基于乳酸-LDH的牦牛肉NADH线粒体介导再生研究
2020-06-29张玉斌李丙子魏红艳余群力
张玉斌 李丙子 雷 芸 魏红艳 韩 鋆 余群力
(1.甘肃农业大学食品科学与工程学院, 兰州 730070; 2.兰州文理学院化工学院, 兰州 730000)
0 引言
牦牛独特的高海拔生长环境和较长的饲养周期使屠宰后的肉色表现出比普通牛肉更暗、消费者更不易接受的深红色,且在冷藏成熟过程中往往伴随着肉色劣变[1-2]。牛肉冷藏和零售过程中的色泽稳定性至关重要[3]。有研究表明,乳酸盐注射增强技术可有效控制有氧贮藏期间冷却肉的褪色[4-5]。乳酸-LDH体系会促进烟酰胺腺嘌呤二核苷酸(NADH)的再生,从而使MetMb的还原能力提高,改善货架期肉色[6],但具体还原机理尚不明晰。因此,有必要基于乳酸-LDH体系对牦牛肉色泽稳定性进行系统研究。
有研究证实,屠宰后的肌肉是有生物化学活性的,并且其有能力通过向被氧化的三价铁肌红蛋白(Mb)增加一个电子来还原高铁肌红蛋白(MetMb)。参与MetMb还原的电子来自NADH或线粒体介导的电子转移[7-8]。在添加了特定的底物后,NADH可以通过在宰后肌肉细胞质[9]中的LDH(cLDH)或线粒体中的LDH(mLDH)再生[10],说明NADH的宰后再生对于延缓肉色劣变至关重要。在分离出的牛心肌线粒体中,添加乳酸盐、LDH和NAD(烟酰胺腺嘌呤二核苷酸氧化态)后产生NADH,这可能是导致耗氧量同时增加的原因[11],NADH可以进入复合物I(NADH脱氢酶)并导致电子传递链中的电子运动。但目前尚无法确定NADH通过乳酸盐-LDH-NAD途径形成后能否被骨骼肌线粒体用于MetMb还原。文献[12]认为,肉类劣变与肌肉线粒体酶活性的关系比与线粒体含量的关系更大,利用TCA(三羧酸循环)循环酶(脱氢酶类)的底物,有助于在宰后骨骼肌中再生还原NADH。文献[13]研究发现,大鼠骨骼肌和心肌中分离出来的线粒体具有不同的蛋白质结构,可以适应每个组织的特定功能。这些研究表明,线粒体在肉色稳定中起着重要的作用。
目前,关于线粒体电子传递链(ETC)与高铁肌红蛋白还原和肉色稳定性的体外孵化研究几乎没有得出确定的结论。此外,由线粒体内三羧酸循环中间代谢物诱导引起的NADH再生过程,通过区分如苹果酸脱氢酶(MDH)和LDH等酶类,进而可以调节MetMb还原、影响Mb氧化还原稳定性,这方面的报道也很少。在线粒体和细胞质之间,由MDH或LDH活性造成的NADH再生在肉色稳定性方面可能起到重要作用。因此,本文假设在LDH中产生的NADH具有改善Mb氧化还原稳定性的潜力,将分离提取出的线粒体和牦牛背最长肌Mb作为研究对象,探究乳酸-LDH模型中乳酸盐对牦牛肉色稳定性的影响,以及肌肉线粒体在ETC中NADH再生和MetMb还原能力,并对体外模型的研究结果进行验证,以期为延缓牦牛肉色泽劣变、保持冷藏期间肉色稳定性提供理论依据。
1 材料与方法
1.1 样品采集与预处理
试验牦牛肉样品由甘肃天玛生态食品科技股份有限公司提供。选取36~48月龄、生长发育良好、健康无病的甘南牦牛6头,屠宰后立即对胴体进行热蒸汽减菌处理,取左胴体背最长肌,剔除表面结缔组织,避光、真空包装后置于冰盒运送回实验室,将运回实验室的样品分为两份,第1份分成4份,在0~4℃条件下分别冷藏1、3、5、7 d,用于测定线粒体膜通透性、膜电位;第2份在24 h内进行肌红蛋白的分离纯化,用于体外孵化试验。
1.2 设备与试剂
TGL-24M型台式高速冷冻离心机(长沙平凡仪器仪表有限公司)、JY92-IIDN型超声波细胞粉碎仪(宁波新芝生物科技股份有限公司)、SP-756P型紫外可见分光光度计(上海光谱仪器有限公司)、AL204型电子天平(梅特勒-托利多仪器有限公司)、抗霉素A(南京森贝伽生物科技有限公司)、K3[Fe(CN)6](上海源叶生物科技有限公司)、NAD(上海源叶生物科技有限公司)、NADH(上海源叶生物科技有限公司)、草氨酸钠(上海源叶生物科技有限公司)、乳酸脱氢酶(美国Sigma公司)。
1.3 试验方法
1.3.1线粒体提取
参照文献[14]的方法提取线粒体。切取适量肉样,用冷藏(4℃)生理盐水洗去表面残血,滤纸吸干后迅速切碎,称取10 g切碎样品,加入100 mL分离液(250 mmol/L蔗糖、10 mmol/L Tris-HCl、1 mmol/L EDTA(乙二胺四乙酸),pH值 7.4),在12 000 r/min转速下匀浆2 min,然后1 500g、4℃离心 15 min,上清液再次在12 000g、4℃条件下离心20 min,沉淀用分离液清洗两次后溶于缓冲液(250 mmol/L蔗糖、10 mmol/L Tris-HCl,pH值7.4)即为线粒体悬浮液(詹纳斯绿B染色鉴定线粒体)。全程低温操作,以上过程均在0~4℃条件下完成。
1.3.2线粒体膜通透性
参照文献[15]的方法测定线粒体膜通透性,测定已制备好的线粒体悬浮液在520 nm波长下的吸光度,依据吸光度的变化来判定线粒体膜通透性的改变。如果溶液在520 nm 处吸光度A520降低,表明线粒体膜通透性增加,反之则表明线粒体膜通透性降低。
1.3.3线粒体膜电位
采用试剂盒测定。线粒体提取试剂盒来自上海索莱宝科技有限公司,线粒体膜电位检测试剂盒(以JC-1为荧光探针)来自北京泛博生物化学有限公司。
线粒体提取试剂盒用于从动物组织中分离出完整而纯化的线粒体。肉样通过冰浴研磨后,添加试剂盒所提供的试剂进行几次离心沉淀后得到重悬线粒体沉淀,立即使用或-70℃保存。荧光探针JC-1聚集在线粒体基质中形成聚合物,产生红色荧光;膜电位较低时,JC-1为单体,产生绿色荧光。通过JC-1从红色荧光到绿色荧光的转变可以很容易地检测到细胞膜电位的下降。将0.9 mL 5倍稀释的JC-1染色工作液加入0.1 mL纯化的线粒体中,孵育20 min,立即用荧光分光光度计检测。加测JC-1单体时激发光波长设置为490 nm,发射光波长设置为530 nm;检测JC-1聚合物时激发光波长设置为525 nm,发射光波长设置为590 nm。
1.3.4牦牛背最长肌Mb的分离纯化
参照文献[16]的方法稍加改进。取切碎后的肉样100 g,在4℃预冷的300 mL提取液(含1 mmol/L EDTA、10 mmol/L pH值 8.0 Tris-HCl、25 g/L Triton X-100(聚乙二醇辛基苯基醚))中混合,在低温高速冷冻离心机中13 000 r/min均质30 s,取出匀浆液,将其在9 500g、4℃下离心10 min,取上清液,用滤纸过滤后得到Mb粗提液。采用硫酸铵溶液对粗提液进行分级沉淀,饱和度梯度由65%至95%,然后4℃条件下3 000g离心10 min后弃去上清液,将沉淀物在5 mmol/L pH值8.5的Tris-HCl缓冲液中溶解。用提前预冷至4℃的相同缓冲液透析10 h,透析液每隔1 h换一次,透析结束后在4℃下5 000g离心10 min,得到Mb初级纯化液。接下来的精细纯化采用Sephadex G-100型凝胶层析分离柱进行。收集540 nm波长下吸光度较高的蛋白溶液,参照文献[17]的方法对分离得到的Mb进行光谱特性和SDS-PAGE(十二烷基硫酸钠-聚丙烯酰胺凝胶电泳)鉴定,如表1所示。
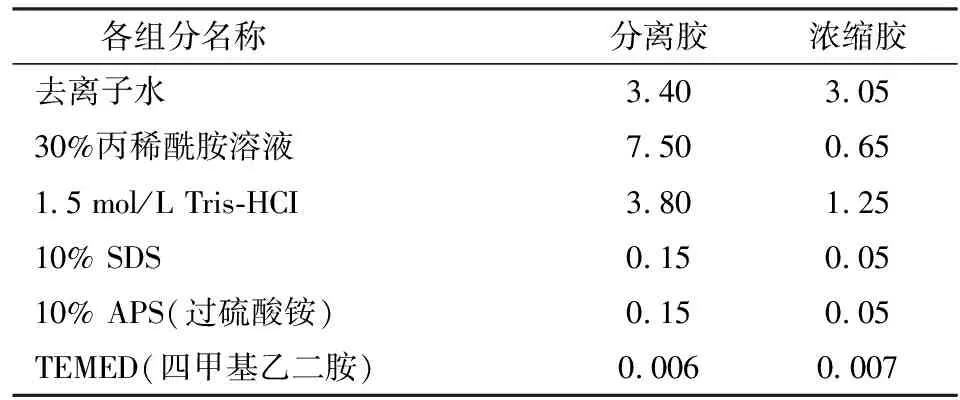
表1 SDS-PAGE电泳凝胶各组分质量分数Tab.1 Formula of SDS-PAGE gel %
1.3.5牦牛背最长肌MetMb制备
参照文献[16]的方法稍作调整。取1.3.4节中分离纯化的Mb溶液5 mL,添加K3[Fe(CN)6]50 mg反应10 min即得到MetMb溶液。上述反应结束后,溶液在3 500g、4℃条件下离心10 min,上清液中多余的Na2S2O4和K3[Fe(CN)6]采用提前预冷至4℃的50 mmol/L、pH 值7.0的Tris-HCl缓冲液透析除去。MetMb溶液必须现用现配,并用Tris-HCl缓冲液按照试验要求调整溶液中蛋白的浓度。
1.3.6线粒体对MetMb还原的影响
参照文献[18]的方法在340 nm处测定吸光度,提取的牦牛背最长肌线粒体(3 mg/mL)与处理好的MetMb(2.5 mg/mL)结合的基础上添加NAD、乳酸钙、乳酸钙-NAD、乳酸钙-LDH-NAD或乳酸钙-LDH-NAD(在加入乳酸钙-LDH-NAD体系之前,线粒体首先和抗霉素A预孵化10 min)。对照组样品只由线粒体(MT)和MetMb组成,不添加乳酸钙或乳酸-LDH-NAD。反应组分的浓度为乳酸钙40 mmol/L,LDH(100活性单位)40 mmol/L,NAD 0.2 mmol/L,所有样品都装在带盖密封的旋口瓶中,以免外部氧气进入线粒体-高铁肌红蛋白反应混合物。各处理组具体情况如表2所示。

表2 添加到牦牛背最长肌线粒体(MT)和MetMb中的不同组合Tab.2 Treatment combinations added to bovine skeletal mitochondria (MT) and myoglobin
注:LLN代表CaL+LDH+NAD;AA代表抗霉素A;SO代表草氨酸钠;“+”表示底物/抑制剂存在,“-”表示不存在。
MetMb(2.5 mg/mL)还原是在pH值 5.6的缓冲液(120 mmol/L KCl,30 mmol/L KH2PO4,30 mmol/L顺丁烯二酸)中使用石英比色皿持续3 min,然后样品在4℃下孵育10 h。在孵育期间的特定时间点(0、4、8、12 h),取出样品并用离心机以10 800 r/min离心5 min,得到的上清液用分光光度计在500~650 nm波长范围内进行扫描,采用文献[19]的方法计算氧合肌红蛋白和高铁肌红蛋白的含量。
1.3.7氧消耗速率的测定
参照文献[20]的方法并略作修改。样品耗氧率利用Clark型氧电极进行测量。在 25℃下校正Oxytherm型液相氧电极,用连二亚硫酸钠(高浓度)校正零氧基线,现用现配,然后用去离子水清洗反应室6次以上。再用反应缓冲液冲洗几次,向反应室加入2 mL氧饱和的缓冲液,盖好反应室盖子(保持密闭,以免外界氧气进入反应室)。开启转子,开始记录氧含量,等待2 min使氧含量平稳。取提取的线粒体及反应物质加入反应室,立即计时,记录5 min内的呼吸速率,即样品的OCR值。测量温度分别保持在4℃以及25℃,即得到肌肉组织在两种不同温度下的氧消耗速率。对照样品只有线粒体和高铁肌红蛋白,没有添加底物或抗霉素A。为了评价复合物Ⅲ(泛醌+细胞色素c氧化还原酶)抑制剂对线粒体呼吸作用的效果,线粒体和抗霉素A提前预孵化2 min,其他没有添加抗霉素A的处理组,添加等量的乙醇,其次添加底物,计算氧消耗速率。
1.4 数据处理分析
采用SPSS 19.0统计分析软件(美国IBM公司)进行试验数据处理,显著性差异(P<0.05)通过LSD法进行比较分析。采用Origin 8.0软件进行图形绘制。试验中所有测定做3次重复。
2 结果与分析
2.1 乳酸钙处理对线粒体膜通透性的影响
图1(图中不同字母表示同种处理冷藏时间内差异显著(P<0.05),下同)为线粒体膜通透性的变化趋势,从图1可以看出,随着冷藏时间的延长,线粒体膜通透性呈现下降的趋势,且1~3 d下降缓慢,从第5天开始下降速度加快,冷藏结束与开始具有极显著的差异性(P<0.05)。线粒体结构损伤的表现之一就是膜通透性的变化,双层膜通透性转换孔增大,膜的流动性改变,导致其失去对线粒体内部与细胞液中物质交换的调控能力。在520 nm波长下吸光度的下降表明线粒体发生肿胀,同时,线粒体内部小分子物质向外释放的过程引起了渗透压的变化,从而升高基质中蛋白质浓度,促使线粒体大面积发生肿胀。线粒体渗透转换孔(MPTP)的开启能够引起线粒体膜通透性的改变,因此通过测定线粒体膜通透性能有效反映线粒体膜的损伤情况。膜通透性增大可直接导致其内部的生理环境急剧变化,进而造成其生理功能损伤,最终可能对其还原MetMb的能力造成一定的影响。
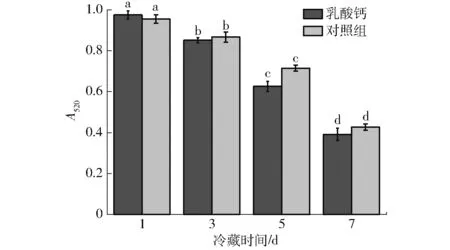
图1 成熟过程中牛肉线粒体膜通透性的变化Fig.1 Changes in mitochondria membrane permeability of beef during aging
2.2 乳酸钙处理对线粒体膜电位的影响
由图2可知,在冷藏期间线粒体的膜电位呈现出逐渐下降趋势,1~3 d下降缓慢,5~7 d下降较快。乳酸钙处理与对照组在第7天时线粒体膜电位分别为0.59 mV和0.47 mV,与第1天相比分别下降了52.03%和60.50%。在冷藏结束时线粒体膜电位降到最低,这与线粒体膜通透性的变化趋势相似,表明MPTP的大量开启能引起线粒体膜通透性的改变,从而造成线粒体内分子质量低于1.5 ku的溶质通透性突剧增,使得膜电位下降瓦解。
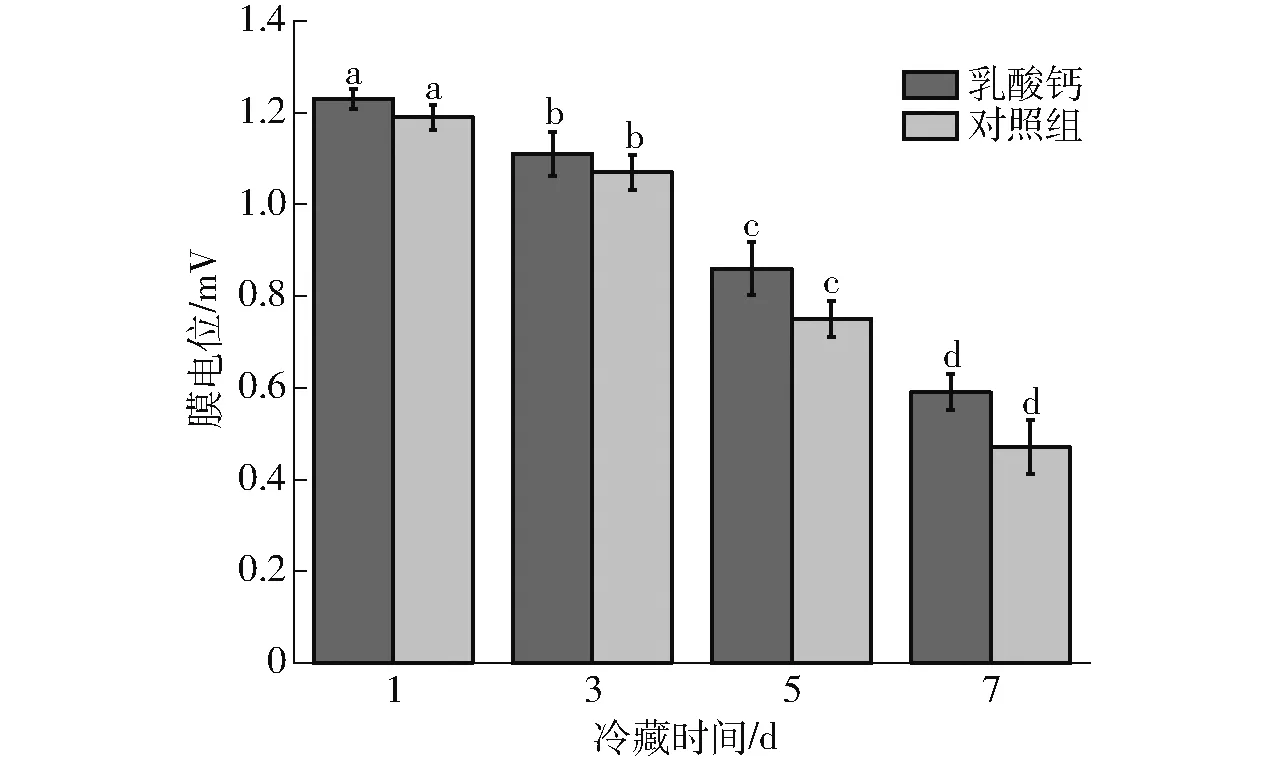
图2 成熟过程中乳酸钙处理对牛肉线粒体膜电位的影响Fig.2 Effects on mitochondrial membrane potential activity of beef during aging
2.3 体外乳酸盐对氧消耗速率的影响
在典型的肉品指标pH值5.6、4℃条件下,与没有添加底物的线粒体组相比,添加了乳酸钙的处理组氧消耗速率有所增加(P<0.05),而LLN组的样品氧消耗速率在所有的试验组中是最高的(P<0.05)。这也与之前的一些研究报道的结果一致,即在生理指标pH值7.4条件下,利用从老鼠骨骼肌和牛心肌中分离出来的线粒体,有能力使用乳酸作为氧气消耗的底物,由LDH活性形成的NADH可以被用于电子传递和还原酶介导的MetMb还原[21]。
2.4 体外乳酸盐对NADH形成的影响
没有添加底物的线粒体对照组和添加了乳酸钙的处理组并没有导致NADH的生成,但CaL-LDH-NAD组合的处理组340 nm处的吸光度增加,这说明NADH的生成能够被电子传递链的复合物Ⅰ所利用。抗霉素A处理组在340 nm处的吸光度比对照组的显著增加(P<0.05),这说明抗霉素A并没有明显阻止NADH的生成,这与本研究中抗霉素A处理后耗氧量显著降低的变化情况不一致,可能是由于乳酸-LDH-NAD形成的NADH被用于酶促的高铁肌红蛋白还原,而不是线粒体氧消耗,因为抗霉素A抑制了电子传递链的复合物Ⅲ[22]。
2.5 体外乳酸盐对线粒体-高铁肌红蛋白还原的影响

图3 乳酸-LDH-NAD体系体外孵化模型中线粒体介导的电子传递对MetMb还原的影响Fig.3 Effects of lactate-LDH-NAD on electron transport linked mitochondria-mediated metmyoglobin reduction in vitro
由图3可以看出,当不添加乳酸盐等底物时,体系内的Mb氧化还原状态几乎未发生改变(P>0.05)。这说明由LDH形成的NADH不能在没有还原酶或电子载体的情况下还原MetMb[23],与对照组相比,添加乳酸钙到MetMb和线粒体的组合中后对MetMb的还原也有一定的影响(P<0.05)。抗霉素A组显著降低了与CaL-LDH-NAD相关的MetMb还原(P<0.05),和其它所有处理组相比,添加到分离的线粒体中的CaL+LDH+NAD体系引起的MetMb还原在所有的试验组中是最高的(P<0.05)。说明当宰后NADH再生时,由CaL-LDH相互作用产生的NADH可以用于ETC介导的非酶促或酶促反应下的MetMb还原。
3 讨论
线粒体膜通透性的变化是用来衡量线粒体结构变化以及功能损伤的一个重要指标[24],主要以提取线粒体悬浮液在520 nm波长下的吸光度变化来判断线粒体膜通透性的变化。线粒体膜通透性改变是线粒体结构损伤的标志,会引起膜流动性、跨膜电位、膜蛋白构象变化等现象,而线粒体中电子传递、氧化磷酸化等重要的生化反应均发生在线粒体膜中,线粒体膜的损伤势必导致线粒体的生理功能衰退[25-26]。具体表现为线粒体膜通透性的增大、线粒体形状、大小改变以及线粒体数目减少,导致的生理损伤主要表现为线粒体标志酶活性的丧失、代谢产物浓度降低和氧消耗速率的减小[27]。文献[8]研究了用琥珀酸盐作为底物的电子传递链介导的MetMb还原。将琥珀酸盐添加到分离提取出的线粒体后,通过复合物Ⅱ(琥珀酸脱氢酶)进入电子传递链,从而使MetMb还原。在添加琥珀酸后,线粒体介导的MetMb还原在宰后45 d内仍然保持活跃。这一观点支持了琥珀酸盐和乳酸盐等底物在电子传递链介导的MetMb还原过程中的作用。在本研究中,添加乳酸钙或者CaL+LDH+NAD组合体系都不同程度地增加了线粒体介导的氧消耗速率和MetMb还原。
文献[28]研究发现,电子传递链介导的MetMb还原发生在较低的氧分压下,这是由氧消耗增加所致。本研究中,与没有添加底物的线粒体组相比,添加了乳酸钙的处理组氧消耗速率有所增加(P<0.05),和线粒体结合的LLN组的样品氧消耗速率在所有的试验组中是最高的(P<0.05)。CaL-LDH-NAD体系增加了耗氧量,创造了一个氧分压以支持电子传递链中电子的可用性。这些电子能够通过在线粒体中存在的电子载体来减少Mb的氧化。降低分压有助于MetMb还原。文献[29]的研究结果表明MetMb的还原是在NADH直接添加到线粒体后。文献[30]报道了在绞碎的羊肉半膜肌直接添加NADH可以增加耗氧量,宰后肉的耗氧量取决于NADH的水平。因此,CaL+LDH+NAD系统增加MetMb还原和氧消耗的能力可能是由于NADH的形成,这一点在本研究中通过在340 nm处的吸光度增加也得到了证实。
有些特异性的抑制剂能够使MetMb还原更有效的电子位点,相关研究发现异戊巴比妥、普罗黄素、抗霉素A、阿霉素、双香豆素等的抑制位点[31]。抗霉素A对几乎所有生物的线粒体呼吸链都有抑制作用,通过作用于细胞色素c将有限的电子传递给三价铁Mb[32]。在乳酸盐的氧化过程中,由ETC产生的电子是导致MetMb还原的原因。Mb作为一种肌浆蛋白,其分子过大不能穿过线粒体外膜,因此,需要一个电子传递介质将电子从ETC转移到MetMb。细胞色素c是一种能将电子从线粒体复合物Ⅲ转移到复合物Ⅳ(细胞色素c氧化酶),并可释放到膜间隙中的功能蛋白质[33]。细胞色素c被认为是在胞质NADH的有氧氧化过程中在线粒体内、外膜之间的电子穿梭体[34]。本研究发现,预孵育之后的线粒体与抗霉素A组显著降低了与CaL-LDH-NAD相关的耗氧量,同时显著降低了与CaL-LDH-NAD相关的MetMb还原,说明抗霉素A能特异性地阻断电子传递链的复合物Ⅲ,阻止电子从细胞色素b传递到细胞色素c,只能通过细胞色素c传递有限的电子给三价铁Mb,抑制了MetMb还原。至于牦牛肉在冷藏过程中体内的MetMb还原是否也是这一途径还需要进一步研究。
4 结论
(1)抗霉素A能特异性地阻断线粒体电子传递链的复合物Ⅲ,阻止电子从细胞色素b传递到细胞色素c,显著降低与CaL-LDH-NAD相关的耗氧量及MetMb还原能力,但并未明显阻止NADH的生成。NADH可以通过充当电子传递链的复合物Ⅰ的底物,进而启动氧消耗,CaL-LDH-NAD体系增加了NADH依赖的还原酶活性。
(2)添加LDH抑制剂草氨酸钠可以降低CaL-LDH-NAD体系对MetMb的还原,但却并未完全抑制MetMb还原,说明除了电子传递链介导的非酶促还原外,CaL-LDH-NAD体系产生的NADH也可用于线粒体内的酶促MetMb还原。
(3)由乳酸-乳酸脱氢酶体系产生的NADH将导致线粒体氧消耗和MetMb向氧合肌红蛋白的转化。因此,当乳酸盐(CaL)底物存在时,线粒体有能力再生出相应的还原物,这些还原能力可以影响氧消耗、MetMb还原以及Mb氧化还原状态的转变,进而起到稳定牦牛肉色泽的作用。
