作用于配体门控氯离子通道3-羟基异噁(噻)唑衍生物的研究进展
2020-08-13钟慧勉巨修练刘根炎
肖 胜,王 倩,钟慧勉,巨修练,刘根炎
武汉工程大学化工与制药学院,湖北 武汉430205
3-羟基异噁唑和3-羟基异噻唑属于五元芳香杂环化合物,它们存在两种互变异构体(图1),3-羟基异噁(噻)唑(烯醇式)和异噁(噻)唑啉-3-酮(酮式)。在非极性溶液中它们主要以烯醇式结构存在,随着溶液极性增加,酮式结构逐渐成为主要存在形式[1-3]。3-羟基异噁(噻)唑芳环上的羟基有相对较高的酸性,这一性质使得它们在药物设计中可以作为羧基的生物电子等排体。3-羟基异噁唑和3-羟基异噻唑的pKa值分别为5.85和7.54,酸性的差异被认为是两种化合物及其类似物表现出不同药理或生理作用的主要因素之一[3-4]。
19世纪60年代中期,科学家们发现了两种天然的3-羟基异噁唑衍生物(图2):蝇蕈醇(muscimol)和鹅膏氨酸(ibotenic acid,IBA)。它们对γ-氨基丁酸(γ-aminobutyric acid,GABA)受体和谷氨酸(L-glutamic acid,glutamate)受体独特的生理活性引发了研究者的极大关注;药物化学家进一步修饰了其结构,合成了一系列具有药理活性的3-羟基异噁(噻)唑衍生物。GABA和glutamate(图2)分别是动物中枢神经系统重要的抑制性和兴奋性神经递质[5-6],它们与各自受体相互作用而广泛参与神经系统的各种功能[7-9]。

图1 3-羟基异噁(噻)唑的酮式和烯醇式的互变异构Fig.1 Keto and enol tautomerization of 3-hydroxyisoxazole and 3-hydroxyisothiazole
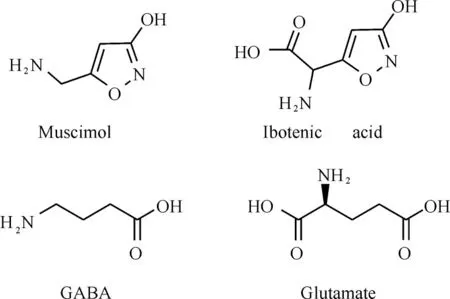
图2蝇蕈醇、鹅膏氨酸、γ-氨基丁酸和谷氨酸的化学结构Fig.2 Chemical structures of muscimol,ibotenic acid,GABA and glutamate
1 GABA和Glutamate门控氯离子通道
20世纪50年代,科学家首次从哺乳动物的脑萃取液中提取了GABA,并鉴定了其分子结构[10]。对一些哺乳动物和无脊椎动物体内GABA的含量测试结果表明,GABA在抑制性神经元中的含量远远高于其在兴奋性神经元中的含量。随后大量的研究表明GABA是动物神经系统中主要的抑制性神经递质[11-13],其作用靶标为GABA受体。Glutamate是动物中枢神经系统中主要的兴奋性神经递质,它与离子型谷氨酸受体(ionotropic glutamate receptors,iGluRs)和代谢型谷氨酸受体(metabotropic glutamate receptors,mGluRs)结合而发挥相应的生理作用[14]。
GABA受体广泛存在于动物的中枢和外周神经系统中,可分为离子型(A型和C型)和代谢型(B型)。A型和C型GABA受体属于半胱氨酸环配体门控离子通道家族[13],因此也被称为GABA门控氯离子通道。GABA通过动作电位触发的去极化从突触前神经元释放到突触间隙中,释放的GABA与位于突触后膜中的离子型GABA受体结合,引起突触后神经元的细胞膜的电位变化。GABA还可激活代谢型的GABA受体以引发细胞内的信号转导[13,15-16]。
离子型GABA受体(图3)由5个相同或不同的亚基构成,因此存在多个不同的亚型[17-19]。目前报道的哺乳动物离子型GABA受体的亚基有19个(α1-6、β1-3、γ1-3、δ、ε、π、θ和ρ1-3),它们可以形成同源或异源的五聚体[17]。存在于哺乳动物大脑中的主要的GABA受体(A型)亚型是由两个α1亚基、两个β2(或β3)亚基和一个γ2亚基构成[20]。含有ρ亚基的离子型GABA受体被归类为C型GABA受体,这种类型的受体往往被认为是一组特殊的A型GABA受体,它和A型GABA受体具有不同的药理特性。A型GABA受体每个亚基由约500个氨基酸残基组成,具有相同的构成部分:较长的N-末端细胞外结构,内含半胱氨酸二硫键环,由200多个氨基酸残基构成;4个跨膜序列(TM1-4);连接TM3和TM4区段的长而可变的细胞内环状结构。每个亚基的TM2结构构成氯离子通道内壁(图3)。GABA结合位点位于两个相邻亚基的N-端细胞外结构之间的正构位点。
B型GABA受体属于G蛋白偶联受体。这种类型的受体通过激活特异性偶联于效应蛋白的G蛋白转导信号,也被称为代谢型GABA受体[21]。B型GABA受体广泛分布在整个神经系统中,并且在一些细胞中与A型GABA受体协同发挥作用。B型GABA受体存在于突触前膜和突触后膜上。在突触前膜,B型GABA受体诱导钾电导并降低钙电导,从而抑制GABA在突触前膜中释放。在突触后膜上,B型GABA受体诱导钾电导,导致钾离子内流,进而引起缓慢的超极化。与其他G蛋白偶联受体相比,B型GABA受体与mGluRs具有高度相似的氨基酸序列[22-23]。
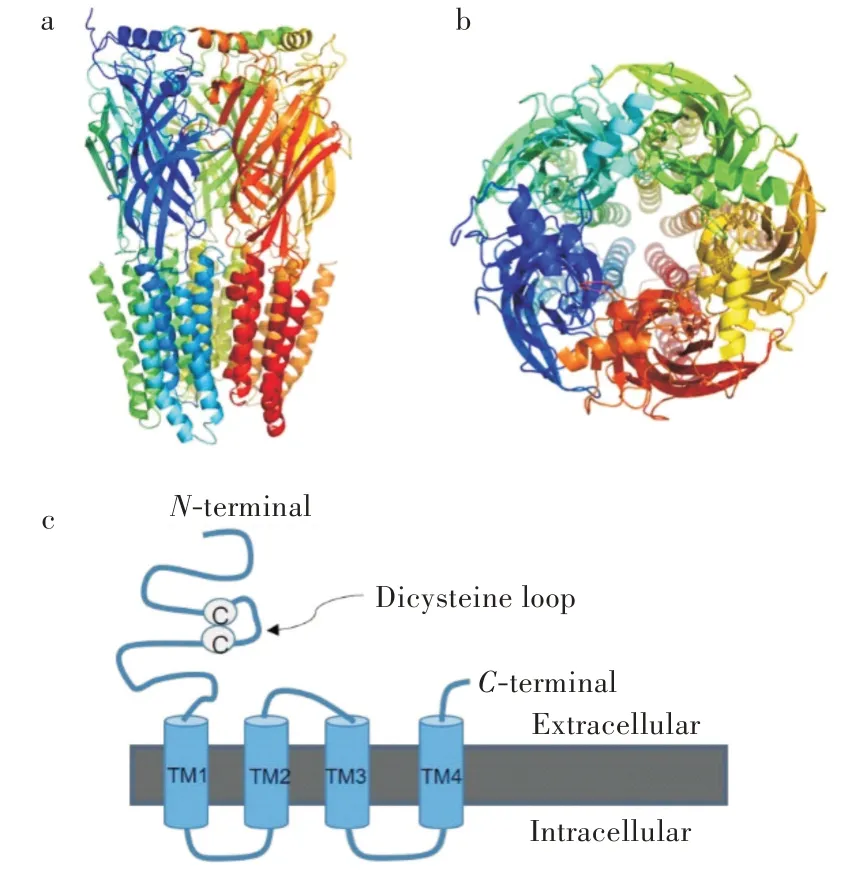
图3 A型GABA受体及其亚基图示:(a)A型GABA受体侧视图,(b)A型GABA受体俯视图,(c)A型GABA受体亚基示意图Fig.3 Schematic presentation of GABAA receptors and its subunit:(a)side view of GABAA receptor,(b)top view of GABAA receptor,(c)subunit of GABAA receptors
iGluRs属于配体门控离子通道家族,根据激动剂的不同的选择性可分为三类,N-甲基-D-天门冬氨酸受体(N-methyl-aspartatereceptor,NMDA受体),α-氨基-3-羟基5-甲基-4-异恶唑丙酸受体(α-amino-3-hydroxy-5-methyl-4-isoxazole-propionicacid receptor,AMPA受体),红藻氨酸受体(kainic acid receptor,KA受体)[24-25]。目前,NMDA受体有(NR1,NR2A-D,NR3A-B)7种亚单位被鉴定,AMPA受体有(GluR1-4)4种亚单位被鉴定,KA受体有(GluR5-7,KA1-2)5种亚单位被鉴定[24-26]。mGluRs是一个与G蛋白偶联的7次跨膜结构。目前,已经发现了8个mGluRs(mGluR1-8)亚基,根据它们氨基酸序列的相似性和信号转导路径可将其 分 为3类[27-28]。第I类mGluRs包 括mGluR1s和mGluR5s;第II类mGluRs包 括mGluR2s和mGluR3s;第III类mGluRs包括mGluR4s、mGluR6s、mGluR7s和mGluR8s。第II类和第III类mGluRs有相似的药理学性质,在突触前膜和后膜均有表达[27-28]。
2 Muscimol及其衍生物
Muscimol是一种天然化合物,最早是在毒蝇鹅膏菌(Amanita muscaria)中发现的[29]。从化学结构上看,muscimol是GABA的3-羟基异噁唑生物电子等排物,其对A型GABA受体表现出较强的激动活性[9,30-31]。此外,muscimol抑制神经元和神经胶质的GABA摄取,同时它也是GABA代谢酶和GABA转氨酶的底物。Thio-muscimol(图4)是人工合成的muscimol含硫类似物,表现出了与muscimol相类似的药理学特性[32-35]。

图4 Thio-muscimol的化学结构Fig.4 Chemical structure of thio-muscimol
研究表明muscimol是昆虫GABA受体的激动剂,其效价一般优于GABA。Liu等[9]对muscimol进行了结构修饰,将5位的氨基替换为酰胺基,合成了5-氨基甲酰基-3-羟基异噁唑(5-carbamoyl-3-isoxazolol,5-CIOL)。这一微小的结构变化使得化合物5-CIOL对家蝇(Musca domestica)GABA受体显示出拮抗而非激动活性。他们还进一步在5-CIOL异噁唑环的4位引入双环芳香基团,得到一系列4-芳基-5-氨基甲酰基-3-羟基异噁唑衍生物(图5),并测定了该系列衍生物对家蝇GABA受体4种剪接变异体(ac,ad,bc和bd)的活性。结果表明这一系列衍生物反而表现出明显的竞争拮抗作用,对4种剪接变异体的IC50值在微摩尔水平,其中5-甲酰氨基-4-(3-联苯基)-3-羟基异噁唑(化合物1,图5)活性最好,IC50值在30~107μmol/L间。其研究结果表明5-CIOL类衍生物属于昆虫GABA受体的竞争性拮抗剂,可作为先导化合物,用来进一步设计和开发作用于昆虫GABA受体的竞争性拮抗剂类杀虫剂。
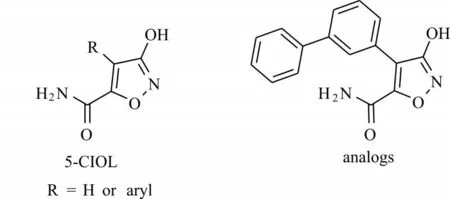
图5 5-CIOL及其衍生物的化学结构Fig.5 Chemical structures of 5-CIOL and its analogs
Muscimol和Thio-muscimol的单环衍生物对GABA受体各种亚型均具有激动或拮抗作用,研究表明将它们修饰为双环衍生物可以得到许多特异性A型GABA受体激动剂或拮抗剂,这些化合物对A型GABA各种亚型的亲和力有较大的区别[36]。4,5-二氢蝇蕈醇(4,5-dihydromuscimol,DHM)的S构型(S)-DMH(图6)是构造muscimol的双环衍生物的关键中间体,是一种具有选择性、高效的A型GABA受体激动剂,而(R)-DMH则是活性很低的GABA受体拮抗剂。4,5,6,7-四氢异噁唑[5,4-c]吡 啶-3-醇(4,5,6,7-tetrahydro-isoxazolo[5,4-c]pyridin-3-ol,THIP)和4,5,6,7-四氢异噁唑[4,5-c]吡啶-3-醇(4,5,6,7-tetrahydroisoxazolo[4,5-c]pyridin-3-ol,THPO)是两个典型的muscimol双环衍生物,其中THIP是一种特异且高效的A型GABA受体激动剂[37],而THPO是一种GABA受体拮抗剂,且对神经胶质运输系统具有一定选择性。Thio-THIP对脊髓中包含δ亚基的A型GABA受体激动作用较弱,但对大脑中包含δ亚基的A型GABA受体有明显拮抗作用。此外,研究表明THIP的 七元环状 类似物THIA对A型GABA受体没有明显的亲和力[38-41]。
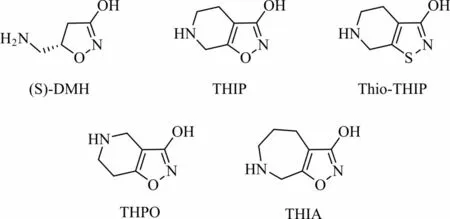
图6(S)-DMH、THIP、Thio-THIP、THPO和THIA的化学结构Fig.6 Chemical structures of(S)-DMH,THIP,Thio-THIP,THPO and THIA
3 4-PIOL及其衍生物
5-(4-哌 啶 基)-3-羟 基 异 噁唑[5-(4-piperidyl)-3-isoxazolol,4-PIOL]和5-(4-哌啶基)-3-羟基异噻唑[5-(4-piperidyl)-3-isothiazolol,Thio-4-PIOL]是两个典型的A型GABA受体部分激动剂[42],它们具有相似的药理学活性。Frølund等[42]发现4-PIOL衍生物对哺乳动物A型GABA受体有较好的竞争拮抗作用。在此研究基础上,Liu等[43]通过对Thio-4-PIOL的结构进一步修饰,探究了其对昆虫GABA受体的活性,发现在异噻唑环的4位引入芳香基团时,Thio-4-PIOL衍生物转化为了昆虫GABA受体的竞争性拮抗剂,且大的芳香基团更有利于提高其竞争拮抗活性。两个典型的Thio-4-PIOL衍生物4-(3-联苯基)-5-(4-哌啶基)-3-羟基异噻唑(化合物2,图7)和4-(2-萘基)-5-(4-哌啶基)-3-羟基异噻唑(化合物3,图7)对昆虫GABA受体表现出较高的竞争性拮抗效应,其对家蝇GABA受体IC50值分别为20.4和29.6μmol/L。部分Thio-4-PIOL衍生物在哺乳动物A型GABA受体中显示出纳摩尔级的竞争拮抗活性[42-43],与此相比,在昆虫GABA受体中该类化合物的活性仍然较低,因此针对这类化合物进行结构优化,有望开发出更有效和选择性的昆虫GABA受体竞争性拮抗剂。
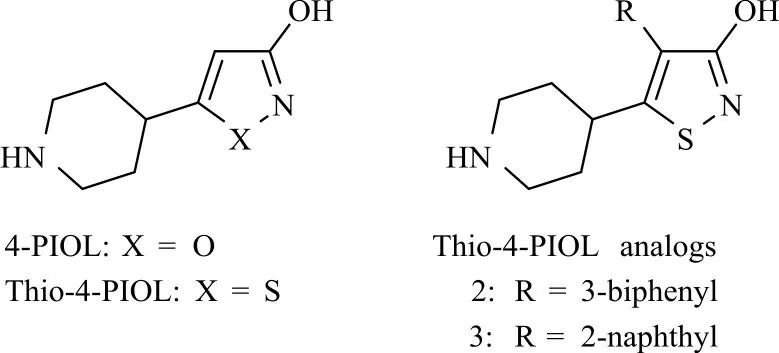
图7 4-PIOL和Thio-4-PIOL及其衍生物的化学结构Fig.7 Chemical structures of 4-PIOL,Thio-4-PIOL and Thio-4-PIOL analogs
4 IBA和Thio-IBA及其衍生物
IBA是一种天然存在的兴奋毒素,最初从毒蝇鹅膏菌中分离出来。由于其对iGluRs的激动作用,它已被广泛用作药理学以及神经毒理学工具[44-45]。IBA是glutamate的3-羟基异噁唑的类似物,它可以与KA受体(亲和力较低)、NMDA受体、第I类mGluRs及第II类mGluRs结 合,发挥 相应的作用。Thio-ibotenic acid(Thio-IBA)是IBA的含硫类似物,最初由Bunch等人合成得到。Thio-IBA表现出了与IBA显著的药理学差异,Thio-IBA和IBA作为激动剂均可以以相似的效率激活第I类mGluRs和 第II类mGluRs,有 趣 的 是 对 于 第III类mGluRs前者的激动效率是后者的500倍[45-47]。基于Thio-IBA的独特药理学活性,Jørgensen等[14]合成了一系列Thio-IBA类似物(图8),探究了它们对mGluRs的结合活性,结果表明,在异噻唑环4位引入小体积取代基(如氯、溴、甲基或乙基)的Thio-IBA的类似物保留了对第II类mGluRs的活性,但对第I类mGluRs和第III类mGluRs的结合活性丧失了。据此可进一步探究其构效关系,有望开发出新型特异性glutamate受体激动剂以及拮抗剂。
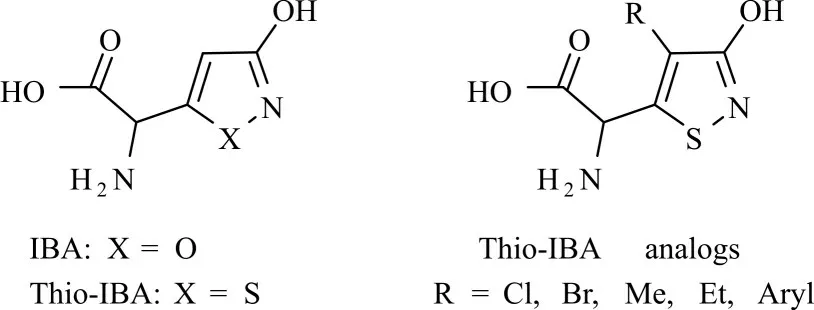
图8 IBA及其衍生物的化学结构Fig.8 Chemical structures of IBA and its analogs
5 结论
3-羟基异噁(噻)唑衍生物具有独特且丰富的药理和生理活性,它们对GABA和glutamate受体表现出一定的特异性和选择性。因此,选取典型的3-羟基异噁(噻)唑衍生物作为先导化合物,进行分子结构修饰,有望得到作用于GABA受体或glutamate受体的新型拮抗剂或激动剂,也可以对它们的生理药理活性进行更深入的研究。
