAS-LmCesA1果蝇的杂交子一代act>LmCesA1(图2:A)、tub>LmCesA1(图2:B)和c601>LmCesA1(图2:C)中均有表达;LmCesA2在不同Gal4品系与UAS-LmCesA2果蝇的杂交子一代act>LmCesA2(图2:A)、tub>LmCesA2(图2:B)和c601>LmCesA2(图2:C)中均有表达。
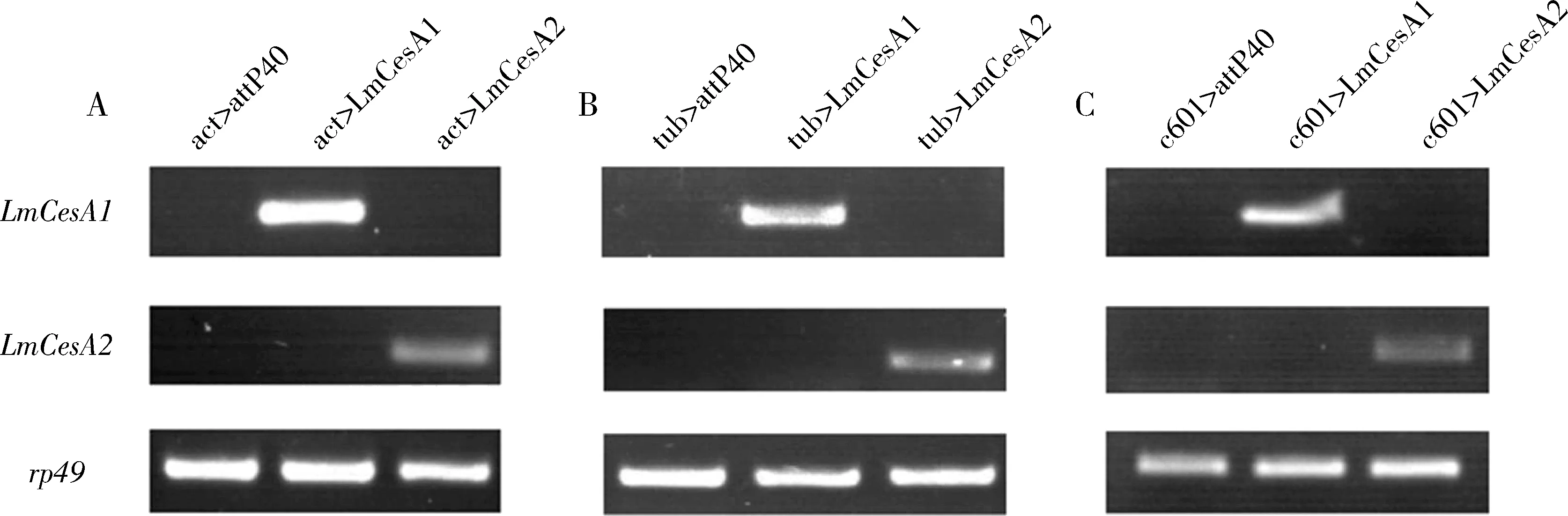
图2 转基因黑腹果蝇RNA水平的鉴定结果Fig.2 Identification results of transgenic Drosophila melanogaster at the RNA levelact>attP40:act-Gal4品系与对照组果蝇RB0006品系的杂交子一代Hybrid offspring of act-Gal4 strain and the control RB0006 strain;act>LmCesA1:act-Gal4品系与UAS-LmCesA1品系的杂交子一代Hybrid offspring of act-Gal4 strain and UAS-LmCesA1 strain;act>LmCesA2:act-Gal4品系与UAS-LmCesA2品系的杂交子一代Hybrid offspring of act-Gal4 strain and UAS-LmCesA2 strain.tub>attP40:tub-Gal4品系与对照组果蝇RB0006品系的杂交子一代Hybrid offspring of tub-Gal4 strain and the control RB0006 strain;tub>LmCesA1:tub-Gal4品系与UAS-LmCesA1品系的杂交子一代Hybrid offspring of tub-Gal4 strain and UAS-LmCesA1 strain;tub>LmCesA2:tub-Gal4品系与UAS-LmCesA2品系的杂交子一代Hybrid offspring of tub-Gal4 strain and UAS-LmCesA2 strain;c601>attP40:c601-Gal4品系与对照组果蝇RB0006品系的杂交子一代Hybrid offspring of c601-Gal4 strain and the control RB0006 strain;c601>LmCesA1:c601-Gal4品系与UAS-LmCesA1品系的杂交子一代Hybrid offspring of c601-Gal4 strain and UAS-LmCesA1 strain;c601>LmCesA2:c601-Gal4品系与UAS-LmCesA2品系的杂交子一代Hybrid offspring of c601-Gal4 strain and UAS-LmCesA2 strain.
c601-Gal4在果蝇中肠上皮细胞中高表达,本研究采用RT-qPCR方法鉴定了目的基因LmCesA1和LmCesA2分别在转基因果蝇c601>LmCesA1和c601>LmCesA2成虫不同组织部位(脑、表皮和肠道)中的表达情况。由图3可知,两个目的基因LmCesA1和LmCesA2均在转基因果蝇的肠道中高表达。LmCesA1在c601>LmCesA1果蝇肠道中的表达量分别是脑和表皮中的7.6和16.7倍。LmCesA2在c601>LmCesA2果蝇肠道中的表达量分别是脑和表皮中的5.4和10.9倍。
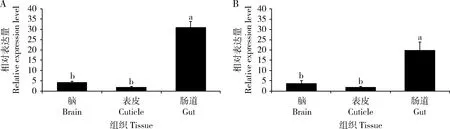
图3 LmCesA1(A)与LmCesA2(B)基因在转基因黑腹果蝇成虫不同组织中的表达谱Fig.3 Expression profiles of LmCesA1 (A) and LmCesA2 (B) in different adult tissues of transgenic strains of Drosophila melanogaster选择脑、表皮和肠道3个组织部位进行分析;每组织设4个生物学重复,实验采用3个技术重复;rp49 为内参基因。数据为平均值±标准误;柱上不同字母代表不同组织之间基因表达量有显著性差异(P<0.05,Duncan氏多重比较检验)。The analyses were conducted in three different tissues,including the brain,cuticle and gut,and rp49 was used as a reference gene.Data are mean±standard errors (SE) of four biological replications (n=4),each with three technical replications.Different letters above bars indicate significant differences in the gene expression level among tissues by Duncan’s multiple comparison test (P<0.05).
2.3 转基因果蝇对马拉硫磷的耐受性
转基因果蝇对马拉硫磷的耐受性研究结果显示:与对照组果蝇(c601>attP40)相比,超表达LmCesA1的转基因果蝇(c601>LmCesA1)和超表达LmCesA2的转基因果蝇(c601>LmCesA2)抗药性均提高,抗性倍数分别为1.06和1.67(表2)。
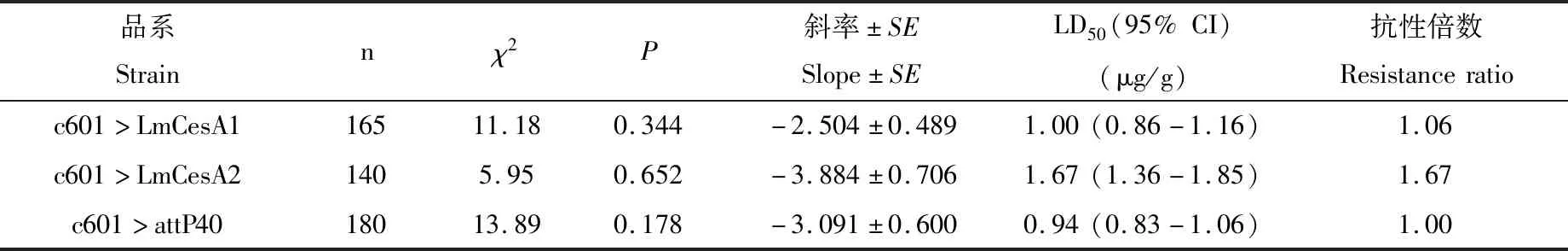
表2 分别超表达LmCesA1和LmCesA2的黑腹果蝇品系对马拉硫磷的耐受性Table 2 Tolerance of Drosophila melanogaster strains overexpressing LmCesA1 or LmCesA2 to malathion
3 讨论
本研究通过转基因果蝇技术阐明飞蝗羧酸酯酶基因(LmCesA1和LmCesA2)与有机磷农药代谢解毒的关系。当前,研究者可采用多种方法鉴定与农药解毒或抗性相关的昆虫羧酸酯酶。例如,Wei等(2016)通过RT-qPCR技术对朱砂叶螨Tetranychuscinnabarinus羧酸酯酶基因(CarE)进行分析,结果显示,相比于敏感品系,有8个羧酸酯酶基因(CarE3,CarE6,CarE7,CarE9,CarE10,CarE13,CarE15和CarE18)参与了甲氰菊酯的解毒过程,有4个羧酸酯酶基因(CarE4,CarE5,CarE14和CarE19)参与了氟甲氰菊酯的解毒过程;Wang等(2017)采用RNAi以及异源表达和细胞毒性实验,证明了桔小实蝇Bactroceradorsalis的羧酸酯酶基因BdB1参与了对马拉硫磷的解毒过程;Feng等(2018)基于家蝇Muscadomestica转录组和基因组数据库对比黑腹果蝇D.melanogaster和冈比亚按蚊Anophelesgambiae,构建羧酸酯酶的系统发育树,最终在家蝇中共鉴定出39个不同功能分支的羧酸酯酶基因。上述研究技术可初步筛选到与抗药性相关的基因,但无法进一步深入探究和验证哪些候选基因具体参与了哪种杀虫剂的代谢。果蝇因其独特的生理特征,如易于室内培养、染色体数目少、表型易于观察以及发育繁殖快且繁殖量大等,被广泛应用于遗传学、发育生物学、医学病理学等研究。果蝇用于经典遗传学领域以揭示真核生物染色体结构,染色体遗传连锁等(Nüsslein-Volhard and Wieschaus,1980)。近年来,许多学者尝试将其他昆虫外源基因转到果蝇基因组上,以研究这些基因的功能(常海静,2011;朱昱璇,2016;Stinchfieletal.,2019;张徐波等,2019)。Zhu等(2010)首次在国际上报道了将转基因果蝇成功应用于赤拟谷盗Triboliumcastaneum抗性基因 P450与溴氰菊酯代谢解毒关系的研究中。黑腹果蝇因其体内具备完善的Gal4/UAS体系,可以实现外源基因在其体内的超表达,进而对靶标基因进行生物测定、生物化学与分子生物学研究,可更深入地阐明该基因的功能。基因缺失性功能研究和基因获得性功能研究是研究基因功能的两种重要的方法。但就目前的实验技术,在飞蝗体内难以实现基因的获得性功能研究。因此,本研究采用Gal4/UAS系统,实现了飞蝗羧酸酯酶基因(LmCesA1和LmCesA2)在黑腹果蝇体内的超表达,以期在果蝇体内完成基因获得性功能研究。
转基因黑腹果蝇RNA水平的鉴定结果显示,3品系Gal4(act-Gal4,tub-Gal4和c601-Gal4)果蝇与转基因果蝇杂交的子一代均可检测出目的基因的表达(图2),表明3品系Gal4果蝇均可启动两个靶标基因的表达,且针对单一基因的mRNA的表达量无显著差异。c601-Gal4在果蝇中肠上皮细胞中高表达,为验证c601-Gal4果蝇与转基因果蝇杂交后代中靶标基因表达部位,本研究采用RT-qPCR方法,进一步验证了靶标基因在转基因果蝇成虫脑、表皮和肠道等不同组织部位的表达情况。结果显示,LmCesA1和LmCesA2在果蝇肠道中高表达,且在果蝇肠道中的表达量分别是表皮中表达量的16.7和10.9倍(图3)。因本课题前期研究显示这两个羧酸酯酶基因(LmCesA1和LmCesA2)主要在飞蝗的中肠和胃盲嚢表达。为尽可能模拟目的基因在飞蝗体内环境,在后续研究中我们采用c601-Gal4果蝇与转基因果蝇杂交后代。
本课题组前期研究结果显示,与对照组飞蝗相比,LmCesA1基因沉默的飞蝗对马拉硫磷的敏感性提高30.9%。而在本研究中,与对照组果蝇(c601>attP40)相比,转入飞蝗羧酸酯酶基因LmCesA1果蝇对马拉硫磷的抗性没有显著变化(表2)。这个结果与前期研究结果存在差异,可能有两个原因:(1)LmCesA1在转基因果蝇中的蛋白水平较低;(2)LmCesA1对马拉硫磷的解毒作用较弱。与对照组果蝇(c601>attP40)相比,转入飞羧酸酯酶基因LmCesA2 果蝇对马拉硫磷的抗性显著升高。这可能是插入了外源羧酸酯酶基因增加了果蝇体内目的基因拷贝数,使得羧酸酯酶活性升高,而产生了抗药性。本课题组通过RNAi结合杀虫剂生物测定实验表明LmCesA2 可能参与飞蝗对马拉硫磷的代谢解毒过程(Zhangetal.,2011)。本研究转基因果蝇对马拉硫磷代谢解毒的研究结论与通过RNAi方法研究LmCesA2 与对马拉硫磷的解毒关系的结论一致。据此我们推测飞蝗LmCesA2极有可能参与飞蝗对马拉硫磷的代谢解毒。
本研究通过转飞蝗羧酸酯酶基因果蝇生物学测定实验结果表明飞蝗LmCesA2极有可能参与飞蝗对马拉硫磷的代谢解毒。但该实验结果仅具有参考意义,并不能完全认定LmCesA1和LmCesA2基因参与飞蝗生物体内杀虫剂代谢解毒过程,其原因有:(1)基因在体内的表达水平和空间分布在黑腹果蝇和飞蝗之间存在差异;(2)在果蝇中,对照品系和抗性品系之间只有一个遗传差异,而在飞蝗体内可能有其他抗性基因影响靶标基因的表达(Dabornetal.,2012)。因此,为进一步确定该基因与农药代谢关系,我们可以在今后的研究中建立飞蝗超表达体系或采用靶标基因的体外表达系统从多维角度全面解析基因与杀虫剂代谢解毒的关系。
