基于宏基因组的青海农用沼气池微生物组成和功能分析
2020-06-29陈来生李全辉钟启文朱德锐
韩 睿 陈来生 李全辉 李 屹 钟启文 朱德锐
(1.青海大学农林科学院青海省蔬菜遗传与生理重点实验室, 西宁 810016;2.青海大学医学院基础医学研究中心, 西宁 810016)
0 引言
发展沼气是一种极具应用前景、可解决农村能源供应问题和环保问题,以及实现农业可持续发展的有效途径[1]。温度是制约沼气发酵效率的关键因素之一[2-3]。青海省自2003年实施农村沼气能源建设项目以来,在开发和建设方面取得了一定成效,但受温度的影响,绝大多数沼气池存在产气量少、产气率低、运行状况差和利用效率低等问题,严重影响了沼气的推广普及[4]。为解决此问题,沼气用户曾采用覆盖塑料薄膜、畜禽舍下建造沼气池及池体进出口加盖等措施给沼气池升温和保温,但仍不能增加产气量和提高利用率,无法满足农户基本需求,亟需开发出针对沼气发酵工艺的新技术。高效稳定的微生物生态系统是沼气池稳定运行和提高沼气产气率的保证。文献[5]研究显示,小容量沼气池中不同底物对微生物群落分布变化及甲烷产生具有影响;文献[6]对沼气生产过程中微生物群落的原位活动及多样性进行了测定,发现Thermotogae和Spirochaetes菌群有活性糖基代谢功能,Fusobacteria和Cloacimonetes菌群有活性氨基酸基代谢功能;文献[7]对生物沼气池的样品进行高通量测序,结果表明,产甲烷的代谢途径中多含有酸化酶、水解酶、醋酸化酶和产甲烷酶的微生物菌群,认为后期生产中可以通过对微生物群落的监控实现优化生产。因此,深入解析沼气发酵过程中微生物的群落组成以及群落的功能、发掘新的重要微生物功能类群、优化群落结构并调节应用,是解决青海沼气池产气量少、产气率低、运行状况差和利用效率低等问题的有效途径。
绝大多数沼气发酵微生物难以培养,无法采用传统纯培养方法研究微生物群落结构和代谢关系。宏基因组技术是以环境中所有微生物基因组DNA为研究对象,利用分子与信息技术系统研究微生物多样性、群落结构以及微生物-环境的互作关系[8]。该技术无需进行繁琐的常规分子生物学实验,有效降低了实验偏差。同时,从微生物群落结构水平上,宏基因组技术可全面认识微生物的生态特征和功能,尤其是涉及各种复杂环境的微生物群落的鉴定[9-14]。迄今为止,针对高海拔、高寒地区农用沼气池微生物群落结构和功能的研究报道较少,尚缺乏足够的理论参考依据。本文借助Illumina HiSeq 3000高通量测序技术,获得青海省典型农用沼气池污泥样品的宏基因组数据,对微生物的群落结构和潜在的功能类群进行系统分析,旨在为后续改进青海沼气发酵工艺、提高沼气利用效率和实现沼气发酵的稳定运行提供一定的理论依据和技术参考。
1 材料与方法
1.1 农用沼气池
沼气池选自青海省乐都县蒲台乡李家台村(36°21′14″N,102°27′22″E)。该村为青海省沼气池普及和推广效果较好的地区之一,池容积为8 m3,采用“一池三改”的模式。选取投料、进出料状况及管护情况基本一致,且运行稳定的2户沼气池为研究对象。其中一户沼气池产气好,全年产气不中断,保证农户的三餐需求,为青海极少数沼气池的典型代表(记为LD-G);另一户沼气池产气差,全年仅5—10月使用(11月至翌年4月基本无法使用),为青海绝大多数的沼气池代表之一(记为LD-B)。2户投料均为:质量分数80%~90%猪粪+质量分数10%~20%人粪。
1.2 样品采集
前期研究表明,青海农用沼气池产气量与发酵温度呈正相关[4,15]。分别采集上述2户全年温度最高时期(9月上旬,即全年月平均产气量最高)和最低时期(3月中旬,即全年月平均产气量最低)的污泥样品进行分析。编号如下:LD-G温度最高和最低时期的样品分别记为LD-GH和LD-GL,LD-B则分别记为LD-BH和LD-BL。所有样品均来自沼气池底部,采集后4℃封存,带回实验室后立即对样品进行处理。采集LD-GH、LD-GL、LD-BH和LD-BL样品的发酵温度分别为15.9、5.7、12.3、3.1℃。
1.3 宏基因组DNA的提取和检测
沼气池污泥样品的基因组DNA采用Qiagen公司的QIAamp Fast DNA Stool Mini Kit进行提取,1%琼脂糖凝胶电泳进行检测。在所有DNA样本送交测序公司前,均经过NanoDrop2000型超微量分光光度计检查DNA的含量及纯度。要求样品质量浓度大于10 ng/μL,样品总质量大于500 ng,样品纯度(OD260/280)介于1.8~2.0之间。
1.4 宏基因组测序及基因预测分析
采用Illumina HiSeq 3000测序平台进行高通量测序。测序数据分别经拼接、过滤、质量剪切等优化处理,得到的reads用于操作分类和功能分析;各reads经IDBA-UD (http:∥i.cs.hku.hk/~alse/hkubrg/projects/idba_ud/)软件组装成contigs后,利用MetaGene (http:∥metagene.cb.k.u-tokyo.ac.jp/)软件进行基因预测,并利用cd-hit算法比较相似性获得聚类后的基因序列。选择核酸长度大于等于100bp的基因,并将其翻译为氨基酸序列。
1.5 物种分析与功能分析
使用BLASTP (BLASTVersion 2.2.28+,http:∥blast.ncbi.nlm.nih.gov/Blast.cgi)将基因集序列分别与NR数据库(Amino acid sequence of non-redundant protein,非冗余蛋白质的氨基酸序列数据库)、eggNOG数据库(evolutionary genealogy of genes: Non-supervised Orthologous Groups,http:∥eggnog.embl.de/)和KEGG数据库(Kyoto Encyclopedia of Genes and Genomes,http:∥www.genome.jp/kegg/)进行比对,即可分别获得物种、COG功能和KEGG 功能注释。比对参数e-value 均设置为1×10-5。
2 结果与讨论
2.1 农用沼气池宏基因组数据分析与基因预测
由表1可知,宏基因组测序得到的原始序列经过碱基质量动态去除处理和长度筛选,并经pair-end信息修正后(表1),青海农用沼气池4个样品共获得2.29亿的reads用于操作分类和功能分析。微生物宏基因组共获得2 604 356条组装序列(contigs)。
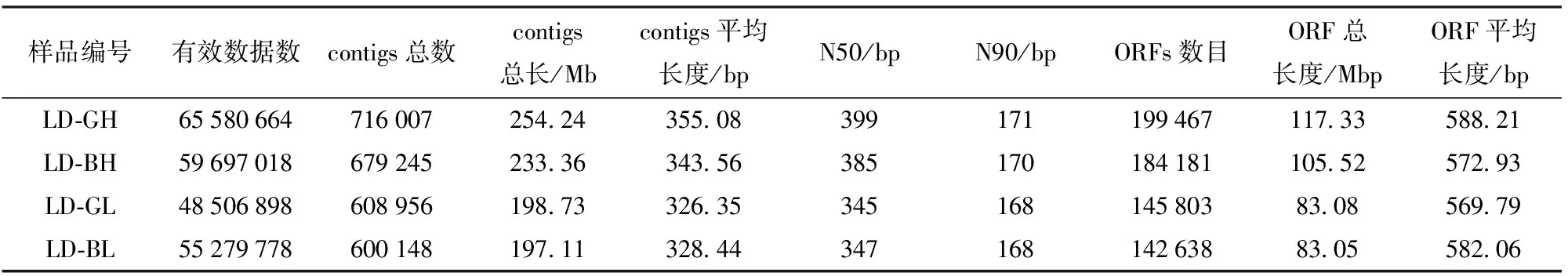
表1 组装及基因预测结果的统计Tab.1 Statistics of metagenome assembly and predicted genes
2.2 农用沼气池微生物群落组成
根据宏基因组序列分析,共得到4域83门158纲364目674科2 299属10 632种。由表2可知,在域分类水平上,所有样品中相对丰度最大的均是细菌,相对丰度为56.41%~86.03%,其次为古菌,相对丰度为3.91%~4.31%。细菌与古菌的总相对丰度为60.32%~90.34%,说明细菌和古菌是青海农用沼气池发酵系统中的主要微生物类群。病毒和真核生物含量较低,相对丰度分别为0.07%~0.11%和0.12%~0.16%。所有样品还包含分类地位尚未明确的微生物类群,相对丰度为9.42%~39.45%,与现有生物数据库中已知序列无相关性,也说明青海农用沼气池发酵系统中生物多样性丰富。综合分析细菌和古菌的相对丰度,同一时期产气好的样品微生物丰度占比均大于产气差的样品,即LD-GH大于LD-BH,LD-GL大于LD-BL。另外,在各样品中病毒均存在,虽然丰度偏低,但可能参与维持沼气池发酵系统的平衡,也可能会对系统中底物降解的效果产生一定影响[16]。因此,青海农用沼气池污泥样品中的病毒类群值得后续深入研究。
2.3 细菌的群落组成
图1显示了青海农用沼气池各样品中细菌在门和属分类水平上的物种组成情况,其中其他类群组(others)表示分别将相对丰度小于1%的门类和属类合并后得到的物种类别。由图1a可知,青海农用沼气池4个样品共计注释门分类13个。所有样品之中,优势门类群依次为拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)和变形菌门(Proteobacteria),总相对丰度(72.37%~74.00%)趋向恒定。通常,上述的3种菌类群是沼气发酵系统中的优势类群,在水解大分子物质、产酸等方面发挥着重要的作用[17-19]。此外,互养菌门(Synergistetes)的相对丰度偏低(1.62%~4.73%),在4个样品中呈现明显的规律性变化,产气好的样品高于产气差的样品,即LD-GH大于LD-BH,LD-GL大于LD-BL,此与沼气产气的变化规律相一致,可能是青海农用沼气池发酵系统中重要的产气功能类群。互养菌门中的所有物种均为严格厌氧菌,广泛分布于胃肠道、污水处理系统、土壤、口腔、厌氧污泥消化器等各种环境中,所占相对丰度较低。早期研究表明其是积极参与厌氧污泥消化的核心细菌类群之一[20-21],可以降解丙酸、丁酸等小分子物质,并能与产甲烷菌进行互营代谢[22-24]。文献[25]在选育耐低温产沼气菌群的过程中,获得一组产沼气菌群5号,将其用厌氧污水驯化后获得另一组菌群Y5S。通过高通量测序技术比较分析产沼气菌群5号、驯化菌群Y5S和厌氧污水原始菌群YJ,结果显示3个菌群均含有Firmicutes、Bacteroidetes、Synergistetes、Proteobacteria、Spirochaetes和Verrucomicrobia。然而,在驯化菌群Y5S中,互养菌门Synergistetes所含丰度最高,是产沼气菌群5号的9.44倍,由此表明Synergistetes门可能是沼气系统中甲烷生产效率的一个重要影响因素。本文的研究结果也证实这一观点,此结论为后续挖掘青海农用沼气池的功能微生物,提供了新的研究方向和思路。
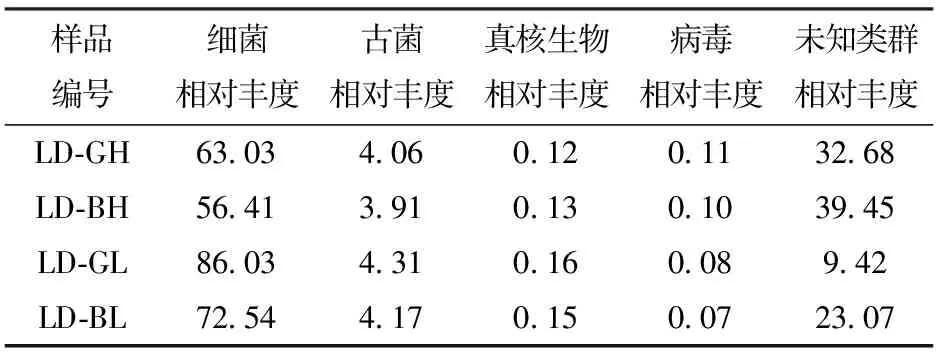
表2 4个样品在域水平上物种的相对丰度Tab.2 Relative abundance of species at domain level in four samples %
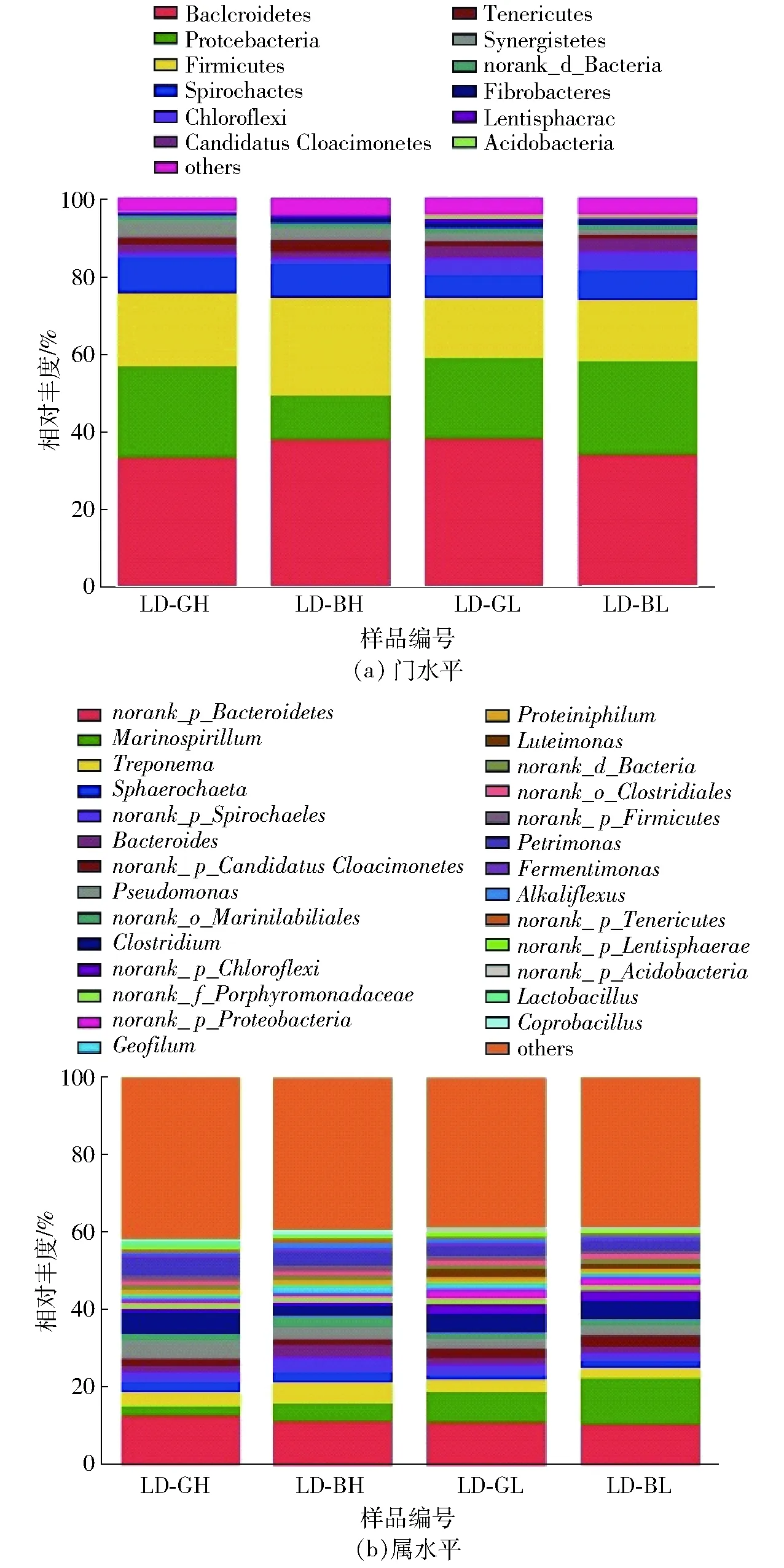
图1 4个样品细菌门和属分类水平统计Fig.1 Bacterial taxonomic classification at phylum and genus level in four samples
青海农用沼气池4个样品共计注释28个属分类(图1b)。在LD-GH、LD-BH、LD-GL和LD-BL样品中,其他类群组(others)的总相对丰度分别为41.70%、39.57%、38.72和38.46%。除其他类群组以外,优势属类群依次为海螺菌属(Marinospirillum,相对丰度6.75%)、梭菌属(Clostridium,相对丰度4.56%)、密螺旋体属(Treponema,相对丰度3.60%)和假单胞菌属(Pseudomonas,相对丰度3.01%)。平行比较分析4个样品,海螺菌属的相对丰度波动变化较大(2.30%~12.46%)。在LD-BL中,海螺菌属的相对丰度(12.46%)最高,其丰度变化与产气存在明显的相关性,丰度越高则产气越差,反之亦然,即LD-GH小于LD-BH,LD-GL小于LD-BL。至此,海螺菌属可能对沼气产气具有一定抑制作用,有待进一步的实验验证。
2.4 古菌的群落组成
分别将相对丰度小于1%的目类和属类合并到其他类群组(others)后,青海农用沼气池4个样品中,共计注释到古菌类群为5目14属(图2)。在目分类水平上,甲烷微菌目(Methanomicrobiales)的相对丰度最高(大于84.90%),且LD-GH大于LD-BH,LD-GL大于LD-BL(图2a)。由图2b可知,14个分类属中,产甲烷菌属(Methanogenium)是优势古菌类群(相对丰度25.41%~31.65%),且丰度变化规律与甲烷微菌目的变化规律相一致。由此表明,在青海农用沼气池发酵系统中,产甲烷菌属是最主要的产气功能类群。产甲烷菌属分类隶属甲烷微菌科,分布广泛,其成员均为氢代谢-营养型产甲烷菌,利用H2/CO2代谢并产生CH4,且大多数属于中温菌(生长温度为30~40℃)[26-27]。极个别产甲烷菌属成员的生长温度宽泛,最低可达到0℃左右,如嗜冷产甲烷菌Methanogeniumfrigidum[28]。本研究中,4个沼气池发酵温度为3.1~15.9℃,属于低温范畴,比甲烷微菌科大多数古菌的正常生长温度要低。在LD-BL样品中,发酵温度仅3.1℃,但产甲烷菌属的丰度仍有25.41%,说明产甲烷菌属可能具有特殊的生理特性和低温适应性。在长期寒冷或温度季节性交替条件下,甲烷菌属参与甲烷的生产,发挥至关重要的作用,可能是青海农用沼气池中最为优势的古菌功能类群[29]。
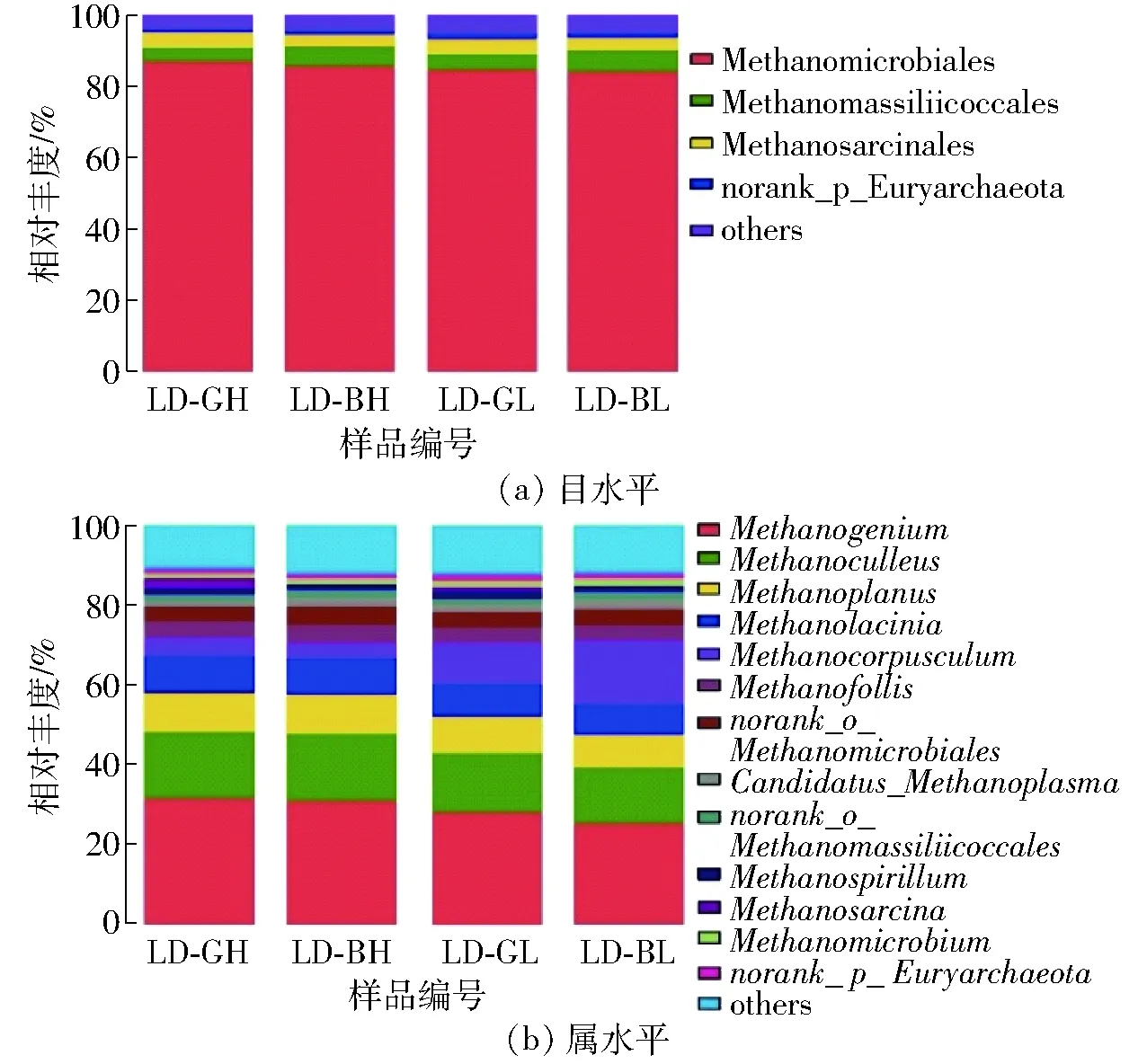
图2 4个样品古菌目和属分类水平统计Fig.2 Archaeal taxonomic classification at order and genus level in four samples
此外,青海农用沼气池还存在几种次优势属群,参与甲烷生产,如甲烷囊菌属(Methanoculleus,相对丰度14.71%)、甲烷粒菌属(Methanocorpusculum,相对丰度11.42%)、甲烷盘菌属(Methanoplanus,相对丰度8.73%)、甲烷裂叶菌属(Methanolacinia,相对丰度8.23%)和甲烷泡菌属(Methanofollis,相对丰度4.19%)。上述这些属群和产甲烷菌属一样,都属于氢代谢-营养型产甲烷菌,参与CO2还原代谢途径,由此表明氢代谢-产甲烷菌可能是青海农用沼气发酵系统中重要的甲烷生产参与者。值得注意的是,甲烷八叠球菌属(Methanosarcina)仅出现在产气好的样品中,如在LD-GH样品中相对丰度为3.47%,在LD-GL样品中相对丰度仅为0.93%,总体相对丰度偏低,且因样品存在差异;但在产气差的LD-BH和LD-BL样品中,均未曾检出。
2.5 COG功能注释
将基因产物与eggNOG数据库比对,获得对应的直系同源序列聚类(COG),得到所有样本COG功能分类统计图(图3)。4个样品中注释到一般功能基因(General function prediction only,R)的相对丰度最高,其次为碳水化合物运输和代谢(Carbohydrate transport and metabolism,G),复制、重组与修复(Replication, recombination and repair,L),氨基酸运输和代谢(Amino acid transport and metabolism,E),翻译、核糖体结构与生物合成(Translation, ribosomal structure and biogenesis,J),能源生产和转换(Energy production and conversion,C),细胞壁/膜生物发生(Cell wall/membrane/envelope biogenesis,M)。此外,未知功能(Function unknown,S)的相对丰度也较高,说明青海农用沼气池样品中可能存在大量未知功能的新型代谢基因。整体而言,注释到各模块的基因丰度,在各样品中分布的变化趋势相对一致,LD-GH略大于LD-BH,LD-GL略大于LD-BL,表明产气好沼气池的样品比产气不好的样品具有更多的有效基因。

图3 COG功能注释图Fig.3 COG function annotation diagram
2.6 微生物群落参与的甲烷代谢关系解析
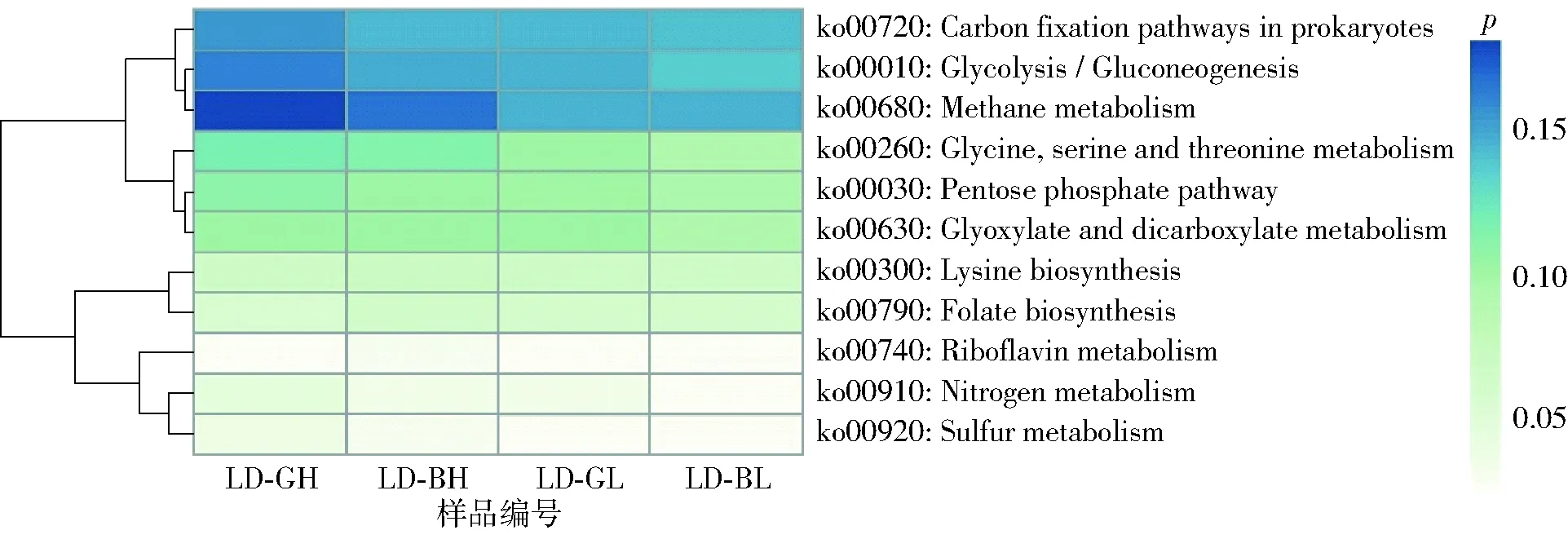
图4 基于甲烷代谢的不同样品KEGG路径热图Fig.4 Heatmaps of KEGG pathways in different samples based on methane metabolism
利用KEGG数据库进行代谢通路分析,共注释获得代谢通路301个,涵盖涉及528 078个基因。参与基因数较多的代谢通路主要集中在氨基酸的生物合成(ko01230)、碳代谢(ko01200)、ABC转运(ko02010)、嘌呤代谢(ko0230)及嘧啶代谢(ko00240)。关联分析甲烷代谢相关的代谢路径,并绘制成热图(图4)。结果显示,4个样品中甲烷代谢(Methane metabolism,ko00680)、原核生物碳固定(Carbon fixation pathways in prokaryotes,ko00720)和糖酵解/糖异生(Glycolysis/Gluconeogenesis,ko00010)的代谢路径较为突出。在4个样品中,上述热点代谢通路的相对丰度均呈现LD-GH大于LD-BH,LD-GL大于LD-BL,直接反映了产气好的沼气池样品具有更强的甲烷代谢能力。
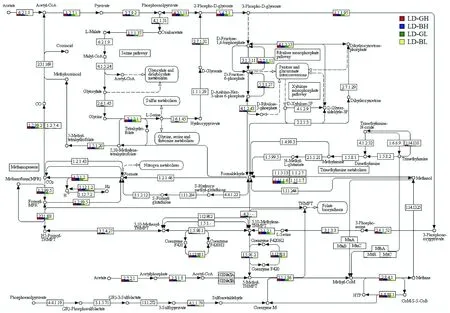
图5 甲烷代谢途径Fig.5 Methane metabolism pathway
在甲烷代谢路径中,产甲烷菌参与的甲烷代谢途径,最终会形成甲基辅酶M(CH3-S-CoM),其在甲基辅酶M还原酶(Mcr,Enzyme commission[缩写为EC]2.8.4.1)的催化下,以CoB为直接的电子供体还原CH3-S-CoM,最终形成CH4[30]。Mcr是甲烷代谢中的关键功能酶,也是产甲烷菌所特有的一种酶,对沼气的产生发挥着至关重要的作用[31-34]。由图5可知,4个样品中产甲烷菌Mcr基因贡献值为LD-GH大于LD-BH,LD-GL大于LD-BL,说明产气好的样品中产甲烷菌Mcr的活性更高。
本研究中,单独分析2种不同类型产甲烷菌参与的甲烷代谢途径。由图5可知,在4个样品中,氢代谢-营养型产甲烷菌参与CO2还原途径的所有通路非常完整。参与这些通路的重要酶,主要涉及四氢甲烷蝶呤甲酰转移酶(EC2.3.1.101)、甲酰四氢甲烷蝶呤环化水解酶(EC3.5.4.27)、甲酰四氢甲烷蝶呤脱氢酶(EC1.5.98.1)以及甲酰四氢甲烷蝶呤还原酶(EC1.5.98.2)等。在4个样品中,基因的贡献丰度为LD-GH大于LD-BH,LD-GL大于LD-BL,表明在产气好的样品中,氢代谢-营养型产甲烷菌以H2作为电子供体,还原CO2产生CH4的能力更强。通常情况下,乙酸代谢-营养型产甲烷菌参与的乙酸代谢过程生产甲烷,首先以乙酸为底物,经乙酸激酶(EC2.7.2.1,Ack)、磷酸转乙酰酶(EC 2.3.1.8,Pta)或乙酰CoA合成酶(EC6.2.1.1,Acs)等酶催化生成乙酰CoA。然后,乙酰CoA再在一氧化碳脱氢酶/乙酰CoA合成酶复合体(Codh/Acs,关键酶),再与四氢甲烷喋呤(H4MPT)经一系列酶及辅酶的催化作用生成CH4。但从图5可知,4个样品中Codh/Acs基因均未检出,说明乙酸代谢-营养型产甲烷菌参与乙酸生产甲烷的代谢通路不完整。本研究中,宏基因组检测到两种乙酸代谢-营养型产甲烷菌,即甲烷鬃菌属(Methanothrix)和甲烷八叠球菌属。前者的相对丰度很低(小于0.1%),从数量上可能导致其能提供的酶基因丰度非常低,近似忽略;而后者尽管有所检出(相对丰度0.93%~3.47%,图3),但由于其具有多细胞集聚,可同时拥有3种CH4合成途径:利用H2/CO2、利用乙酸或利用甲基合成CH4[35]。本研究中,甲烷八叠球菌属可能仅参与利用H2/CO2生成甲烷的代谢途径,而未通过利用乙酸生成甲烷的代谢途径。
此外,青海农用沼气发酵系统中,可能还存在其他微生物参与乙酸的各类生化代谢,从图5可知其代谢过程。首先,乙酰CoA在丙酮酸铁氧还蛋白氧化还原酶(EC1.2.7.1)的催化作用下生成丙酮酸,再在一系列酶的作用下丙酮酸生成亚甲基四氢甲烷蝶呤。然后,由氢代谢-营养型产甲烷菌经CO2还原途径最终生成CH4。对比分析4个样品,其他微生物参与的乙酸代谢通路,涉及所有酶基因的相对丰度为LD-GH大于LD-BH,LD-GL大于LD-BL。通常情况下,沼气系统中所生成的CH4,60%~70%来自乙酸盐的转化,约30%源自H2氧化和CO2还原。本研究发现青海农用沼气池发酵系统与目前的主流观点不一致,可能是因为青海独特的气候类型和地理环境所致,特别是这种特殊的低温发酵条件不利于乙酸代谢-营养型菌株行使功能[36]。因此,青海农用沼气池发酵系统中,甲烷产生的生化代谢过程可能主要依赖于H2/CO2合成途径。
3 结论
(1)基于宏基因组技术获得的物种注释表明,青海农用沼气池发酵系统中蕴藏着丰富的微生物种群。细菌可划分为13门28属,优势门分类类群依次是拟杆菌门、厚壁菌门和变形菌门,在发酵系统中发挥着关键作用。互养菌门可能是沼气生产重要的功能类群。在属水平上,优势类群是海螺菌属、梭菌属、密螺旋体属和假单胞菌属。古菌可划分为5目14属,隶属甲烷微菌目的产甲烷菌属是最优势古菌类群,也是产甲烷的最主要功能类群。
(2)基于代谢功能注释结果表明,产气好比产气差的样品具有更多的有效基因,参与甲烷代谢相关路径的功能基因丰度更高。在所有样品的甲烷代谢途径中,氢代谢-营养型产甲烷菌参与CO2还原途径的全部通路,非常完整,但产气好的样品功能酶基因丰度更高,产甲烷的能力更强。在青海农用沼气发酵系统中,甲烷产生的途径主要依赖于H2氧化/CO2还原的合成。
