PPARγ基因多态性与中国北方人群散发性帕金森病相关性分析
2020-06-29
(青岛大学附属医院神经内科,山东 青岛 266003)
帕金森病(PD)是一种神经系统退行性疾病,可能是由于黑质纹状体分泌的多巴胺减少导致[1]。过氧化物酶体增殖物激活受体γ(PPARγ)作为甾体类激素受体超家族成员之一,在脑基底节区和一些多巴胺受体的表达部位均有分布[2],这一现象暗示PPARγ和PD可能存在某种关联。PD动物模型实验中,PPARγ激动剂具有保护神经元免受损伤的功能[3]。目前研究发现基因的多态性与PD的易感性密切相关[4],但PPARγ基因多态性与PD相关性的研究较少,且性别、年龄、地域以及种族等因素均会对疾病的易感性产生影响,因此,本研究通过对PD患者年龄、性别的分层分析,探讨PPARγ基因多态性对中国北方人群散发性PD易感性的影响。
1 对象与方法
1.1 研究对象
选取2017—2019年于我院神经内科门诊确诊为PD的患者(PD组)391例, 患者均符合英国PD脑库的诊断标准[5],且为居住于北方地区的汉族人群;同时排除有血管源性帕金森综合征、脑外伤、脑膜脑炎及其他神经系统疾病病史者,排除PD家族病史者;选取同期健康查体者(对照组)391例,对照组纳入和排除标准:年龄、性别与PD组相匹配;居住于北方地区的汉族人群;无PD家族病史。PD组男206例,女185例,平均年龄为(62.42±9.32)岁;对照组男213例,女178例,平均年龄为(61.08±10.16)岁。两组间年龄和性别比较均无统计学差异(P>0.05),具有可比性。同时再按照年龄和性别进行亚组分组,PD组中发病年龄≤50岁者为早发性PD组(B1组),>50岁者为晚发性PD组(B2组);B1组和B2组再分别按照性别分为男B1组(B1-1组)、女B1组(B1-2组)、男B2组(B2-1组)、女B2组(B2-2组);对照组按照同样的分组方法进行分组,分为A1~2组及A1-1~B2-2组。所有研究对象本人或家属均签署知情同意书,并获得我院伦理委员会批准(QYFYWZLL25594)。
1.2 DNA提取及基因型检测
空腹采集两组研究对象的外周静脉血2 mL,置于乙二胺四乙酸抗凝管中,采用北京天根生化科技有限公司DNA提取试剂盒(DP318)提取DNA,产物置于冰箱中-20 ℃保存。本实验设计参考相关文献报道的方法[6-7],应用限制性片段长度多态性聚合酶链式反应(PCR-RFLP)技术检测研究对象的基因分型。PCR总体系为25 μL, 包括基因组DNA2.5 μL, TaqDNA聚合酶12.5 μL,前后的引物各0.5 μL及双蒸水9 μL。PPARγ基因rs1801282位点引物序列为F: 5′-GCCAATTCAAGCCCAGTC-3′,R:5′-GATATGTTTGCAGACAGTGTATCA-GTGAAGGAATCGCTTTCCG-3′。扩增反应条件为:94 ℃预变性5 min;然后94 ℃变性30 s,64 ℃退火30 s,72 ℃延伸30 s,共30个循环;再72 ℃延伸5 min,最后4 ℃终止反应。PCR产物经BstUⅠ(纽英伦生物技术北京有限公司)60 ℃孵育过夜。PPARγ基因rs3856806位点引物序列为F:5′-AGGTTTGCTGAATGTGAAGC-3′,R:5′-GGTGAA-GACTCATGTCTGT-3′。扩增反应条件:94 ℃预变性5 min;然后94 ℃变性30 s,57.3 ℃退火30 s,72 ℃延伸30 s,共30个循环;再72 ℃延伸5 min,最后4 ℃终止反应。PCR产物经BsaAⅠ(纽英伦生物技术北京有限公司)60 ℃过夜孵育。两位点酶切后产物于25 g/L琼脂糖凝胶上电泳,进行成像分析确定基因分型。
1.3 统计学方法
采用SPSS 21.0软件进行统计学分析,以Hardy-Weinberg平衡定律评估所有受试者是否具有代表性,两组间年龄比较采用t检验,性别、基因型和等位基因频率分布在两组间差异采用χ2分析,采用Haploview 4.2软件对PPARγ基因 rs1801282以及rs3856806两个位点的单体型频率及是否连锁不平衡进行分析,以r2>0.33认为两位点存在连锁不平衡,以P<0.05为差异有显著性。
2 结 果
2.1 性别、年龄对PPARγ基因多态性的影响
rs1801282和rs3856806位点的基因型分布均符合Hardy-Weinberg平衡定律(P>0.05)。B1与A1组rs1801282位点等位基因频率比较差异有显著性,携带G等位基因可能会增加早发性PD的易感性(χ2=7.953,P<0.05,OR=11.170,95%CI=1.403~88.911);B1-1组与A1-1组等位基因频率分布比较差异具有统计学意义(χ2=7.630,P<0.05,OR=1.123,95%CI=1.030~1.223)。见表1。B1组与A1组、B1-1组与A1-1组、B1-2组与A1-2组、B2组与A2组、B2-1组与A2-1组、B2-2组与A2-2组rs3856805多态性位点的基因型和等位基因频率分布比较,差异均无显著性(P>0.05)。见表2。
2.2 PD组和对照组PPARγ基因rs1801282位点和rs3856806位点单体型和连锁不平衡分析
对PPARγ基因rs1801282和rs3856806位点进行单体型和连锁不平衡分析,PPARγ呈现基因多态性,两位点间存在较弱连锁不平衡(D′=0.678,r2=0.087),PD组和对照组间C-C、C-T、G-T、G-C 4种单体型分布相比较差异均无显著统计学意义(P>0.05)。见表3。
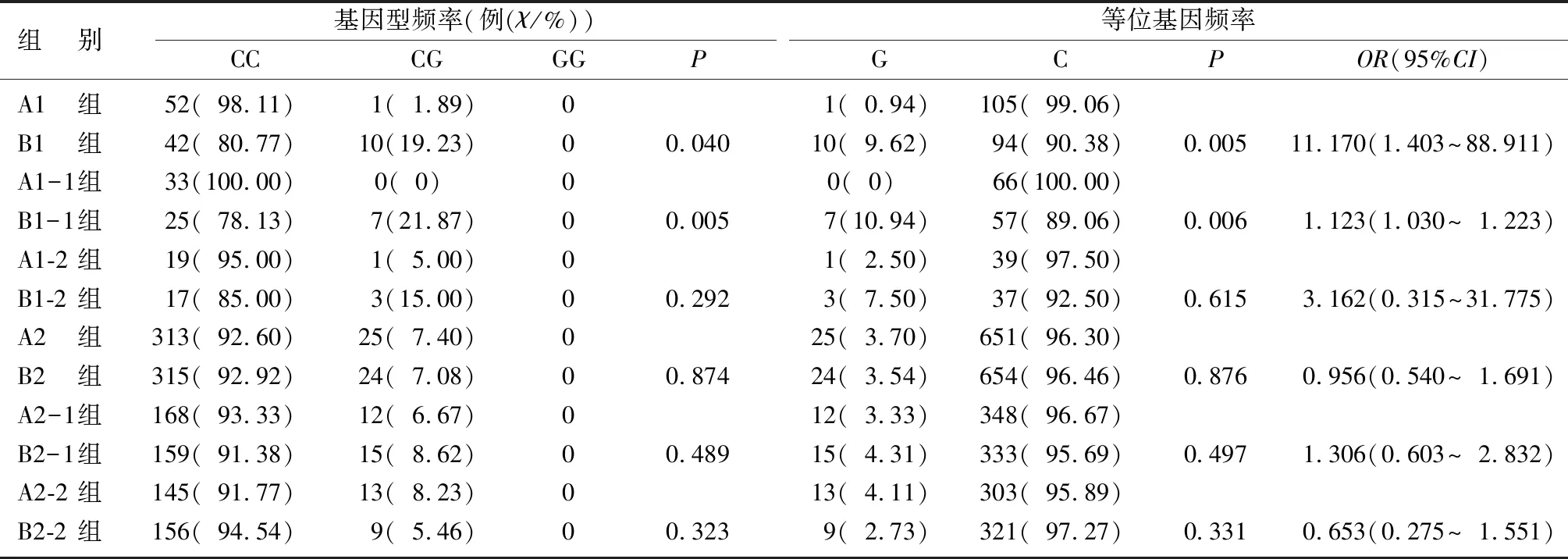
表1 受试者rs1801282基因型频率和等位基因频率分布情况
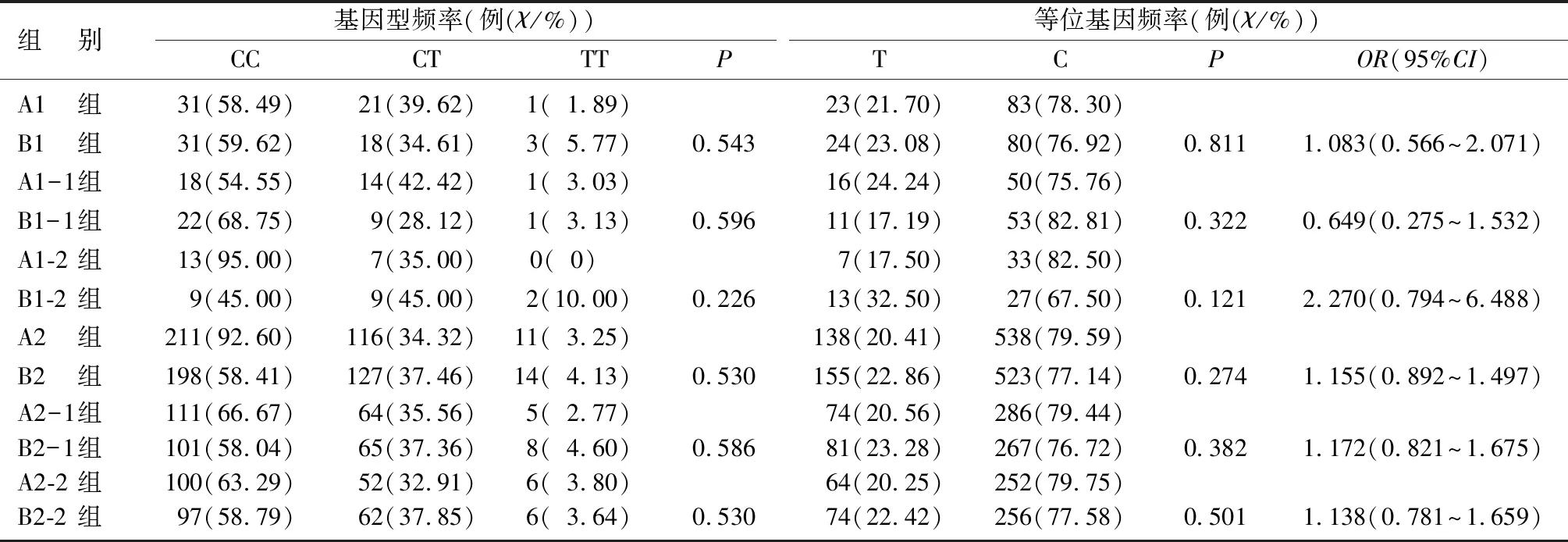
表2 受试者rs3856806基因型频率和等位基因频率分布情况

表3 两组间PPARγ单体型分布比较
3 讨 论
研究显示,PPARγ受体被激活后对PD会发挥保护作用。硫代巴比妥类化合物MDG548作为功能性PPARγ激动剂,在PC12细胞中保护细胞免受H2O2氧化和1-甲基-4-苯基吡啶离子(MPP+)神经毒性,在MPTP处理的小鼠中可减少其黑质中的反应性小胶质细胞以及iNOS生成[8]。在人神经胶质瘤SH-SY5Y细胞中,罗格列酮可改善SOD、Bcl-2以及Bax的表达,保护线粒体免受MPP+引起的损伤[9]。此外,PPARγ激动剂吡格列酮通过抑制6-羟基多巴胺诱导的PD模型大鼠黑质内小胶质细胞增殖及核转录因子NF-κB活化,进而抑制神经系统炎症反应,保护多巴胺能神经元[10]。
PPARγ基因位于染色体3p-25的位置[11],此基因上的多态性位点可能与疾病的易感性有关。文献报道PPARγ基因rs1801282位点多态性可能与神经退行性疾病阿尔茨海默(AD)易感性相关。意大利白种人群中携带G等位基因的80岁及以上老年人患AD的风险是对照组的2倍[12]。在对英国高加索人群的研究中发现,rs1801282位点的G等位基因对女性AD起保护作用,但在男性中作用相反[13],提示这一位点的基因变异可能对男性和女性的作用机制不同。
由于性别、年龄、地域和种族等因素均会对疾病易感性产生影响,因此本研究通过年龄、性别亚组分析,进一步明确不同因素对疾病的影响。在本研究中,对所有研究对象进行年龄、性别亚组分析及单体型分析后,未发现rs3856806位点多态性在各组间
有统计学差异,提示rs3856806位点多态性可能对中国北方人群散发性PD的发生无影响,这与之前对日本及中国南方人群的调查结果一致[14-15]。本研究结果显示,B1组及B1-1组中rs1801282位点G等位基因频率分别高于A1组及A1-1组,表明G等位基因可能是中国北方人群早发性PD及男性早发性PD的危险因素,提示遗传因素可能对PD的发病时间有影响,且早发性PD对男性、女性的致病机制可能不同。国外学者发现rs1801282位点G等位基因可能会使PPARγ基因的转录活性降低[16],据此推测其可能使PPARγ受体表达水平下降,间接导致PPARγ对神经系统的保护作用下降,使PD易感性增加。相关研究证实遗传因素对早发性PD(发病年龄≤50岁)比晚发性PD(发病年龄>50岁)影响更为明显[17],本研究结果也再次验证这一结论。
综上所述,两位点所构建的四种不同单体型与PD无相关性,且两位点只存在较弱的连锁不平衡,本研究首次揭示了PPARγ基因rs1801282位点等位基因G可能会增加中国北方人群早发性PD及男性早发性PD的患病风险。因此,本文研究可能会为未来PD的基因预测以及治疗提供数据的支持。PPARγ基因rs1801282位点与PD易感性相关的作用机制仍需进一步探究。
