非生物胁迫因子对棘孢木霉嗜铁素及钙调磷酸酶基因表达的影响
2020-09-05王越,赵蕾
王 越, 赵 蕾
(山东师范大学 生命科学学院,山东 济南 250014)
木霉菌(Trichodermaspp.)是一类种类繁多、功能多样的丝状真菌,是当前绿色农业生产中最具应用潜力的生防微生物之一,能增强植物抗生物和非生物胁迫的能力[1-2]。随着对木霉菌生防机制的深入研究,木霉菌产生的次级代谢物在生防、促生及诱导植物抗病中的作用日益受到重视,迄今为止,已经明确结构的木霉菌次级代谢产物有100多种,其中对嗜铁素(siderophore)的研究近年来受到关注。嗜铁素又称铁载体,是微生物在缺铁环境中分泌的一种能够特异性螯合兼转运三价铁离子到微生物或植物体细胞内的小分子水溶性化合物。真菌嗜铁素属于异羟肟酸型,均含有N5-acyl-N5-hydroxyornithine结构单元[3],对真菌自身的生长发育有重要作用[1,4]。进化树分析表明,木霉属中SidA基因 和NPS6 基因参与嗜铁素的合成,并起关键作用[3]。尽管目前对某些真菌(如曲霉菌)嗜铁素的结构、类型有了较为清楚的了解,但逆境对木霉菌产生嗜铁素的影响及调控机制尚不明确,有待于在理论上进一步完善与补充。本实验室已经分离鉴定的高产嗜铁素棘孢木霉(Trichodermaasperellum)菌株T6是本实验的重要材料。钙调磷酸酶(calcineurin)是迄今发现的唯一受Ca2+/钙调素调节的丝/苏氨酸蛋白磷酸酶,由一个催化亚基(CnA) 和一个调节亚基(CnB) 组成,在真核生物中广泛存在[5-8]。其酶活功能主要由A亚基体现,B亚基主要起调节作用[9]。钙调磷酸酶与多种真菌抗逆性有关,在隐球酵母(Cryptococcusneoformans)、白色念珠菌(Candidatropicalis)、粗糙脉孢菌(Neurosporacrassa)等真菌中,具有调控菌体生长、细胞壁完整性、致病性[10-12]以及对外源离子胁迫反应[13]等重要作用。目前,随着生物学技术的发展,丝状真菌中钙调磷酸酶的功能也越来越多的受到人们的关注,但钙调磷酸酶信号通路对木霉次生代谢产物的调控和机制并不明确,对钙调磷酸酶信号通路与嗜铁素合成关系的研究也未见报道。缺铁胁迫能诱导木霉产大量的嗜铁素,本实验室前期发现木霉菌及其嗜铁素能够促进植物的生长[14],但其他胁迫因素对木霉嗜铁素合成的影响尚不明确,与菌体抗胁迫能力相关的钙调磷酸酶基因与嗜铁素基因之间的调控关系也不清楚,因此,本研究选择常见的土壤环境胁迫因子NaCl、pH以及参与调节钙调磷酸酶途径的Ca2+作为影响因子,分别研究其对棘孢木霉菌T6的嗜铁素产量以及对嗜铁素合成关键基因(SidA)与钙调磷酸酶催化亚基基因(CnA)表达的影响,从而初步明确木霉抗逆和嗜铁素合成的分子调控机制,对进一步扩大木霉菌在农业生产中的应用具有重要意义。
1 材料与方法
1.1 材料
1.1.1 实验菌株 棘孢木霉菌(Trichodermaasp-erellum)T6(由本实验室分离鉴定保藏)。
1.1.2 培养基 PDA培养基,LNM(Liquid Nutrient Medium)培养基。
1.1.3 试剂 RNA提取试剂(大连TaKaRa生物科技有限公司);反转录试剂盒和PCR试剂盒(南京诺唯赞生物科技有限公司);引物合成(上海生工生物工程有限公司)。
1.1.4 仪器与设备 生化培养箱(SPX-250B-Z型,上海博迅实业有限公司医疗设备厂);振荡培养箱(SPX-250B-D型,上海博迅实业有限公司医疗设备厂);立式压力蒸汽灭菌器(上海博迅实业有限公司医疗设备厂);冷冻离心机(5410R型,德国 Eppendorf 公司);数显恒温水浴锅(HH-6型,国华电器有限公司);T6新世纪-紫外可见分光光度计(北京普析通用仪器有限责任公司);PCR仪(6331型,德国 Eppendorf 公司)。
1.2 方法
1.2.1 孢子悬液的制备 将棘孢木霉菌T6接种到PDA平板上,28 ℃培养至木霉菌丝体表面被绿色孢子覆盖,向平板中倒入适量无菌水,无菌枪头多次吹打混匀。吸取适量体积的高浓度孢子液加入含玻璃珠的锥形瓶中,摇匀打散孢子,置于显微镜下血球计数板计数,将孢子浓度调整为1×107cfu/mL,备用。
1.2.2 嗜铁素产量的测定[15]取1 mL浓度为107cfu/mL孢子悬液,接种至LNM液体培养基,28 ℃、180 r/min振荡培养5 d后,11 000 r/min,离心15 min,取上清。按体积比1∶1加入CAS检测液,充分混匀后于黑暗中室温静置30 min,630 nm处测吸光值(As),以双蒸水为对照调零。另取空白培养基与CAS检测液等体积混匀,以其吸光值作参比值(Ar),根据公式((Ar-As)/Ar) × 100 %,计算出嗜铁素的相对含量。按上述方法分别测定在不同的NaCl浓度、CaCl2浓度及pH 值条件下的嗜铁素产量,作图比较。其中NaCl浓度分别为0、100、300、500、800、1 000 mmol/L,CaCl2浓度分别为0、0.5、1、3、5、10 mmol/L,pH值分别为2.9、4.9、6.9、8.9、10.9。所有处理均设3组重复。
1.2.3 环孢菌素A(Cyclosporin A, CsA)对棘孢木霉菌T6嗜铁素产量的影响 向LNM培养基中加入钙调磷酸酶专性抑制剂CsA,使其终浓度为30 μmol/L,另设空白对照,分别接种1 mL浓度为107cfu/mL棘孢木霉菌T6孢子悬液,28 ℃、180 r/min摇瓶培养5 d、11 000 r/min、离心15 min,取上清液,测定其嗜铁素含量及菌体的生物量,计算单位菌体嗜铁素含量。
1.2.4 胁迫对棘孢木霉菌T6嗜铁素基因与钙调磷酸酶基因表达的影响 取1 mL浓度为107cfu/mL的孢子悬液分别加到不同处理的LNM液体培养基中,28 ℃、180 r/min振荡培养5 d后,收集菌丝,提取各样品总RNA,并反转录合成第一链cDNA作为PCR模板。50 μL半定量PCR扩增体系含: 2×TaqMaster Mix(诺唯赞,南京),正反向引物(10 mmol/L)各1 μL,模板cDNA 100 ng。每个样品3次重复。扩增程序:95 ℃预变性5 min;95 ℃ 30 s,56 ℃ 30 s,72 ℃ 60 s,30个循环;72 ℃,7 min。将PCR产物进行琼脂糖凝胶电泳,观察并拍照。

表1 引物序列
2 结果与分析
2.1 胁迫因子对棘孢木霉菌T6嗜铁素产量的影响
振荡培养5 d后,测量嗜铁素相对含量。与对照组(0 mol/L NaCl)相比,NaCl浓度为100、300、 500、800以及1 000 mmol/L时,其嗜铁素产量分别下降了2.174%、8.597%、31.64%、81.92%以及100%(图1A),NaCl浓度为1 mol/L时,完全抑制了菌体T6的生长。由图1B可知,CaCl2浓度为0.5 mmol/L时,嗜铁素含量略有增加,而随着CaCl2浓度的增加,嗜铁素相对含量降低,CaCl2浓度为10 mmol/L时,嗜铁素的合成被完全抑制。另外,pH 4.9时,T6的胞外嗜铁素产量最高,pH 6.9次之,pH 8.9最低,即pH的增加会抑制嗜铁素的产生(图1C)。
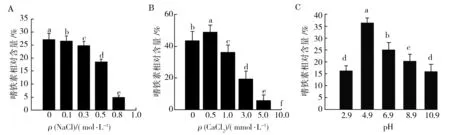
图1 不同处理对棘孢木霉T6嗜铁素产量的影响Fig.1 Effect of different treatment on siderophore production of T. asperellum T6图中a、b、c、d、e、f代表显著性差异a,b,c,d,e,f represents a significant difference in the figures
2.2 CsA对棘孢木霉菌T6嗜铁素产量的影响
按1.2.2方法操作,取上清液,测定其嗜铁素含量及菌体的生物量。结果见图2,加入CsA后单位菌体嗜铁素相对含量增加,说明钙调磷酸酶活性被抑制反而促进了嗜铁素的产生,推测钙调磷酸酶信号通路负调控木霉嗜铁素的产生。
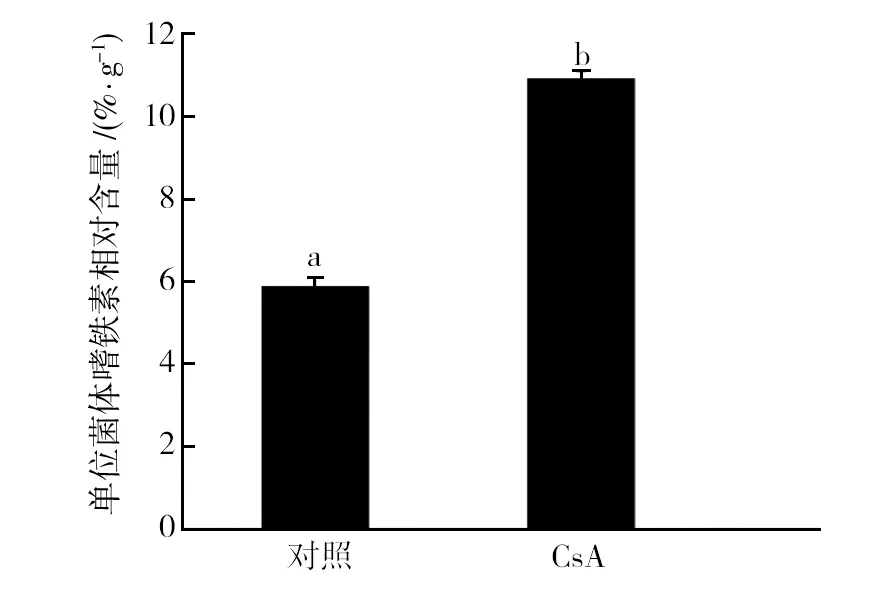
图2 CsA对棘孢木霉T6嗜铁素产量的影响Fig.2 Effect of CsA on the siderophore production of T. asperellum T6
2.3 胁迫因子对棘孢木霉菌T6 SidA和CnA基因表达量的影响
由图3可知,每种处理由上向下依次为钙调磷酸酶催化亚基基因(CnA)、嗜铁素合成关键基因(SidA)以及内参基因(Act)。NaCl浓度的增加可以促进CnA基因的表达,而SidA基因的表达下调,且显著低于CnA基因表达量。CaCl2处理下,CnA基因表达量随着CaCl2浓度的升高而增加,当CaCl2浓度为10 mmol/L时,其表达量最高;SidA基因的表达量先增加后降低,且当CaCl2浓度为10 mmol/L时,SidA基因的表达已被完全抑制,该结果与嗜铁素含量测定结果相一致。不同pH条件下,SidA基因表达量均下降,pH 4.9时SidA基因表达量最高,该结果与嗜铁素含量测定结果相一致。而pH 10.9时CnA基因表达量未提高,原因可能是过碱条件不适合菌体生长。

图3 不同处理对棘孢木霉T6 SidA基因与CnA基因的影响Fig.3 Effect of different treatment on SidA gene and CnA gene of T. asperellum T6
3 讨 论
胁迫对木霉生长的影响是多方面的,既包括生长的变化,也包括各种生理生化等方面的变化。例如,随着NaCl浓度增加,哈茨木霉(T.harzianum) SH2303的生长速度、产孢量和几丁质酶活性明显下降[16]。本实验室前期研究发现,60 mmol/L的NaCl能够提高棘孢木霉Q1的酸性磷酸酶与碱性磷酸酶产量[17],但当NaCl浓度为200 mmol/L时,棘孢木霉Q1的嗜铁素产量受到抑制[18]。本研究发现NaCl浓度为300~1 000 mmol/L时不仅降低了棘孢木霉菌T6的嗜铁素产量,且NaCl浓度越高,嗜铁素产量越低。另外,高浓度CaCl2(10 mmol/L) 和高pH (pH 8.9) 对菌株T6嗜铁素产量也具有抑制作用,说明胁迫因子能够降低棘孢木霉菌T6的嗜铁素产量。
钙调磷酸酶基因参与真菌对逆境胁迫的响应。高盐条件下,酿酒酵母(Saccharomycescerevisiae) 胞内钙离子信号增强,钙调磷酸酶信号途径被激活,钙调磷酸酶基因表达上调[19]。李志勇等[20]利用实时荧光定量PCR方法分析了粟弯孢霉叶斑病菌(Curvularialunata) 中ClCna基因和ClCnb基因在非生物胁迫下的表达模式,发现在高浓度NaCl和山梨醇诱导下,ClCnb与ClCna基因表达量均升高,说明高盐和高渗透胁迫可以调控粟弯孢霉叶斑病菌钙调磷酸酶信号通路。本研究对SidA基因与CnA基因表达进行半定量PCR分析发现,在高浓度钠盐和高浓度钙盐胁迫下,棘孢木霉菌T6CnA基因表达上调,而SidA基因表达量下降。说明钠盐和钙胁迫可以调控棘孢木霉菌T6CnA基因与SidA基因。另外,高pH下钙调磷酸酶催化亚基基因表达量高于低pH,该结果与玉米黑粉病菌(Ustilagomaydis) 钙调磷酸酶2个亚基UmEna1和UmEna2基因受高pH 诱导[21]的结果相一致。
嗜铁素的合成常常受菌体内部信号途径的调控,目前国内外关于真菌钙调磷酸酶基因与嗜铁素基因相互关系的研究并不多。本实验室前期研究表明,sidA基因是棘孢木霉合成嗜铁素的关键基因之一,且参与调控木霉的生长和胁迫抗性[22]。在米根霉(Rhizopusoryzae) 中,钙调神经磷酸酶途径参与并调节缺铁胁迫[23],而缺铁胁迫又能促进嗜铁素的产生。本研究发现,钙调磷酸酶抑制剂CsA处理提高了嗜铁素产量,这与半定量PCR结果一致,即高盐胁迫和高pH下嗜SidA基因与CnA基因表达呈负调控关系,但具体分子调控机制仍有待进一步研究。