金沙江流域不同海拔处云南松生态弹性及生长衰退过程*
2020-07-28申佳艳李帅锋黄小波王绍武苏建荣
申佳艳 李帅锋 黄小波 王绍武 苏建荣
(1. 中国林业科学研究院资源昆虫研究所 昆明 650224; 2. 国家林业和草原局云南普洱森林生态系统国家定位观测研究站 普洱 665000; 3. 云南省永仁县林业和草原局 永仁 651400)
近100年来,气候变暖背景下区域极端气候事件发生的频率和强度正逐年增加,已受到全球广泛关注(Plummeretal., 1999; Goswamietal., 2006; Dai, 2013)。大量研究发现,频发的气候干扰事件导致森林衰退频繁发生(Allenetal., 2010; Zhangetal., 2014; Liangetal., 2015; Andereggetal., 2016; Cailleetal., 2017; Gaoetal., 2018),且以极端干旱造成的影响最严重(Zhangetal., 2014; Yangetal., 2018)。大部分极端干旱对树木生长和大尺度森林生态系统影响的研究表明,干旱会增加林木死亡率,减弱森林生产力及碳汇能力(Tangetal., 2004; Borkenetal., 2006; Missonetal., 2010; Suseelaetal., 2015)。这种作用在接近于森林分布极限的干旱和半干旱区尤为明显,树木呈现出径向生长减弱、死亡率增加等不同程度的生长衰退现象(Abramsetal., 1998; Eilmannetal., 2007; 2011; Phillivsetal., 2009; Dimitriosetal., 2010; Zhangetal., 2014; Liangetal., 2015; Yangetal., 2018)。
生态弹性是树木抵抗干扰及受干扰后恢复到原初状态的能力(Thompsonetal., 2009)。准确评估森林对当前和过去气候变化的反应有助于理解森林对未来气候变化的适应能力。树木生态弹性研究是极端干扰事件频发背景下森林生长动态研究的必要内容 (Law, 2014),而长时间尺度的树木生长变化数据可用于量化树木的生态弹性。在抵抗力弱的区域内,极端干扰容易引发树木的生长衰退(Xuetal., 2016)。树木对干旱的抵抗力及恢复力是评定其干旱脆弱性的重要指标(Vicente-Serranoetal., 2012),干旱会通过影响树木的生态弹性使其生长恢复过程产生差异(Huangetal., 2018)。
虽然利用遥感观测等手段能有效反映当前环境变化下的森林变化特征及趋势(于伯华等, 2009; Zhaoetal., 2012),但是气候与树木生长在年际间和年内尺度上的巨大差异对树木生长动态研究方法提出了更高要求。当前,树木年轮方法已广泛用于树木生长对干扰事件和环境胁迫的响应(Helmanetal., 2016; Gazoletal., 2018)、森林衰退原因(Beieretal., 2008; Gazoletal., 2016; Vitalietal., 2017)、年际间森林生长动态评估等研究(Lorimeretal., 1989; Nowackietal., 1997); 利用树木年轮数据可有效鉴定森林衰退的过程和特点(Frittsetal., 1989),分析树木生长受抑制的程度、时间长短及空间范围(Amorosoetal., 2012),进而揭示森林衰退机制。
近100年来,我国西南地区极端干旱事件频发,对森林生长动态造成严重影响(李强子等, 2010; Yangetal., 2012; 李忆平等, 2015; 赵志平等, 2015),但目前有关极端干旱对森林生长影响的研究甚少。作为长江中下游的重要生态屏障,金沙江流域森林的长期健康动态研究显得愈发重要。云南松(Pinusyunnanensis)是西南地区的乡土树种,主要分布在极端干旱影响最明显的西南地区西部。本研究以生态脆弱的金沙江流域云南松为对象,通过树木年轮学方法建立径向生长与气候因子的关系,探究限制该区云南松径向生长的主要气候因子以及不同海拔处云南松应对极端干旱的生态弹性,以期揭示该区云南松生长衰退的机制和原因,预测森林健康阈值,从而对该区森林进行更加有效的保护。
1 研究区概况
研究区位于云南高原北部边缘的永仁县白马河林场内(101.32°—101.87° E,25.87°— 26.53° N),是金沙江流域云南松的典型分布区。永仁县地处云南省与四川省交界处,地势西北高东南低,以中低山丘陵为主,研究区海拔926~2 884.7 m,相对高差1 958 m。金沙江流经永仁境段,从永仁与大姚交界的顺山滩起至与元谋交界的母猪滩止,总长157 km。气候为北亚热带西南季风型,冬春干燥、夏秋多雨,干湿季分明,因相对高差较大而立体气候明显。年均气温17.4 ℃,最冷月(1月)平均气温1.9 ℃,最热月(5月)平均气温28.9 ℃,极端最高气温35.2 ℃,极端最低气温-4.4 ℃; 年降水量884 mm,6—10月降水量占全年的90%(图1),年均日照时长2 824.4 h。主要土壤类型为黄红壤、中性紫色土、红壤和酸性紫色土。优势树种为云南松,伴生树种有滇青冈栎(Cyclobalanopsisglaucoides)、尼泊尔桤木(Alnusnepalensis)、厚皮香(Ternstroemiagymnanthera)和高山栲(Castanopsisdelavayi)等。

图1 永仁县气象站月均气温、月均最高气温、月均最低气温和月降水量年内变化(1952—2016)Fig.1 Monthly mean temperature, monthly mean maximum temperature, monthly mean minimum temperature, and monthly precipitation in Yongren Meteorological station(1952-2016)
2 研究方法
2.1 样品采集与年表建立
按国际树木年轮库(ITRDB)标准及树木年轮采样原则与方法(Fritts, 1971),于2018年11月在高海拔(2 740 m,云南松天然次生林,伴生滇青冈栎,树龄60~130年,树间距3~7 m,坡度25°,坡向西南)、中海拔(2 340 m,云南松天然林,伴生滇青冈栎、厚皮香,树龄80~160年,树间距10~15 m,坡度10°,南坡)和低海拔(1 845 m,云南松天然次生林,伴生滇青冈栎、高山栲,树龄45~60年,树间距3~5 m,坡度15°,坡向西南)3个海拔段,随机选取生长微环境大致相同、健康、龄级较大、树木间相互遮蔽较小、立地条件较差的云南松(对气候变化敏感性更强),在胸高处(1.3 m)不同方位钻取2根年轮样芯(取样方向首选南向,2根样芯方向垂直),3个海拔段共选取70株云南松样树,采集了140根样芯。其中高海拔云南松20株(胸径28.3~49.6 cm)、样芯40根,中海拔云南松30株(胸径41.0~64.5 cm)、样芯60根,低海拔云南松20株(胸径26.7~45.5 cm),样芯40根。
采集的样芯按树木年轮样品处理流程处理。自然风干后用白乳胶固定在样本槽内,用不同粗细的砂纸逐级打磨至光、滑、亮状态,且能在40倍显微镜下清晰观察到年轮轮廓。在双筒显微镜下目测交叉定年后,用LINTAB 6.0 年轮宽度测量仪(精度为0.001 mm)(Rinntech, Heidelberg, Germany)测量树轮原始宽度。通过比较不同树木个体间的年轮序列,鉴别出缺轮、伪轮及测量误差,确定样芯上每个年轮准确的生长年份,最后在COFECHA程序中(Holmes, 1983)检验交叉定年结果,剔除与主序列相关性差、年轮序列不完整及难以交叉定年的样芯,最终剩余55株样树的109根样芯进入主序列(表1)。用ARSTAN程序(Cook,1985)中的负指数函数或样条函数拟合去除因遗传因素或个体干扰产生的抑制、释放等生长趋势后,最终得到各海拔段云南松标准年表(STD,图4)。各海拔段标准年表公共区间统计特征如表1所示,3个年表的样本总体代表性(EPS)均大于0.85,能较好地代表研究区各海拔段云南松树轮宽度变化的基本特征。
2.2 气象数据
因研究区气象站器测资料不完整,采用荷兰皇家气象局服务网(https:∥climexp.knmi.nl/start.cgi)CRU网格点数据。本研究采用CRU TS 3.22的区域月均气温、月降水量和帕尔默干旱指数。帕尔默干旱指数(PDSI)用于衡量大气水分供应和土壤水分需求的累积效应,是综合考虑了降水、温度和蒸发的水分条件指标。鉴于树木生长对气候响应存在“滞后效应” (Frittsetal., 1965),故选取上年5月至当年10月的气象数据,共计51个气象变量,分析标准年表数据的气候响应。
2.3 数据处理
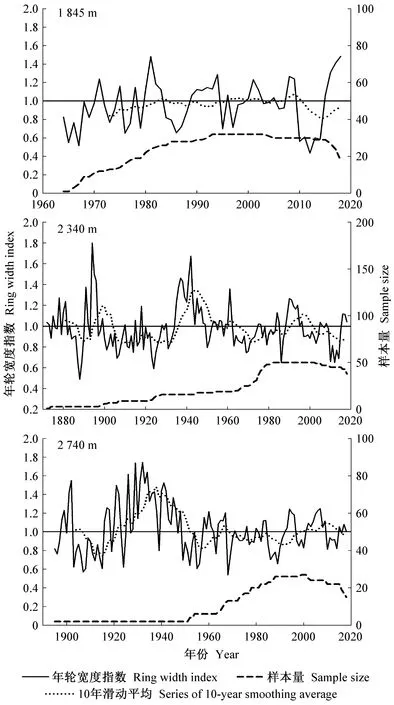
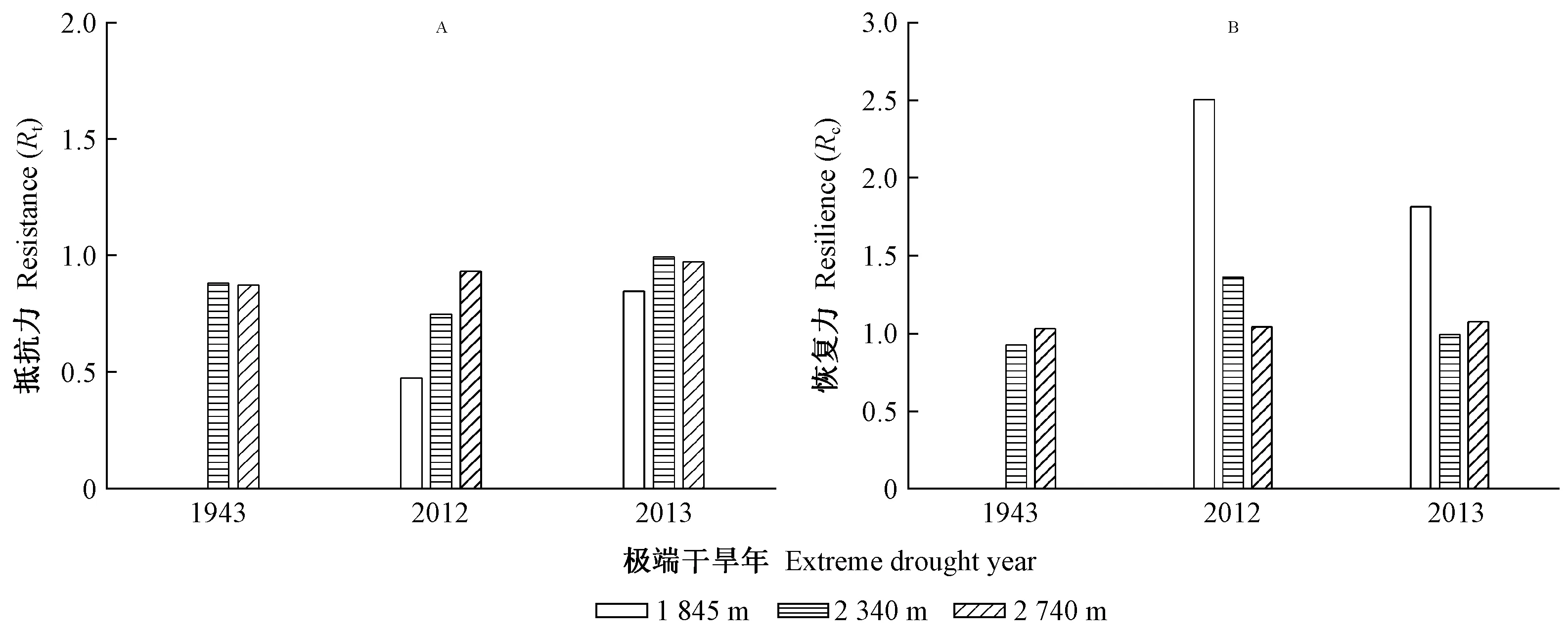
研究区降水集中于6—10月份,而11月至翌年5月降水极少,是干旱发生的主要时期。因此利用上年11月至当年5月的平均帕尔默干旱指数来定义研究区的极端干旱事件。根据该指数干旱等级评定标准: -0.5 利用树木径向生长变化率判定云南松是否发生生长抑制,计算公式如下(Payetteetal., 1990; Nowackietal., 1997): GCi=(M2-M1)/M1。 式中: GCi为树木在第i年表现出的前后各5年的径向生长变化率,为减小个别年份生长变化对生长抑制事件检测的影响,以5年为滑动平均计算云南松径向生长变化率,M1和M2分别表示前5年(包括当年)和后5年(不包括当年)树轮宽度指数的平均值。径向生长变化率低于-25%表示树木生长受到抑制和发生了衰退,超过75%表示树木生长释放。 利用Lloret等(2011)的公式,计算云南松的抵抗力和恢复力: Rt=Gr/Gb; Rc=Ga/Gr。 式中:Rt和Rc分别为树木的抵抗力和恢复力;Gr为极端干旱事件当年的树轮宽度指数;Gb、Ga分别为干旱前、后4年的平均树轮宽度指数值。Rt大于0.75表示对极端干旱有高抵抗力,Rc大于1.25表示在极端干旱事件后仍有较强的恢复能力。 运用Dendroclim2002软件中的响应函数(Biondietal., 2004)分析树轮宽度指数与气温、降水量及帕尔默干旱指数间的响应关系。利用CANOCO 4.5软件(Braak, 1994)中的冗余分析(RDA),进一步验证云南松径向生长与气候因子间的关系。用Sigmplot 10.0软件作图。 如表1所示,样本总体代表性为0.861~0.962,均大于树木年轮分析中总体代表性阈值0.85。年表的一阶自相关系数为0.443~0.681,说明该区云南松上年径向生长量对当年生长有明显影响,且影响力随海拔升高而增强。第一特征向量百分比达到31.25%~36.04%,信噪比达到5.53~10.13,说明这3个年表都包含了大量气候信息,年表质量可靠。 表1 不同海拔云南松树轮宽度标准年表(STD)统计特征Tab.1 Statistics of tree-ring width standard chronology (STD) of P. yunnanensis at different altitudes 不同海拔云南松径向生长与月均气温、月降水量和帕尔默干旱指数的关系如图2所示。低海拔云南松径向生长与当年5月平均气温显著负相关(P<0.05),与当年9月平均气温显著正相关(P<0.05),与生长季初期1月、3月和5月降水量显著正相关(P<0.05),与上年生长季初期5月、生长季后期10—12月、当年生长季初期和旺期1—6月帕尔默干旱指数显著正相关(P<0.05)。 中海拔云南松径向生长与当年生长季初期3月、5月平均气温显著负相关(P<0.05),与上年生长季旺期6月降水量显著负相关(P<0.05),与当年生长季初期2、3和5月降水量显著正相关(P<0.05),与上年生长季后期10—12月、当年生长季初期和旺期1—6月及生长季后期10月帕尔默干旱指数显著正相关(P<0.05)(图2)。 高海拔云南松径向生长与上年生长季旺期7月平均气温显著正相关(P<0.05),与上年生长季后期12月降水量显著正相关,与上年生长季旺期7月帕尔默干旱指数显著负相关(P<0.05),与当年生长季初期1、2、3和5月帕尔默干旱指数显著正相关(P<0.05)(图2)。 图2 不同海拔云南松径向生长与区域气候要素相关关系Fig.2 Correlation coefficient between radial growth of P. yunnanensis of different altitudes and climatic factors 各海拔段的云南松树轮宽度标准年表与平均气温、降水量及帕尔默干旱指数的冗余分析(图3)表明,不同海拔云南松径向生长受气候变化的影响具有一致性,在51个气候变量中,上年7月平均气温、当年7月降水量、当年9月平均气温及当年5月帕尔默干旱指数对径向生长的影响达到显著水平(P<0.05),其中与上年7月平均气温、与当年9月平均气温、当年5月帕尔默干旱指数显著正相关(P<0.05),与当年7月降水量显著负相关(P<0.05)。高海拔云南松径向生长受气温影响较大,而中、低海拔云南松径向生长受降水影响更多。 图3 不同海拔云南松树轮宽度标准年表与气候因子的冗余分析Fig.3 Redundancy analysis between tree ring width standard chronologies of different altitudes and the climatic factors 低海拔区云南松树轮宽度标准年表(图4)显示,树轮宽度指数在1980—1987和2009—2014年间明显减小, 1986年树轮宽度指数最低值达0.66, 2012年树轮宽度指数最低值达0.44。图5表明,低海拔云南松径向生长变化百分率在1983—1985年小于-25%, 2009—2011年达-32.7%~-48.0%, 2010年达到最低; 云南松在1978—1990年及2004—2015年发生了生长衰退。 中海拔区云南松树轮宽度标准年表(图4)显示,树轮宽度指数在1877—1890、1900—1907和1988—2012年间呈明显减小趋势,1888年树轮宽度指数最低值达0.49, 1906年最低值达0.69, 2012年最低值达0.65。图5显示,中海拔云南松径向生长变化百分率在1884—1886年小于-25%,在1885年达-30%以上,1897—1900年达到-28.28%~-31.06%,近期在2010年达-28.83%; 云南松在1880—1891、1893—1904和2006—2015年发生了生长衰退。 高海拔区云南松树轮宽度标准年表(图4)显示,树轮宽度指数在1903—1908、1949—1957、1985—1990和2010—2014年呈明显减小趋势, 1907年最低值达0.57, 1955年最低值达0.61, 1989年最低值达0.66, 2014年最低值达到0.82。图5表明,高海拔云南松径向生长变化百分率在1903—1906年小于-25%, 1903年最低值达-42.8%, 1947—1949年均小于-25%, 1949年最低值达-33.9%, 1985—1986年均小于-25%, 2010年为-24.6%(接近于-25%)。 图4 不同海拔云南松树轮宽度标准年表Fig.4 Tree ring width standard chronologies of P. yunnanensisat different altitudes 图5 不同海拔云南松径向生长变化率(箭头指云南松径向生长发生衰退)Fig.5 Percentage change in radial growth of P. yunnanensisat different altitudes(arrows indicate the radial growth decline of P. yunnanensis) 根据帕尔默干旱等级评定标准,最终确定1943、2012和2013年为金沙江流域极端干旱年份(图6)。1943年,中、高海拔区前一年平均树轮宽度指数均在1.44~1.65水平。2012年,各海拔区前一年的平均树轮宽度指数在0.60~0.94水平。2013年,各海拔区前一年的平均树轮宽度指数均在0.43~0.91水平。 图6 极端干旱年份的确定Fig.6 Identification of extreme drought years 不同海拔云南松应对极端干旱的抵抗力和恢复力如图7所示。在1943年,中、高海拔云南松抵抗极端干旱的能力无明显差异,而在干旱之后高海拔区云南松体现出的恢复力要比中海拔区强。在2012年,高海拔区云南松抵抗力最强,其次是中海拔区,低海拔区最弱,而恢复力则相反。在2013年,中海拔区云南松的抵抗力略强于低海拔和高海拔区,而恢复力表现为低海拔区最强,高海拔区略高于中海拔区。 图7 金沙江流域不同海拔云南松应对极端干旱的抵抗力(Rt)和恢复力(Rc)Fig.7 Resistance (Rt) and resilience (Rc) of P.yunnannesis to extreme drought at different altitudes in the Jinsha River Basin 低海拔区云南松应对极端干旱的抵抗力表现出随时间延长而增强的趋势,而恢复力表现出下降趋势; 中海拔区云南松抵抗力随时间延长无明显差异,恢复力在2012年极端事件时有所增强; 高海拔区云南松的抵抗力和恢复力随时间变化稳定。 金沙江流域云南松径向生长的主要影响因素:在低、中海拔区是生长季初期3月和5月的平均气温、生长季初期和旺期的降水量及干旱强度; 在高海拔区是生长季旺期7月的平均气温、上年生长季后期12月的降水量及生长季初期的干旱强度。这说明温度是高海拔云南松生长的限制因子,而水分是中、低海拔云南松生长的主要限制因子。3—5月是云南松形成层分化活跃期,此时的水热条件对形成层分化有关键作用(Begumetal., 2012; Rossietal., 2016),而该时期是研究区的旱季,在降水补充缺乏情况下,高温会产生更强的干旱胁迫,抑制形成层活动。这也是当年5月帕尔默干旱指数与3个海拔段云南松径向生长都显著正相关的原因。生长季旺期的云南松需要充足的水分来保证生长,这段时期虽有充足降水补充,但中、低海拔较高的温度会加速土壤水分蒸散消耗,造成水分供应短缺而影响云南松生理活动(Lévesqueetal., 2014)。7月高温对树木生长的促进作用在其他地区高海拔地段也有类似发现。Panthi等研究发现,夏季温度是影响滇西北白马雪山和石卡雪山长苞冷杉(Abiesgeorgei)径向生长的主要因子(Panthietal., 2018),高海拔区的树木生长对温度具有更强的敏感性(Krausetal., 2016; Heetal., 2017)。随海拔升高,降水量有所增加,气温逐渐下降(Yuetal., 2011),冷湿环境不宜树木生长,这也是高海拔区云南松生长受7月降水量限制的主要原因。 云南松衰退事件可从径向生长历史中判定。研究表明树木发生生长抑制是生长衰退的信号,长时间的生长抑制表明树木发生了生长衰退(Amorosoetal., 2012)。金沙江流域的云南松树轮年表及树木径向生长变化率反映了在1880—1891、1893—1907、1949—1957、1978—1990和2004—2015年发生了不同程度的生长衰退。生长衰退的强度存在海拔差异,相同时段内低海拔区的生长衰退最明显,其次是高海拔区,中海拔区较弱。结合研究区域云南松径向生长与气象因子的关系(图2、3)发现,不同海拔的云南松径向生长都与帕尔默干旱指数显著正相关,尤其是当年5月;明显的森林衰退均发生在极端干旱事件年之后,时间衔接。由此推测,长期干旱是诱发树木径向生长下降进而引发生长衰退的主要原因。关于青藏高原森林衰退的研究也发现近期的森林衰退与气候变暖导致的干旱频率增强有关 (Fangetal., 2018)。在干旱胁迫下,树木会关闭叶片气孔以防止水分散失,光合作用会减弱,干旱期间伴随的高温会使蒸腾和呼吸作用加强,固碳作用受抑制(Adamsetal., 2013; Panthietal., 2017)。全球很多地区干旱引发的树木生长衰退案例均与水分利用有关(Mcdowelletal., 2016; Mooreetal., 2016; Youngetal., 2016),长期处于水分亏缺环境下的树木在受到更强胁迫时容易出现较高死亡率,导致森林衰退加剧(Liangetal., 2006; 2015; Liuetal., 2013)。极端干旱条件下,更加温暖环境下的树木会因干旱胁迫作用加剧而比常规环境下的树木提前死亡(Adamsetal., 2009); 气候温暖或干燥区生长的树木更接近于森林群落的绝对气候界限,更易受到可利用水分限制,当干旱发生时也更易受到影响(Fieldetal., 1983)。金沙江流域低海拔区云南松生长环境中的温度和干燥程度均高于中、高海拔区,相关分析表明低海拔区云南松生长易受生长季初期温度和降水的影响,而该时期恰是高温少雨阶段,极端干旱发生时可利用水分较少,树木生理活动受阻严重。研究表明,干旱环境下树木会因发生碳饥饿和水力衰竭而死亡(Allenetal., 2010; Hartmann, 2015),引发森林衰退。这可能也是低海拔区域云南松生长衰退较中、高海拔区域严重的原因。 在极端干旱年份,各海拔段云南松树轮宽度指数均呈较低水平,说明极端干旱事件对云南松生长有明显抑制作用。不同海拔的云南松对相同年份的极端干旱有不同的抵抗力和恢复力。中海拔区抵抗力最强,其次是高海拔区,低海拔区最弱; 恢复力则以低海拔区最强,高海拔区次之,中海拔区最弱。表明不同生境下的树木生态弹性不同,较干燥地区的森林对极端干旱的抵抗力较低,更易发生树木死亡和生物量下降(Longoetal., 2018),且树木的抵抗力会随纬度、土壤湿度和植被指数的增加而增强(Gazoletal., 2016)。森林的恢复力主要取决于树木生长趋势及森林对死亡损失的补充能力(Fangetal., 2018)。低海拔区云南松长期处于较干旱环境,对干旱的适应能力强于中高海拔区,遇到严重干旱事件干扰后能及时修复极端干旱影响(Xuetal., 2016)。高海拔区林线附近的森林林木稀疏,在极端事件干扰后恢复力较弱,并有可能会被林线附近的草地或灌丛替代,因此一旦受到极端事件干扰就难以恢复到原始状态(Schefferetal., 2012; Levineetal., 2016)。 本研究时间尺度上,高海拔区云南松的抵抗力和恢复力稳定, 低海拔区云南松的抵抗力增强,但恢复力下降, 中海拔区云南松的抵抗力稳定,恢复力在2012年极端干旱事件后增强。研究发现,极端干旱环境下树木的抵抗力和恢复力之间存在一种平衡关系,抵抗力弱的区域在极端干旱后会体现出较强的恢复力 (Gazoletal., 2016),并且干旱频率的增加及持续时间的延长都会导致树木的抵抗力和恢复力发生改变(Ponceetal., 2013)。 本研究运用树木年轮学方法,建立了金沙江流域不同海拔的云南松树轮宽度年表。在此基础上,通过树轮宽度对气象因子响应的分析表明: 中、低海拔的云南松生长主要受水热条件影响,高温和水分胁迫是主要限制因子; 高海拔的云南松生长主要受生长季初期水热条件及生长季旺期7月温度的限制; 接近森林分布气候界限环境的树木更容易发生生长衰退现象; 用抵抗力和恢复力指标评估云南松的生态弹性发现,水、热条件适宜环境下生长的云南松抵抗极端事件干扰的能力更强,接近生长极限环境的云南松受极端事件干扰后的恢复能力更强。3 结果与分析
3.1 不同海拔云南松树轮宽度标准年表统计特征
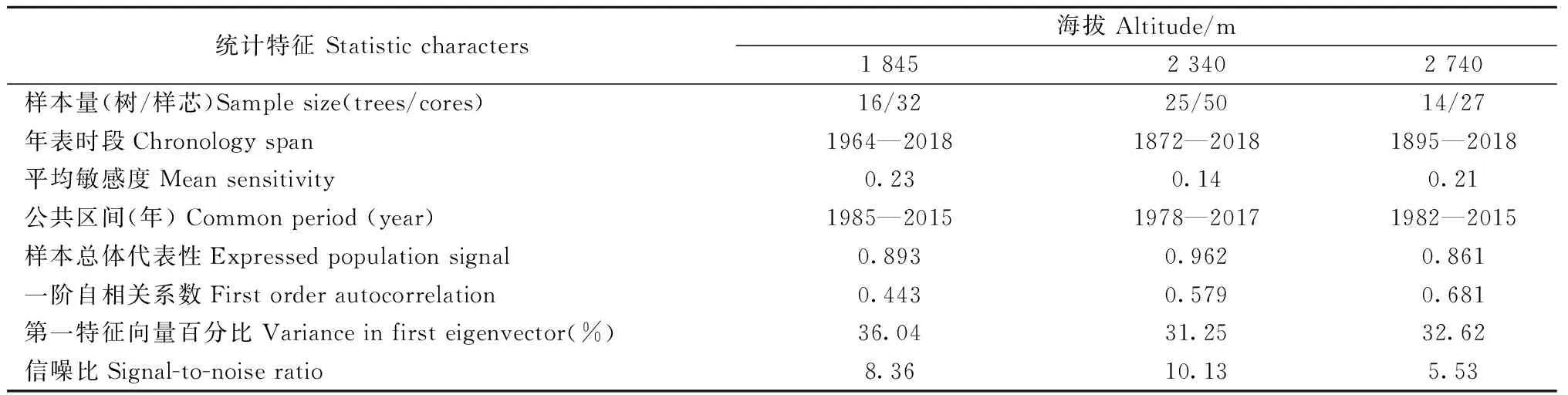
3.2 不同海拔云南松径向生长与气象因子的关系
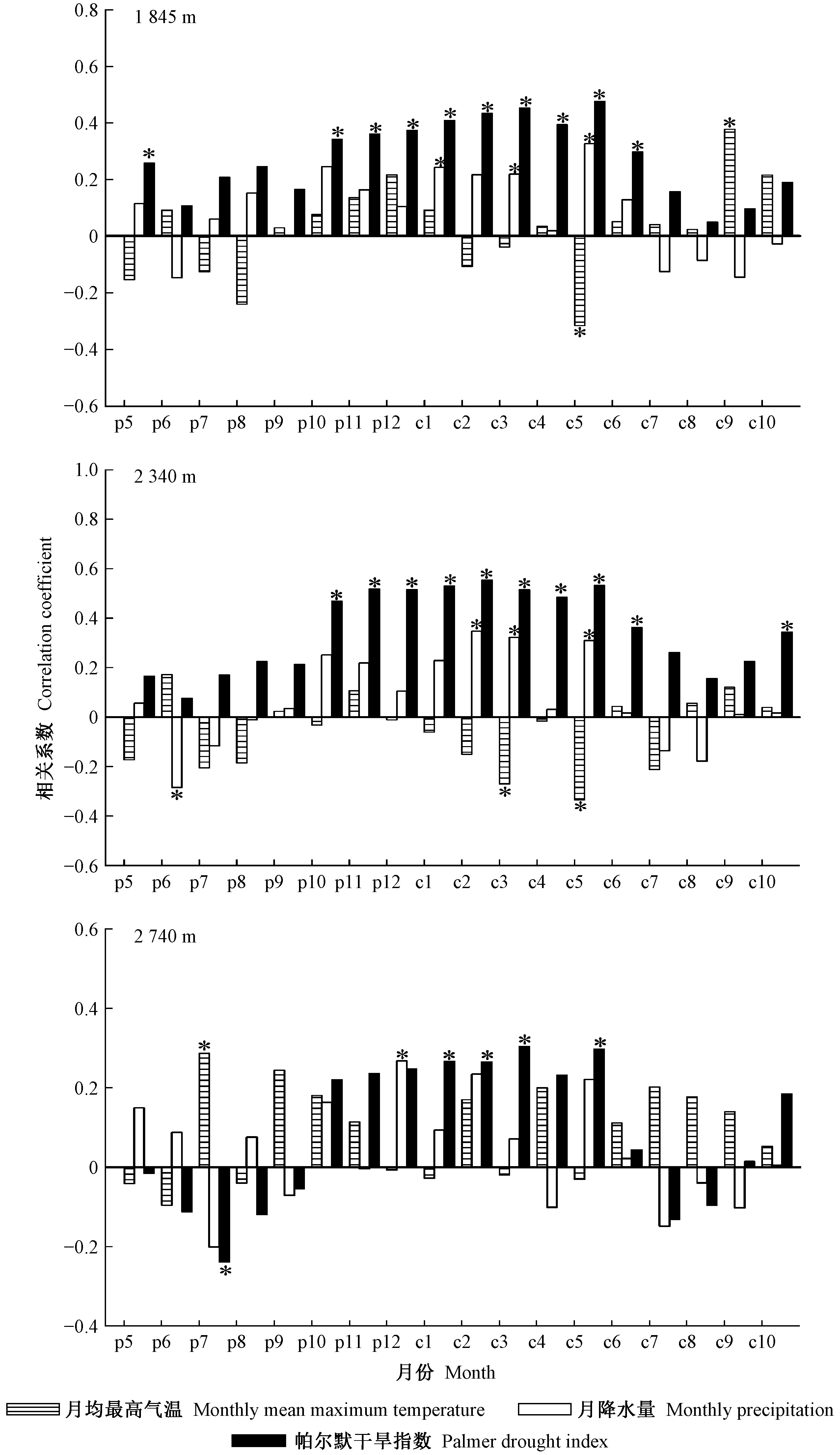
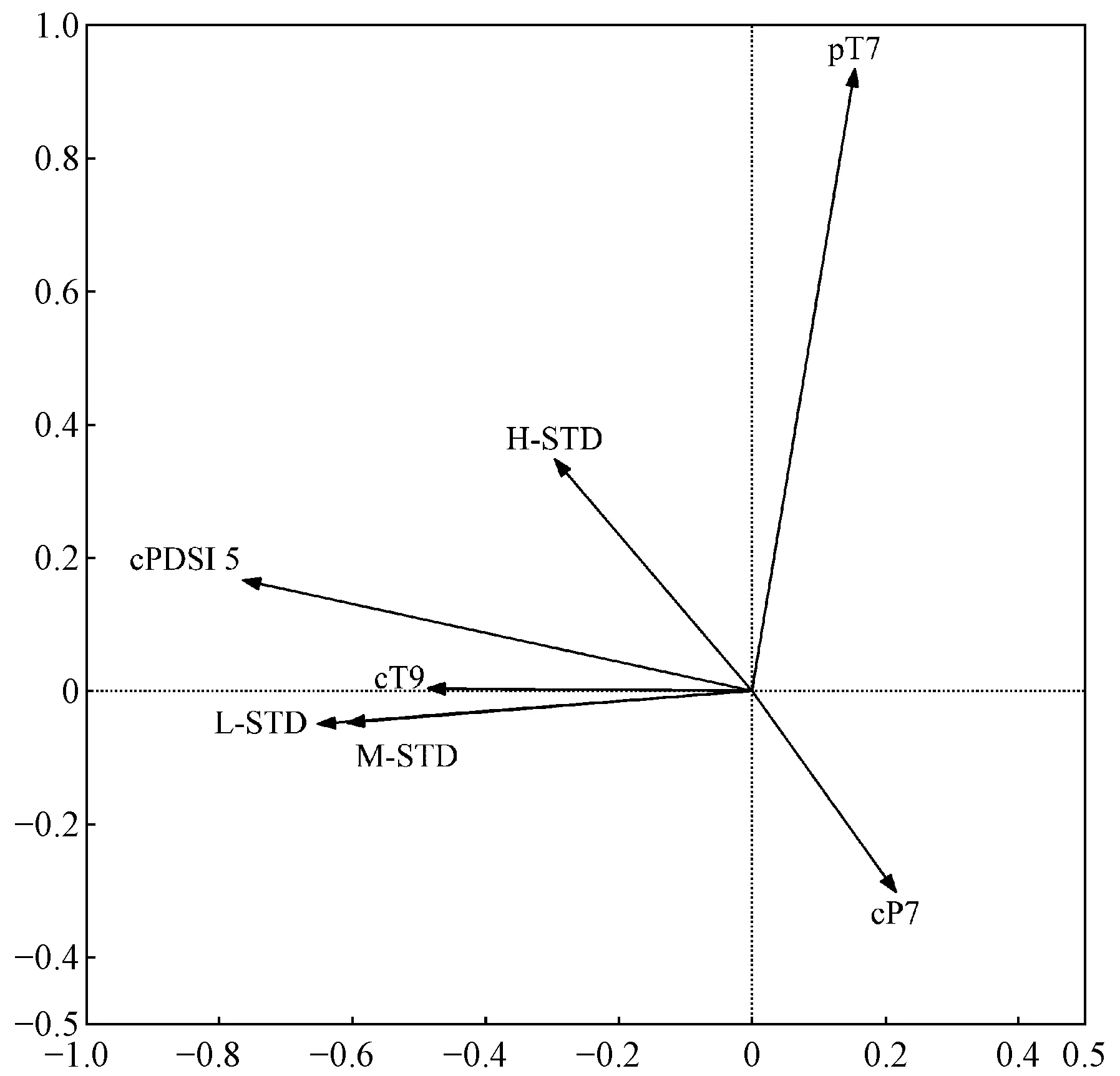
3.3 不同海拔云南松树木生长衰退过程

3.4 不同海拔云南松的抵抗力和恢复力

4 讨论
4.1 影响不同海拔云南松生长的主要气象因子
4.2 不同海拔云南松生长衰退历史及原因
4.3 不同海拔云南松的抵抗力和恢复力
5 结论
