重组扬州鹅IFN-α的表达与活性研究
2020-06-05朱孟玲徐孝宙刘海侠王会聪王永娟
朱孟玲,王 岑,曹 霞,徐孝宙,刘海侠,王会聪,王永娟
(1. 江苏农林职业技术学院,江苏 句容 212400;2. 南京农业大学,江苏 南京 210095;3. 南京中医药大学 翰林学院,江苏 泰州 225300;4. 江苏农牧科技职业学院,江苏 泰州 225300)
扬州白鹅,又称扬州鹅,是我国首次利用国内鹅种资源育成的扬泰地方品种,被誉为我国第一个新鹅种,是理想的中型鹅种。但是,其饲养过程常受多种传染性疾病影响,特别是如小鹅瘟、鹅禽流感、鹅法氏囊病、鹅副粘病毒病等病毒性传染病威胁着鹅养殖业的发展[1]。目前,防制病毒性疾病的有效方法主要有疫苗免疫、天然抗病毒药物应用等。干扰素作为一类重要的细胞因子,具有广谱的抗病毒活性、良好的抗肿瘤活性以及高效的免疫调节等功能[2-3]。自1986 年Lefevre 等首次克隆了猪干扰素以来[4],干扰素已从之前的单一成分发展为多成分、多功能制剂,广泛应用于不同畜禽传染病,如新城疫、禽流感、鸡传染性法氏囊病等病毒性疾病的预防与治疗[5]。重组扬州鹅干扰素与其它干扰素一样,具有广谱、高效、安全等优点,是鹅传染病防治中一种重要途径。在鸡、鸭等家禽类的干扰素药物已经投入防病治病的今天,鹅干扰素在世界范围内的研究却只处于起步阶段[6-7],相关的研究报道较少,因此鹅干扰素的研究尤为迫切。
本研究分别通过原核和真核表达系统制备重组扬州鹅干扰素,将原核表达产物免疫小鼠后获得高效价抗体,填补了尚无鹅干扰素抗体的空缺;同时,为深入研究扬州鹅干扰素的分子生物学特性和抗病毒作用机理以及研制鹅病毒性疾病的新型制剂奠定基础。
1 材料与方法
1.1 主要实验材料及实验动物 载体pET-32a(+)、载体pcDNA3.1(-)、工程菌E.coli DH5α菌株、E.coli BL21(DE3)菌株由江苏省兽用生物制药高技术研究重点实验室保存;水泡性口炎病毒(VSV)由扬州大学成大荣教授惠赠;鹅胚和成年扬州鹅由江苏畜禽遗传资源研究所提供;4 周龄雌性BALB/c 小鼠购自扬州大学动物医学比较中心。
1.2 主要试剂 T4 DNA 连接酶、限制性内切酶、RevertAid M-MuLV Reverse Transcriptase、Taq DNA 聚合酶、1 kb DNA Ladderr 购自Fermentas 公司;质粒提取试剂盒、胶回收试剂盒购自爱思进公司;SDS-PAGE 电泳试剂盒、羊抗鼠IgG-HRP、羊抗鼠IgG-FITC、DAB 显色试剂盒购自北京康为世纪生物科技有限公司;5×SDS-PAGE Loading Buffer、96孔酶标板购自生工生物工程(上海)有限公司;ELISA 显色液购自北京索莱宝科技有限公司;胎牛血清、DMEM 培养液购自赛默飞世尔生物化学制品有限公司;脂质体转染试剂盒Lipofectamine 2000 购自Invitrogen 公司;Triton X-100 购自上海江莱生物科技有限公司;中性红指示剂购自上海研臣实业有限公司。
1.3 扬州鹅IFN-α全长基因和成熟肽基因的扩增根据GenBank 中鹅α-干扰素(GoIFN-α)基因序列(HQ115583)设计扩增GoIFN-α基因全长序列的引物(F1:5'-ATGCCTGGGCCATCAGCCCCAC-3'/R1:5'-TTA GCGCATGGCGCGGGTGAGG-3'),以及扩增成熟肽基因的引物(F2: 5'-AAGGATCCTGCAGCCCCCTGCGCCTC-3'/R2:5'-GCAAGCTTTTAGCGCATGGCGCGGG TGA-3'),引物由上海英潍捷基公司合成。
采鹅翼静脉抗凝血,按照淋巴细胞分离液说明书分离淋巴细胞,按照TRIzol 说明书提取淋巴细胞总RNA,反转录获得cDNA,以其为模板, 分别以F1/R1、F2/R2 进行PCR 扩增,产物经1.5%琼脂糖凝胶电泳检测。
1.4 GoINF-α全长基因和成熟肽基因的克隆与鉴定 将纯化回收的扬州鹅IFN-α 成熟肽基因与pET32a(+)连接,将连接产物分别转化E.coli DH5α受体菌, 增菌培养后提取重组质粒经BamH I/Hind III酶切鉴定,阳性产物命名为pET-mGoIFN-α。
将回收的扬州鹅IFN-α 全长基因克隆于pcDNA3.1(-)载体,重组质粒命名为pcDNA-GoIFN-α,经Xho I 酶切鉴定后阳性重组质粒由上海英潍捷基公司测序鉴定。
1.5 高免血清制备 常规方法诱导pET-mGoIFN-α/BL21。表达产物经SDS-PAGE 检测,1 mol/L KCL 染色,切下预期大小的蛋白条带液氮研磨,BCA 法测其浓度。按100 μg/只重组蛋白颈背部皮下注射免疫5 只BALB/c 小鼠,每10 d 免疫一次,4 免后第3 d 再加强免疫,眼眶静脉采血,分离血清,备检。
1.6 抗体效价与特异性检测 以纯化后的mGoIFN-α作为抗原, 羊抗鼠IgG-HRP(1∶100 000)作为二抗,ELISA 法测定血清中扬州鹅IFN-α成熟肽抗体效价。设置未免疫小鼠血清为阴性对照。同时,以纯化的mGoIFN-α为抗原,收集的免疫小鼠血清作为一抗,羊抗鼠IgG-HRP(1∶10 000)为二抗,western blot 检测和分析抗体特异性。
1.7 GoIFN-α抗病毒活性检测 参考脂质体转染说明书,将制备的无内毒素的pcDNA-GoIFN-α质粒转染自制的鹅胚成纤维细胞(GEF),以自制的抗mGoIFN-α抗体为一抗,羊抗鼠IgG-FITC(1∶10 000)为二抗,采用间接免疫荧光试验(IFA)检测GoIFN-α重组蛋白(rGoIFN-α)表达情况。确定表达后将其大量转染DEF,收集细胞及上清,冻融后离心收集上清,并用含2% FBS 的DMEM 培养液10 倍比稀释(100~10-10),保存备用。
在96 孔细胞培养板上培养GEF,待单层细胞长至约90%时接入倍比稀释的rGoIFN-α,继续37 ℃,5% CO2培养24 h 后每孔接入100 TCID50VSV。同时设空白对照组(仅GEF)和病毒对照组(仅VSV)[8-10],每隔12 h 观察一次。当发现病毒对照组75%出现病变时,用含中性红染液的DMEM 染色,酸性乙醇脱色,测定OD550nm值。以能够保护50%的GEF 细胞免受VSV 攻击的稀释度为一个活性单位(U),确定蛋白的抗病毒活性。
2 结 果
2.1 GoINF-α基因扩增鉴定 利用RT-PCR 法扩增GoIFN-α成熟肽基因和IFN-α全长基因,分别获得486 bp 和576 bp 的片段,与预期结果一致(图1),且测序结果正确,表明扩增得到目的基因。
2.2 重组质粒鉴定 重组质粒pET-mGoIFN-α 经BamH I 与Hind III 双酶切后,在约500 bp 和5 900 bp处可见目的条带(图2);重组质粒pcDNA-GoIFN-α用Xho I 酶切后检测,在约400 bp 与5 500 bp 处可见目的条带(图3);均与预期相符,表明重组表达质粒正确构建。
2.3 重组蛋白表达鉴定 pET-mGoIFN-α/BL21 经诱导表达后SDS-PAGE 鉴定, 结果显示: 在36.3 ku处出现目标条带,与预期大小相符(图4),表明重组蛋白以包涵体的形式存在且表达量较大。

图1 PCR 扩增产物鉴定Fig.1 The identification of PCR amplicons

图2 pET-mGoIFN-α的酶切鉴定Fig.2 Restriction analysis of pET-mGoIFN-α
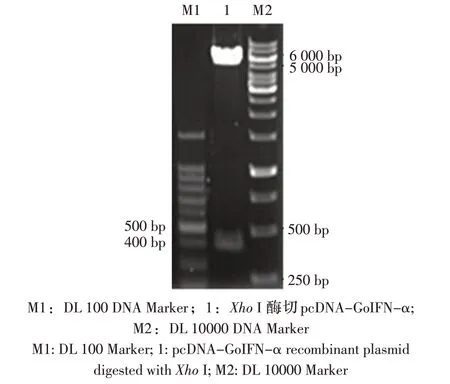
图3 pcDNA-GoIFN-α 的酶切鉴定Fig.3 Digestion analysis of pcDNA-GoIFN-α
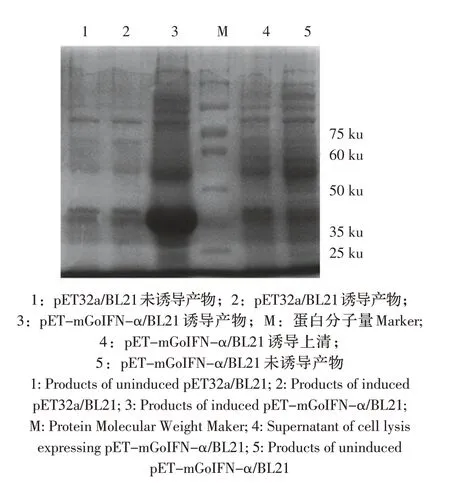
图4 mGoIFN-α的表达Fig.4 Expression of mGoIFN-α
2.4 抗体效价与特异性检测 ELISA 检测结果显示免疫鼠抗mGoINF-α 制备的抗体效价为1∶16 000。Western blot 分析结果显示,在目的蛋白对应位置36.3 ku 处出现特异性显色(图5)。表明免疫鼠抗体具有较强的特异性。
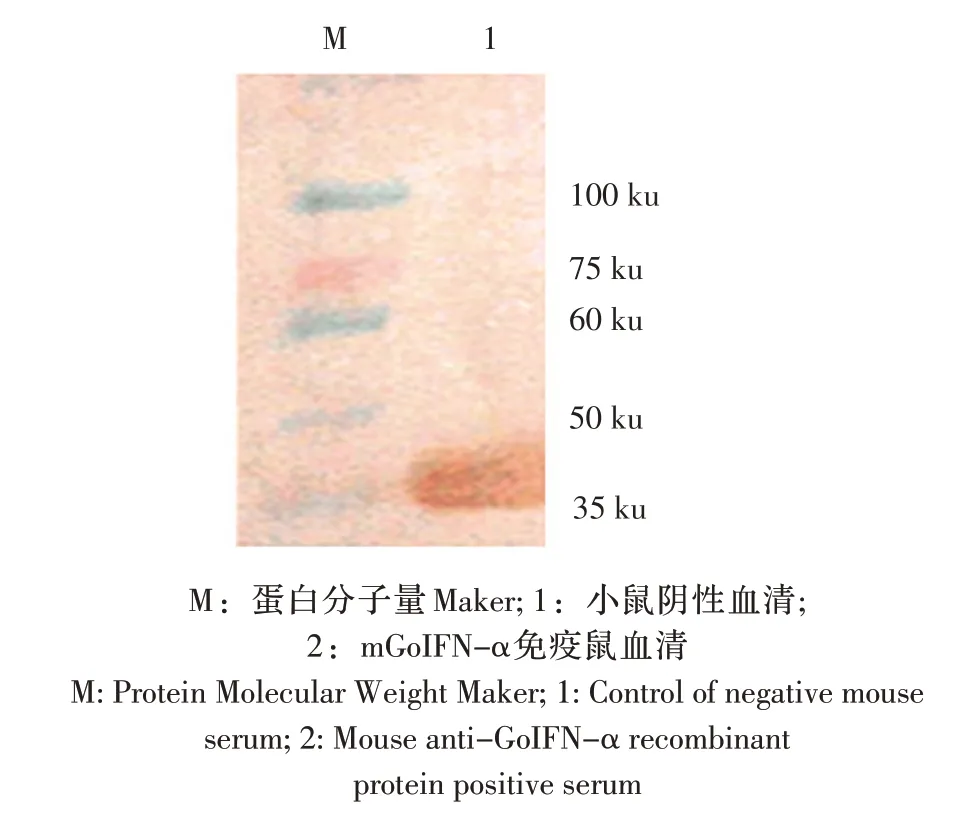
图5 鼠抗血清western blot 检测Fig.5 Reactivity analysis of immune serum in mice by western blot
2.5 rGoIFN-α 在GEF 中 的 表 达 检 测 pcDNAGoIFN-α转染GEF 48 h 后,IFA 检测rGoIFN-α的表达情况,结果显示,pcDNA-GoIFN-α转染的GEF 中出现荧光,且细胞核周围荧光亮度最强,而空载体pcDNA3.1转染的GEF中无荧光(图6)。表明rGoIFN-α在GEF 中得到表达。
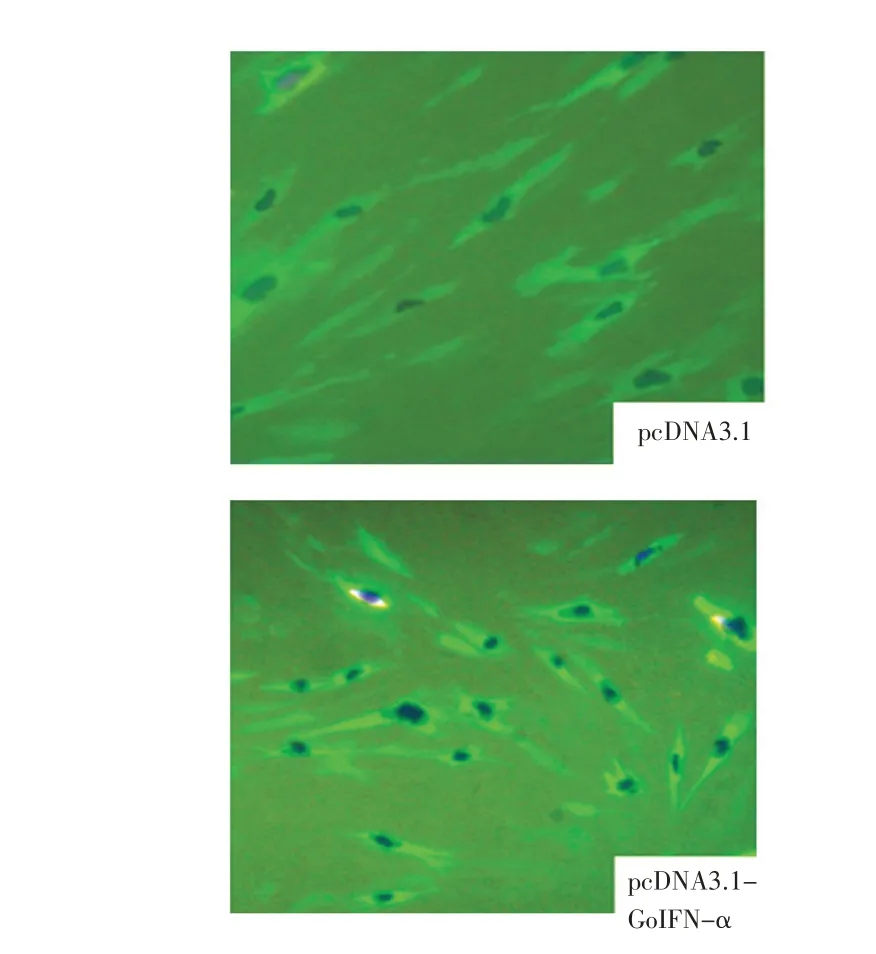
图6 rGoINF-α表达的IFA 鉴定Fig.6 Indirect immunofluorescence analysis for pcNDA-GoIFN-α expression
2.6 rGoIFN-α抗VSV 活性检测 采用细胞病变抑制法,在GEF-VSV 系统中检测rGoIFN-α的抗病毒活性。接毒后约36 h,经显微镜观察:正常生长的GEF 未出现病变(图7A); 加入不同稀释度rGoIFN-α的细胞均发生了轻微病变,稀释倍数越低GEF 病变越弱,甚至有些未出现病变;稀释倍数越高GEF 皱缩状态越严重,与只加入病毒的阳性对照组相比,细胞残缺程度明显较轻(图7B);VSV 阳性对照组GEF 严重皱缩,病变程度高(图7C)。

图7 抗病毒活性鉴定Fig.7 Antiviral activity analysis
计算真核表达的rGoIFN-α抗VSV 活性约为1.20×105U/mL,比 活 性 为3.84×104U/mg(n=3)。表 明rGoIFN-α能够保护GEF 免受VSV 攻击,具有较高的抗VSV 活性。
3 讨 论
表达外源蛋白主要采用植物表达系统、原核表达系统、真核表达系统等。原核表达系统具有生物结构简单、生长周期短、遗传稳定、价格低廉、操作简便、菌体密度高、便于快速大规模发酵培养等优点[11],同时原核表达系统从诱导直至表达的整个过程均相对易控,是多数蛋白表达科研工作者的首选系统。本研究选用大肠杆菌原核表达系统进行表达,主要是基于大肠杆菌可以快速表达外源蛋白以制备免疫原的优点,能在短时间内获得可用于动物免疫的纯化蛋白,进而获得特异性抗血清。但是原核表达产物常以包涵体的形式存在,包涵体的纯化、变性、复性操作繁琐[12],且蛋白容易失去活性,所以本研究又选择了能对外源蛋白进行加工修饰的真核表达系统,以确保所表达的重组INF 接近天然状态,具有较好的活性[13-14]。以自制的抗rGoIFN-α抗体为一抗,IFA 检测真核表达产物,结果显示,rIFN-α能在GEF 中表达,在细胞核周围的荧光尤为明显,从细胞核向胞浆发散。但是多次重复实验后发现,在相同的转染时间后,每个细胞的荧光亮度不尽相同,推测这可能与GEF 自身的形态大小与其摄入目的基因的片段量的多少有关。
本研究将原核表达的mGoIFN-α经SDS-PAGE 电泳切胶后的产物作为抗原,用液氮对凝胶进行辅助研磨,直接注射入小鼠体内,并没有额外加入免疫佐剂,却获得了效价较高的特异性抗体,这可能是跟凝胶本身也可作为一种特殊的佐剂发挥缓释功能有关[15-16]。另外,采用切胶回收制备免疫原,制备方法简单、回收量大、产物纯度高、具有佐剂活性、成本低廉,可以克服常规过柱纯化方法的一些缺点。
与其它禽类IFN 相比,GoIFN-α 研究起步较慢,在世界范围内的研究只处于刚刚起步阶段。本研究主要围绕抗病毒功能较强的扬州GoIFN-α分别做原核表达和真核表达,制备的高效价抗体解决了目前没有抗GoIFN-α抗体的问题,也为真核表达时的IFA 检测提供了必须的生物材料;真核表达的重组扬州鹅IFN-α具有较好的抗病毒活性,为进一步开展扬州鹅IFN 的分子生物学特性和抗病毒作用机理、研制防治鹅病毒性疾病的新型制剂奠定了基础。
