江苏省沉海圩乡村湿地水稻田种植模式转变后的生态恢复效果
2020-08-04罗祖奎李扬徐曦屈铭志马进孙文黄舒欣仝慧林
罗祖奎 李扬 徐曦 屈铭志 马进 孙文 黄舒欣 仝慧林

摘要: 为了检验沉海圩乡村湿地传统稻田转化为生态稻田之后的生态恢复效果, 在2018 年7 月—2019 年5 月对景观湖泊、生态稻田、传统稻田和居民区河道四类湿地的两栖类、底栖动物和水质进行野外调查及实验室处理. 结果表明: ①两栖类种类在景观湖泊、生态稻田、传统稻田和居民区河道依次减少; 两栖类数量在景观湖泊和生态稻田之间无显著差异, 但两者的数量均显著高于传统稻田和居民区河道. ②底栖动物种类和数量在稻田潮湿季节四类湿地之间均无显著差异; 但在稻田干旱季节生态稻田均处于最低水平.③生态稻田水质的有机氮和磷的指标在四类湿地中处于中等水平, 但叶绿素a 浓度最低. 总体而言, 沉海圩稻田种植模式转变后生态恢复效果在一定程度上得到了提高, 但缺乏科学管理不利于物种稳定, 针对生态稻田管理提出了建议.
关键词: 物种多样性; 水质; 生态稻田; 生态恢复; 生态管理
中图分类号: Q959.7;Q958.1 文献标志码: A DOI: 10.3969/j.issn.1000-5641.201931009
0 引 言
水稻是世界上的三大農作物之一,种植面积占全球农作物总面积的13%,其中,超过90%的水稻生产和消费均在亚洲国家,传统稻田大规模的使用农药促使害虫耐药性增强及其自然天敌的减少被认为是亚洲水稻田生态系统害虫爆发的主要原因,传统稻田大量使用农药和化肥对生物多样性及生态系统服务功能有负面影响,使用杀虫剂反而提高了害虫群落的多样性指数,增强了害虫群落的稳定性,现代农业需要在确保粮食增产的基础上,同时保护环境和生物多样性,为了增加水稻田的物种多样性,有学者提出了“生态稻田”的概念,生态稻田的功能在于提高生境多样化,使食物链趋于多层次州,其目的是将农业生产对生物多样性的负面影响降低到最小程度,生态稻田一般是通过生态工程构建景观多元化,提升生态稻田生态系统复杂性,抑制天敌的种类和数量,减少害虫的爆发。
同内外对生态稻田物种多样性及生态服务功能进行了大量的研究,比如:通过在田埂上种植开花植物提高生态稻田生境多样性,并对生态稻田和常规稻田中节肢动物进行比较;对水稻单作、稻一鱼和稻一鱼一鸭3种不同稻作方式下抑制杂草能力、稻飞虱虫数量、稻纵卷叶螟虫数量、蜘蛛数量的差异性以及抗稻瘟病效果进行比较,多位学者通过在田埂上种植高密度的多种陆生植物斑块构建生态稻田,对比生态稻田与传统稻田中鸟类等物种的结构差异性及生态服务功能差异性;通过构建水鸟所需斑块、水位和食物生境,对比生态稻田与工业稻田水鸟多样性和群落结构,施用植物制剂农药并释放赤眼蜂构建生态稻田,比较生态稻田与传统稻田的鱼类、两栖动物、软体动物、节肢动物和环节动物的差异性,虽然不同工程中构建生态稻田的处理方式不同,但是构建生态稻田的原理都是通过建造多样化的生境和多层次的食物链结构抑制害虫爆发。
从2010年起,江苏省苏州市开始实施生态补偿,水稻种植是重要的补偿对象,其中2014年和2015年就有24 653,9万元用于补偿水稻种植,占这两年补偿金总额的78%,沉海圩乡村湿地(后文简称沉海圩,见图1)属于苏州常熟市的生态丁程示范点之一,2012年开始,沉海圩打造“生态、生产、生活”三生融合的宜居、宜游的湿地乡村,生态工程中一项重要的议题就是水稻田生态修复示范,在2016年部分水稻种植由传统种植模式转变为生态种植模式,生态稻田具体操作如下:水稻从播种到收获期间均不施任何农药和肥料,每年水稻收割后留下较长的稻茬,通过翻耕掩埋这些稻茬作为底肥,水稻在生长期间进行一次集中人工除草,田埂上部分种植黄豆Glycine spp,等植物、部分让野草自然生长用来增加生境多样性,这些水稻田实施生态T程后,生态稻田的两栖类和底柄动物等物种多样性及物种组成与传统稻田的是否有差异?生态稻田的水质是否有提高?生态稻田在多种生境类型中的总体生态环境质量所处的位置如何?诸多的科学问题都有待探究,由于实施生态T程的时间不长,我们预判生态稻田生境质量提升及物种多样性增加效果不会太明显,本文研究的目的在于通过检验生态稻田的物种多样性及水质情况,分析生态稻田管理不足之处,为生态稻田的管理提供建议。
1 研究方法
1.1 研究区域概况
沉海圩(E 12°46'2.02'E 120°46'21.21”,N 31°43'38.28"~N 31°44'10.62";见图1)位于江苏省常熟市虞山镇谢桥福圩村,占地面积122 ha.其巾水稻田约53 ha.该区域地势低洼,水系发达.白2016年实施水稻田生态工程后沉海圩主要呈现4种湿地类型.(1)生态稻田.自2016年开始实施生态种植之前为传统稻田,每年在5-月种植水稻,其他月份土地闲置,11月底收割水稻后停止灌溉,直到翌年5月播种水稻才灌水.每年水稻收割后土壤被翻耕,将水稻茬掩埋作为肥料.(2)传统稻田.主要为稻一麦一年两熟制,在水稻种植期间通过沟渠引水灌溉.小麦种植期间土壤干旱,但干旱程度比同时期生态稻田略低.常用农药包括毒死蜱、醚菊酯、吡虫.噻嗪酮和丙溴磷等,常用化肥包括碳酸氢铵、磷酸二氢钾、氯化钾和过磷酸钙等.(3)景观湖泊.近岸生长有芦苇Phragmitcs australis、水花生Altcrnanthcra philoxcroidcs(Mart)Griscb、菱角Trapa bicornis Osbcck、莎草Cypcraccac spp、香蒲Typha angustata Bory et Chaub等植物,生态工程建设中义在近岸人为种植了睡莲科Nymphacaccaespp、茭白Zizania latifolia和千屈菜Lythrum salicaria Linn等植物,由于景观湖泊水较深且与外界隔开,水质相对较好,类似于自然湿地.(4)居民区河道.大部分河道都修建了水泥堤岸,无滩涂,水生植物主要为沉水植物.为了净化水质,工程后期小部分河道人工种植了茭白和菱角等,实施“植物浮岛”技术降解污染.沉海圩的自然地理条件见文献[15].
1.2 野外调查及实验室处理
1.2.1 生境斑块及水体连通性调查
于2018年7月用无人机航拍获得沉海圩120 m垂直正射照片(图片精度为5 cm).按照航拍照片划分生境斑块(见图1).对所有生境斑块边界的水体联通情况进行调查,发现除了居民区河道的水体有局部与外界相通,其他生境的水体与外界均不连通,每种生境内部水体根据堤岸边界呈现小斑块分布.在2019年4月底实施水体连通工程,将不同类型的湿地之间以及同种湿地不同斑块之间的堤岸进行连通.
1.2.2 两柄类调查
2018年7月和8月、2019年5月共3次埘4类湿地生境巾的两柄类进行调查.每次记录湿地生境岸边1 m及水巾2 m,累汁宽3 m,长150 m的样线巾两柄类种类和数量.每类生境每次调查6 7条样线,3次累汁调查20条样线.日调查时间在19:30~23:OO进行.
1.2.3 底柄动物调查及处理
2018年11月和2019年4月进行底柄动物调查.生态稻田和传统稻田用平口铁锹挖泥块.景观湖泊和居民区河道水相对较深,用抓斗采泥器在距離岸边20~50 cm的地方抓泥.四类生境取泥块长×宽×高均为10 Cm×10cm×10 Cm 11月,调查期间水稻已经成熟但没有收割,生态稻田和传统稻田均比较潮湿.同一季节同一生境巾的泥块采集间距至少20 m以上随机采集.虽然4月份生态稻田(闲置,泥土被翻耕)和传统稻田(种植小麦)都没有水,也与其他湿地生境采集了同样数量的泥块.同一季节四类生境每次均采集10个泥块,用60日不锈钢网筛现场筛洗泥块巾的底柄动物,装入塑料瓶并用75%的酒精浸泡,带同实验室后再次清洗并参照专业书籍鉴定.鉴定完毕的底柄动物用烘箱(型号:LH 9023A 340 Cm×325 cm×300 CIII)在80℃温度条件下进行24 h的烘干处理,烘箱内放人干燥剂.烘箱冷却2 h后取出底柄动物样品用电子秤称重(精确度0.1g).
1.2.4 水质采集
分别在2018年的7月、8月和¨月以及2019年的1月和4月进行水样采集.结合实际情况,每次在每个堤岸形成的隔离斑块巾取一份水样.由于每类生境堤岸隔离形成的斑块数相差较大,而且水稻田季节性干涸或者水太浅导致尤法采集水样,因此,四类生境巾水样样本数相差较大,在生态稻田、传统稻田、景观湖泊和居民区河道分别采集到6份、6份、15份和49份样本.生态稻田和传统稻田一般取表层水,而景观湖泊和居民区河道取水深度为0.3~0.5 m.每次在距离岸边0.5 m的位置用玻璃采样器采集水样.封闭并用保存剂同定带同实验室静置24 h再进行监测,水质指标包括:pH值、氧化还原电位(oPR)、浊度(TURB)、氦氮(NH3-N)、亚硝酸盐氮(N02-N)、硝酸盐氮(N03-N)、叶绿素a浓度(chl-a)、溶解氧(Do)、总磷(TP)、商锰酸钾(cODM。)指数,各指标分析方法参考相关文献.
1.3 数据分析
因为底柄动物采集分别在11月(水稻未收割且稻田潮湿)和4月(稻田干旱)两个时间段,稻田的水分布情况不同,可能对底栖动物的分布有影响,所以在比较四类生境之间底栖动物的种类、数量和干重时将11月和4月的数据分开比较.
检验不同生境之间两柄类数量差异性、分别检验4月(稻田干旱)不同生境之间底柄动物数量和干重差异性时,数据均进行转换使之同时满足正态分布和方差齐性要求,转换后的数据用One-wayANOVA进行分析,不同生境之间变量两两比较均用LSD法.检验不同生境之间两柄类种类差异性、检验4月(稻田干旱)的底柄动物种类在不同生境之间差异性、分别检验11月(稻田潮湿)底柄动物种类、数量和干重在不同生境之间差异性时,数据均不能满足正态分布要求.全部选用Kruskal-wallis H检验.
全文平均值表示为Mean土SD,显著水平α=0.05.数据分析在Spss 19.0上完成,作图用MicrosoftExcel 2003.
2 结果与分析
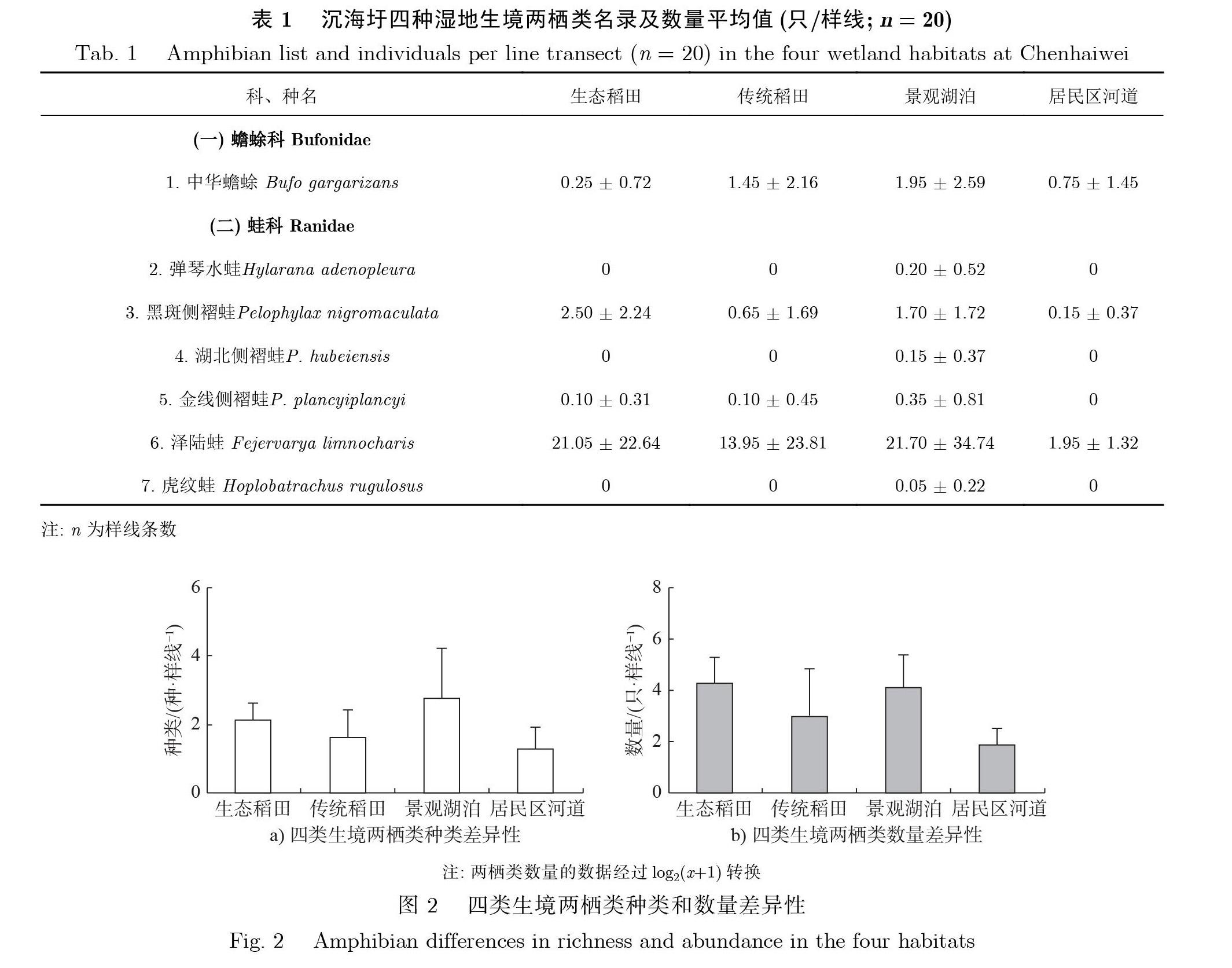
2.1 两栖类
共记录到7种两柄动物(见表1).四类生境之间两栖类种类存在显著差异(x2=23.55,df=3,p < 0.001; Kruskal-wallis H 检验; 见图2), 平均值从高到低依次为景观湖泊、生态稻田、传统稻田和居民区河道. 其中, 弹琴水蛙Hylarana adenopleura、湖北侧褶蛙Pelophylax hubeiensis 和虎纹蛙Hoplobatrachus rugulosus 只在景观湖泊有记录. 金线侧褶蛙Pelophylax plancyiplancyi 在居民区河道没有记录, 在其他3 种生境中都有记录. 中华蟾蜍 Bufo gargarizans、黑斑侧褶蛙Pelophylaxnigromaculata 和泽陆蛙 Fejervarya limnocharis 在四类生境中均有记录. 四类生境之间两栖类数量存在显著差异(F(3, 76) = 15.59, p < 0.001; One-way ANOVA); 景观湖泊和生态稻田两者之间两栖类数量无显著差异, 但两者的两栖类数量均显著高于传统稻田和居民区河道; 传统稻田的两栖类数量显著高于居民区河道. 生态稻田中数量最多的是泽陆蛙, 占生态稻田蛙类总数量的88%. 生态稻田中的黑斑侧褶蛙在四类生境中最丰富(见表1). 总体而言, 生态稻田的两栖类多样性低于景观湖泊、与传统稻田相近、高于居民区河道.
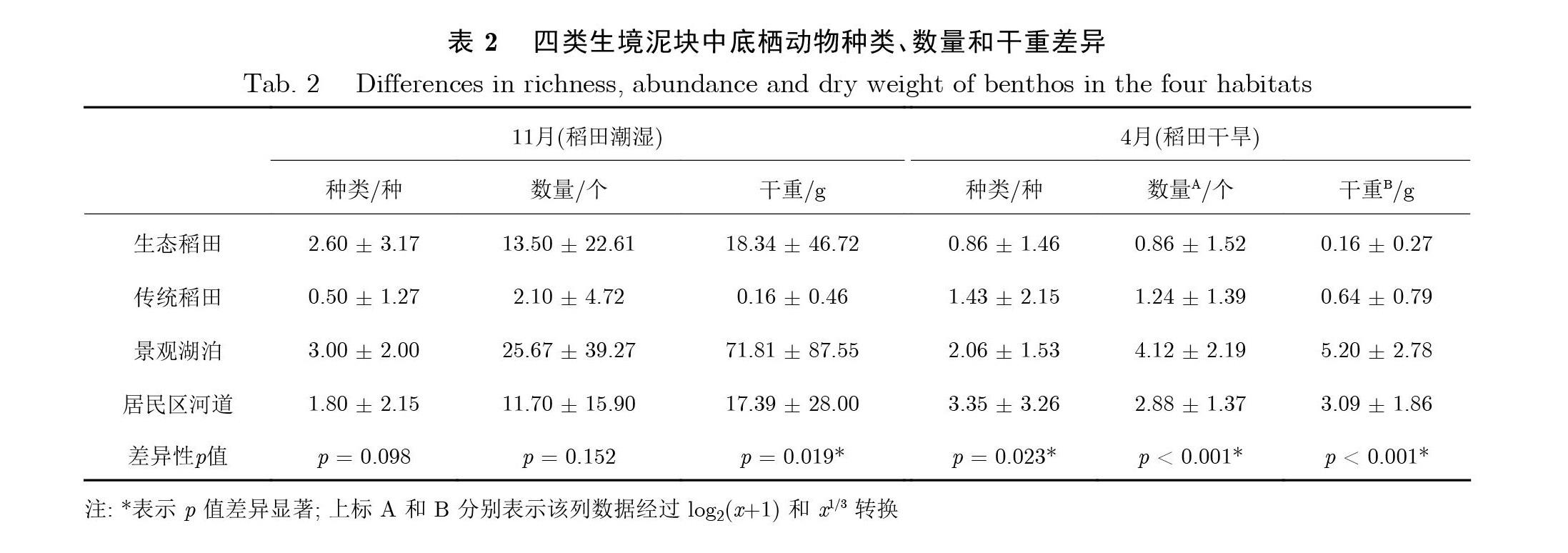
2.2 底栖动物
11 月(稻田潮湿), 四类生境泥块中底栖动物种类平均值(c2 =6.30, df = 3, p = 0.098; KruskalwallisH 检验; 见表2) 和数量平均值(c2 = 5.28, df = 3, p = 0.152) 均无显著差异. 但是, 底栖动物干重平均值有显著差异(c2 = 9.90, df = 3, p = 0.019), 干重平均值从高到低依次为景观湖泊、生态稻田、居民区河道和传统稻田.
4 月(稻田干旱), 四类生境之间底栖动物种类平均值存在显著差异(c2 = 9.49, df = 3, p = 0.023;Kruskal-wallis H 检验; 见表2), 平均值从高到低依次为居民区河道、景观湖泊、传统稻田和生态稻田.底栖动物数量平均值存在显著差异(F(3, 36) = 8.02, p < 0.001; One-way ANOVA), 景观湖泊显著高于居民区河道、传统稻田和生态稻田; 居民区河道显著高于传统稻田和生态稻田; 传统稻田和生态稻田之间无显著差异. 四类生境之间底栖动物干重平均值存在显著差异(F(3, 36) = 14.12, p < 0.001), 景观湖泊显著高于居民区河道、传统稻田和生态稻田; 居民区河道显著高于传统稻田和生态稻田; 传统稻田和生态稻田之间无显著差异.
总体而言, 在稻田潮湿季节(11 月), 四类生境中底栖动物的多样性差异性不显著, 但底栖动物多样性明显受到缺水(4 月) 的影响.
2.3 水 质
pH 值在四类生境之间存在显著差异(F(3, 72) = 4.61, p = 0.005; One-way ANOVA; 见表3), 生态稻田显著高于其他三类生境, 传统稻田、景观湖泊和居民区河道三者之间无显著差异. NO2-N 在四类生境之间存在显著差异(c2 = 10.70, df = 3, p = 0.013; Kruskal-Wallis Test 检验), 平均值从高到低依次为: 传统稻田、居民区河道、生态稻田、景观湖泊. Chl-a 浓度在四类生境之间存在显著差异(F(3, 72) =4.66, p = 0.005), 居民区河道显著高于其他三类生境, 生态稻田、传统稻田和景观湖泊之间无显著差异. DO 在四类生境之间存在显著差异(F(3, 72) = 31.77, p < 0.001), 生态稻田和传统稻田之间无显著差异, 但两者均显著高于景观湖泊和居民区河道, 景观湖泊与居民区河道之间无显著差异; TP 在四类生境之间存在显著差异(F(3, 72) = 4.87, p = 0.004), 传统稻田与生态稻田之间无显著差异, 但传统稻田显著高于景观湖泊和居民区河道, 生态稻田、景观湖泊和居民区河道之间无显著差异. CODMn 指数在四类生境之间存在显著差异(F(3, 72) = 11.30, p < 0.001), 生态稻田显著高于其他三类生境, 传统稻田、景观湖泊和居民区河道之间无显著差异. 四类生境之间的ORP(F(3, 72) = 0.50, p = 0.682)、TURB(F(3, 72) = 0.97, p = 0.413)、NH3-N(F(3, 72) = 1.88, p = 0.140) 和NO3-N(c2 = 3.89, df = 3, p =0.273) 差异均不显著. 总体而言, 生态稻田的水质优于传统稻田和居民区河道, 但次于景观湖泊.
3 讨论
3.1 物种多样性影响因素
(1) 两栖类多样性. 由于生态稻田人为设计增加了生境和食物链的复杂程度, 为天敌提供了栖息场所, 大量研究表明生态稻田比传统稻田有更高的生物多样性及更多的天敌[1-2,5-6,19], 但是, 生態稻田和传统稻田在水生植物、两栖类和大型无脊椎动物等都无法超过临近自然湿地中的物种多样性[19]. 来自沉海圩的两栖类研究得出与前人研究相似的结论, 生态稻田中两栖类多样性总体上高于传统稻田和居民区河道, 次于景观湖泊(类似于自然湿地). 沉海圩的生态稻田不使用农药和化肥并增加稻田四周田埂上植被多样性对提高两栖类多样性有利, 但是, 生态稻田在生境复杂程度、水质(见表3) 和常年水位恒定性等方面都次于景观湖泊, 食物链复杂且水位恒定有利于多种两栖类与其他物种共存[9,19-20],所以, 沉海圩的生态稻田闲置期不灌水对两栖类多样性保护不利. 因皮肤辅助呼吸的需要, 两栖类对水的依赖性强, 移动能力弱, 导致黑斑侧褶蛙和金线侧褶蛙等成为栖息地的“忠实”物种[20], 稻田干旱容易导致两栖类死亡, 也不利于蛙类产卵, 重新灌水后外界两栖类重新迁入的速度也慢. 沉海圩实施了水体连通工程, 内陆水体连通降低了水体之间的隔离程度, 缩短了水体的距离, 为两栖类提供了更多的栖息地生境[20]. 水质污染会影响两栖类的结构与分布[20], 景观湖泊在四类生境中的氮和磷的各项指标均最低(见表3), 表明其是四类生境中受到面源污染程度相对最小的生境, 这可能是只在景观湖泊中记录到弹琴水蛙、湖北侧褶蛙和虎纹蛙的原因. 金线侧褶蛙[20-21] 和黑斑侧褶蛙[21] 的分布往往与池塘边植被盖度正相关, 景观湖泊中的植被种类在四类生境中最为复杂, 对提高这两种蛙的数量有利,但是居民区河道为陡峭的水泥堤岸, 没有滩涂及滩涂植被, 对金线侧褶蛙和黑斑侧褶蛙等需要岸边植被浓密的两栖类分布不利.
(2) 底栖动物多样性. 水稻田的土壤动物大多属于表聚性物种, 75% 以上的个体都分布在0 ~ 10 cm的土层, 种类和数量随土层加深急剧减少[22]. 沉海圩的生态稻田和传统稻田缺水期较长, 干旱对居于土壤表层的底栖动物多样性有负面影响[23]. 底栖动物直接或者间接以有机质为食[24], 沉海圩的生态稻田不使用任何肥料, 仅靠前一年的水稻茬为底肥导致肥力不足, 这不仅不利于底栖动物群落的发展,而且实地观察发现生态稻田的水稻生长情况不及传统稻田的生长情况好. 粪肥的长期施用对提高土壤有机质的碳源和氮源有重要作用, 这不但为底栖动物提供了食物基质, 也为土壤微生物提供了食物基质, 而土壤微生物的增加又为底栖动物提供了新的食物来源[25]. 但是, 夏季由于高温的原因, 有机质的含量却对小型底栖动物的丰富度有负面影响[24], 所以使用粪肥对传统稻田的底栖动物的影响可能与季节有关.
[ 9 ] 魏波. 从生态系统角度谈水稻田病虫害的综合防治 [J]. 中国科技财富, 2009(2): 121. DOI: 10.3969/j.issn.1671-461X.2009.02.107.
[10]HORGAN F G, RAMAL A F, VILLEGAS J M, et al. Ecological engineering with high diversity vegetation patches enhances birdactivity and ecosystem services in Philippine rice fields [J]. Regional Environmental Change, 2017, 17(5): 1355-1367. DOI: 10.1007/s10113-016-0984-5.
[11]王凯学, 张清泉, 陈丽丽, 等. 生态稻田及常规稻田节肢动物群落结构特征的比较研究 [J]. 植物保护, 2013, 39(3): 31-35. DOI: 10.3969/j.issn.0529-1542.2013.03.007.
[12]张丹, 成升魁, 杨海龙, 等. 传统农业区稻田多个物种共存对病虫草害的生态控制效应—以贵州省从江县为例 [J]. 资源科学, 2011, 33(6): 1032-1037. DOI: CNKI: SUN: TRZY.0.2011-06-005.
[13]谢汉宾, 莫英敏, 张姚, 等. 以水鸟保育为目标的水稻田构建技术及效果评估 [J]. 长江流域资源与环境, 2017, 26(11): 1919-1927. DOI: 10.11870/cjlyzyyhj201711021.
[14]张秀菊, 龙媚, 闻振东, 等. 常熟市水生态补偿现状及问题探讨 [J]. 人民长江, 2017, 48(10): 13-17. DOI: 10.16232/j.cnki.1001-4179.2017.10.004.
[15] 苏永春, 勾影波, 郁达, 等. 江苏常熟虞山土壤动物群落多样性研究 [J]. 生物多样性, 2004, 12(3): 333-338. DOI: 10.3321/j.issn:1005-0094.2004.03.005.
[16]齐钟彦, 马绣同. 中国动物图谱(软体动物, 第4册) [M]. 北京: 科学出版社, 1985.
[17] 刘文亮, 何文珊. 长江河口大型底栖无脊椎动物 [M]. 上海: 上海科学技术出版社, 2007.
[18]霍元子, 何文輝, 罗坤, 等. 大型溞引导的沉水植被生态修复对滴水湖水质的净化效果 [J]. 应用生态学报, 2010, 21(2): 495-499. DOI: 1001-9332(2010)02-0495-05.
[19]MALTCHIK L, STENERT C, BATZER D P, et al. Can rice field management practices contribute to the conservation of species fromnatural wetlands? Lessons from Brazil [J]. Basic and Applied Ecology, 2017(18): 50-56. DOI: 10.1016/j.baae.2016.10.002.
[20]梁国付, 李成蹊, 白鑫, 等. 农业景观中环境因素对两栖类生物分布的影响 [J]. 应用生态学报, 2019, 30(1): 301-308. DOI: 10.13287/j.1001-9332.201901.008.
[21]吴迪, 岳峰, 罗祖奎, 等. 上海大莲湖湖泊湿地两栖动物群落分布及生境选择模式 [J]. 复旦学报(自然科学版), 2011, 50(3): 268-273.DOI: 10.1007/s11783-010-0264-4.
[22]林兰稳, 钟继洪, 谭军, 等. 土地利用方式对土壤动物多样性的影响 [J]. 生态环境学报, 2012, 21(10): 1678-1682. DOI: 10.3969/j.issn.1674-5906.2012.10.008.
[23]罗祖奎, 李扬. 钳嘴鹳觅食生境选择及食物特征分析 [J]. 西北师范大学学报(自然科学版), 2019, 55(4): 1-8. DOI: 10.16783/j.cnki.nwnuz.2019.04.014.
[24]刘均玲, 袁超, 何永姑, 等. 东寨港红树林小型底栖动物丰度与Chla、有机质的相关性 [J]. 生态学报, 2019, 39(1): 189-195. DOI: 10.5846/stxb201711262113.
[25]郭莹, 王一明, 巫攀, 等. 长期施用粪肥对水稻土中微生物群落功能多样性的影响 [J]. 应用与环境生物学报, 2019, 25(3): 593-602.DOI: 10.19675/j.cnki.1006-687x.2018.09028.
[26]XING G, CAO Y, SHI S, et al. N pollution sources and denitrification in waterbodies in Taihu Lake region [J]. Science ChinaChemistry, 2001, 44(3): 304-314. DOI: 10.1007/BF02879621.
[27]熊正琴, 邢光熹, 沈光裕, 等. 太湖地区湖、河和井水中氮污染状况的研究 [J]. 农村生态环境, 2002, 18(2): 29-33. DOI: 10.3969/j.issn.1673-4831.2002.02.007.
[28]谢迎新, 邢光熹, 熊正琴, 等. 常熟地区河湖水体的氮污染源研究 [J]. 农业环境科学学报, 2006, 25(3): 766-771. DOI: 10.3321/j.issn:1672-2043.2006.03.044.
[29] 孙菲, 袁鹏, 程建光, 等. 宜兴市殷村港叶绿素a与影响因子的多元分析 [J]. 环境工程, 2017, 35(9): 53-57. DOI: 10.13205/j.higc.201709011.
[30]于婷婷, 徐奎栋. 长江口及邻近海域秋冬季小型底栖动物类群组成与分布 [J]. 生态学报, 2013, 33(15): 4556-4566. DOI: 10.5846/stxb201204280613.
(责任编辑: 张 晶)
