丁苯酞对慢性酒精中毒大鼠内质网应激相关基因的影响
2020-05-29董家行姜万举刘佳聪朱甲婧张占蕊张瑞岭杜爱林
董家行 姜万举 刘佳聪 朱甲婧 张占蕊 张瑞岭 杜爱林
(1新乡医学院生理学与神经生物学教研室 中英脑功能损伤联合实验室 河南省高校脑研究重点实验室培育基地,河南 新乡 453003;2新乡医学院第二附属医院 河南省生物精神病学重点实验室)
慢性酒精中毒在全世界许多国家都是一个社会性健康难题,据调查,我国约4.7%的成年居民存在过量饮酒的状况,很多地区都存在着酒精滥用的现象,对社会生产力的稳定发展造成了较大的威胁〔1〕。慢性酒精中毒患者会出现需要饮酒的强迫性体验,而在停止饮酒后,往往容易出现戒断综合征。患者时常会恶心、呕吐、心中难受,甚至会表现出肢体震颤等躯体疾病,然而这些症状在饮酒后则会自行消除,同时会出现幻觉、妄想及相应的情绪和行为异常,易有攻击或暴力行为,甚至自杀。有报告指出,酒精中毒者的自杀率比一般人群高2~4倍,10%~15%的嗜酒者有自杀行为〔2,3〕。此外,长期不良的饮酒嗜好能够促进各类躯体及神经系统疾病的发生发展,让患者难以适应社会,人际关系紧张,更易失业,使经济更加困难。有关研究表明慢性酒精中毒可导致大脑萎缩〔4,5〕,过量酒精会损伤记忆相关的重要脑区,如海马区、前额叶皮层等,导致慢性神经炎症的发生、神经营养物质减少及神经毒性物质释放,进而导致神经细胞大量凋亡〔6,7〕。而目前的科学研究水平针对这种病尚无良好的药物来进行治疗。近年来,对慢性酒精中毒性引发的大脑神经退行性疾病的病理研究,为慢性酒精中毒后的生物信息学研究及发展打下了坚实的基础。丁苯酞(NBP)是中国自主研发出的抗缺血性脑卒中药物。研究证明NBP可通过提高脑血管内皮一氧化氮(NO)和前列环素(PGI)2的水平,抑制谷氨酸释放,降低花生四烯酸含量,抑制氧自由基活性等机制对脑缺血产生治疗效果〔8〕。NBP能影响海马内硫化氢(H2S)及CA1、CA3区5羟色胺(HT)表达量进而改善慢性酒精中毒症状〔9〕。动物药效学研究表明,NBP抗脑缺血效果良好,能够增加缺血区毛细血管数量,显著改善脑区的微循环,减轻脑水肿,缩小大鼠局部脑缺血的梗死面积〔10〕。研究表明,NBP可改善线粒体功能和促进脑能量代谢,抑制神经细胞的凋亡,抑制血栓形成等〔11〕。因此推测,NBP可能对慢性酒精中毒导致的神经功能损伤有改善作用,可促进患者功能恢复。本实验基于NBP干预后慢性酒精中毒大鼠大脑海马的基因表达数据集,结合了生物信息学的分析方法,筛选出差异表达基因,采用实时荧光定量聚合酶链反应(qPCR)对关键mRNA的表达进行验证。试图从基因层次探讨NBP对慢性酒精中毒的影响及其改善机制。
1 材料与仪器
1.1实验动物 清洁级健康雄性SD大鼠36只,由郑州大学动物中心提供。实验动物随机分为正常组、模型组和用药组各12只。喂养4 w后,统一处死,取大脑海马区组织。正常组喂养蒸馏水,用植物油进行灌胃。模型组喂养6%的酒精溶液4 w,并用植物油进行灌胃,造成慢性酒精中毒;而用药组也喂养6%的酒精溶液,前2 w用等量的植物油进行灌胃,后 2 w则用等量的植物油稀释过的NBP 20 mg/kg进行灌胃。
1.2仪器与试剂 主要试剂耗材为Illumina Gene Expression Sample Prep Kit和Solexa测序芯片(flowcell),主要仪器是Illumina Cluster Station和Illumina HiSeqTM2000系统,均来源于华大基因公司。BNP(批号:118170424)购于石药集团恩比普药业有限公司。
Morris水迷宫是由圆形水池、隐藏于水面之下的可移动平台、图像自动采集和处理分析系统组成。圆形水池直径为150 cm,高50 cm,以池壁上的4个等距离点划分为4个象限(东北,东南,西南,西北),每个象限池壁圆弧中点为大鼠入水点,平台放置于SW象限中心,并没入水面以下2 cm。图像自动采集和处理分析系统记录动物游泳轨迹数据用于指标的提取及分析。实验恒定,水温控制在25℃,在水池周围安放蓝色不透光布帘保证水池水面光线均一,实验期间保证环境的安静。
1.3定位航行实验 定位航行实验能够有效检测出实验动物在水迷宫中的学习记忆能力。实验开始前30 min将大鼠置水迷宫所在房间以适应环境,将大鼠用手托起令其面向池壁,以半随机方式选择入水点轻轻放入水中,每天每只训练 4 次,4个不同象限各1次,实验进行4 d,每只共计训练16次。若在60 s内登陆上平台,则将其搜寻并登上平台花费的时间作为潜伏期。反之,实验者则需要将其引导至平台,同时要将潜伏期记录为60 s。待登陆平台后,让其在平台上停留30 s。每天实验后及时统计并计算各组游泳距离、潜伏期和平均游泳速度。
1.4芯片数据处理及差异表达基因(DEGs)的筛选 处死大鼠后,每组6只完整取出大脑海马组织,并快速放入液氮中保存。提取出海马中的RNA,使Illumina基因表达的样品试剂盒和Solexa测序芯片对模型组及用药组的海马基因表达进行检测,并获取相应的基因表达数据集。借助基因芯片获取了原始的慢性酒精中毒大鼠海马基因表达数据集,之后对该数据集进行一系列预处理,并参照Audic〔12〕的数字化基因表达谱差异基因检测方法,筛选出了最终的差异表达基因的数据。
1.5基因功能显著性富集分析 基因本体(GO)能够对差异表达基因进行功能分类,它可从分子功能、细胞组成和生物活动过程等3个层面对目标基因进行详细的描述。在获取海马mRNA的表达谱后,借助GO体系对DEGs进行精准注释,并进一步做富集化分析,最终得到所有差异基因中明显富集的所有分类条目,方便对慢性酒精中毒后转录组学进行深入的研究。
1.6信号通路显著性富集分析 京都基因与基因组百科(KEGG)数据库可以快速查询到基因所在通路的详细信息,包括其代谢途径、所处的细胞信号转导通路、产物及相互作用的基因等信息,帮助研究者高效地定位具体的研究方向。同时还可以对未知的序列比对(BLAST)序列进行比对,找出具体代谢途径,迅速定位某段序列所处的通路,其功能很强大,数据覆盖广泛。借助KEGG对所有差异表达基因进行信号通路富集分析,将得到的详细的通路富集信息表读取到R语言中,加载并运行专业用于科研绘图的ggplot2包,将信号通路条目作为纵坐标,把各个转导通路富集的P值作为横坐标,绘制出一张详细的信号通信富集气泡图。在该气泡图中,各个通路代表的气泡越接近纵坐标轴,即表明其结果可信度越高;气泡越大则代表信号通路中所包含的差异基因的总数越多;颜色越偏向红色则显示该条信号通路的富集程度更高。
1.7蛋白质间相互作用网络构建 功能蛋白关联网络分析(STRING)数据库囊括了大量的蛋白质功能信息,并且能够精准比对未知的BLAST序列,给出其详尽的功能简介。且该数据库还可以对不同的蛋白质之间直接或间接功能的相关性进行查询,支持对目标蛋白质和未知蛋白质之间是否存在相应的作用方式展开预测,可以有效地帮助研究者对获取的差异基因表达谱及蛋白间互作方式进行综合研究。借助STRING数据库研究蛋白质间的相互作用关系,并将相关数据导入到Cytoscape,使用MCODE插件对其子网络进行聚类分析。
1.8qPCR检测关键基因的表达量 完整取出大鼠的海马体组织,从两组中各筛选出3个样本进行qPCR实验,用1 ml TRIzol制备匀浆,提取总RNA,测定其纯度及浓度,按试剂盒要求进行反转录操作,并进行PCR扩增。实时荧光定量PCR体系由20 μl的反应体系组成:SYBR(20×) 0.3 μl,Platinum®Taq DNA聚合酶(5 U/L)0.2 μl,ddH2O 14 μl,10×PCR缓冲液(-)2 μl,Mg2+(50 mmol/L)1 μl,dNTPs(10 mmol/L) 0.5 μl,正义链(10 mol/L) 0.5 μl,反义链(10 mol/L)0.5 μl,Template(就是反转录产物,也即cDNA)1 μl。反应中的Ct值数据的采集采用校正的阈值设定,实时荧光定量PCR 的方法以β-肌动蛋白(ACTB)作为内参基因,采用2-ΔΔCt法进行相对定量。 扩增条件为:95℃ 60 s,95℃ 5 s,60℃ 40 s,反应40个循环,引物序列:SEC61g上游:5′-GGACTCGATTCGGCTGGTTA-3′,下游:5′-GCCGATGAACCCCATGATAG-3′;NDUFA4上游:5′-TGCAGGAGCACCTGAGTCAGT-3′,下游:5′-CAAGGAAGGCACCGGATCT-3′;NDUFA2上游:5′-CAACGGTACGTGGAGCTGAA-3′,下游:5′-ACCTCAGCAGCACTCAGATTGT-3′。
2 结 果
2.1模型组与正常组学习记忆功能的对比 第1天,两组平台潜伏期无统计学差异(P>0.05);第2~4天模型组平台潜伏期显著延长(P<0.001),提示模型组学习记忆能力受到损伤。见表1。

表1 正常组与模型组水迷宫平台潜伏期的 比较
与正常组比较:1)P<0.001
2.2数据预处理及DEGs的筛选 对芯片数据进行相应的处理后,与模型组相比,用药组共585个DEGs。320个基因表达上调,265个基因表达下调。模型组与用药组相比有208个DEGs。
2.3差异基因的GO富集分析和通路分析 选取富集程度较高的前10个GO条目进行分析。用药组与模型组之间对比,GO富集的生物学过程分析表明差异基因主要涉及膜去极化调节,磷酸盐代谢过程,磷的代谢过程,生物过程调节,生物调节,抗原刺激的炎症反应,Ⅰ型干扰素产生的调节,Ⅰ型干扰素产生的正调控作用,细胞代谢过程,C4二羧酸转运等生物学过程,见表2;分子功能表明,DEGs主要涉及催化活性,作用于酯键的水解酶活性,磷蛋白磷酸酶的活性,二羧酸的跨膜转运活性,(醌)活性,磷酸酶的活性,酸性氨基酸跨膜转运活性,蛋白质丝氨酸/苏氨酸激酶活性,阳离子:糖转运体活性,水解酶活性等功能,见表3。KEGG富集分析结果见图1,富集程度最高的前10条信号通路主要与氧化磷酸化通路,黏合连接,蛋白酶体通路,脂肪细胞因子信号通路,丝裂原活化蛋白激酶(MAPK)信号转导通路,破骨细胞的分化,黏着,幽门螺杆菌感染中的上皮细胞信号转导,帕金森病通路,聚酮糖的生物合成等信号通路相关。表4、5是参与细胞信号通路中的DEGs。
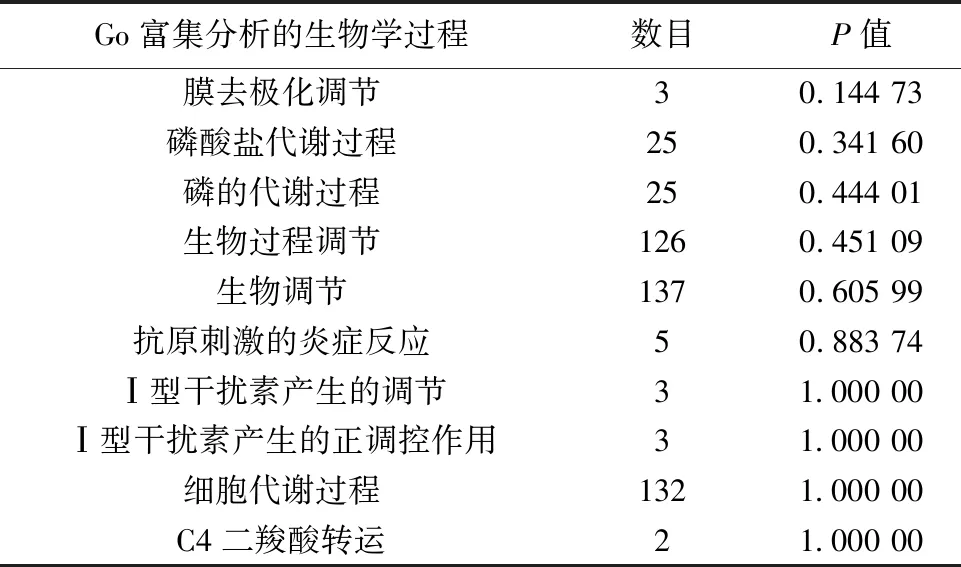
表2 差异基因涉及的生物过程
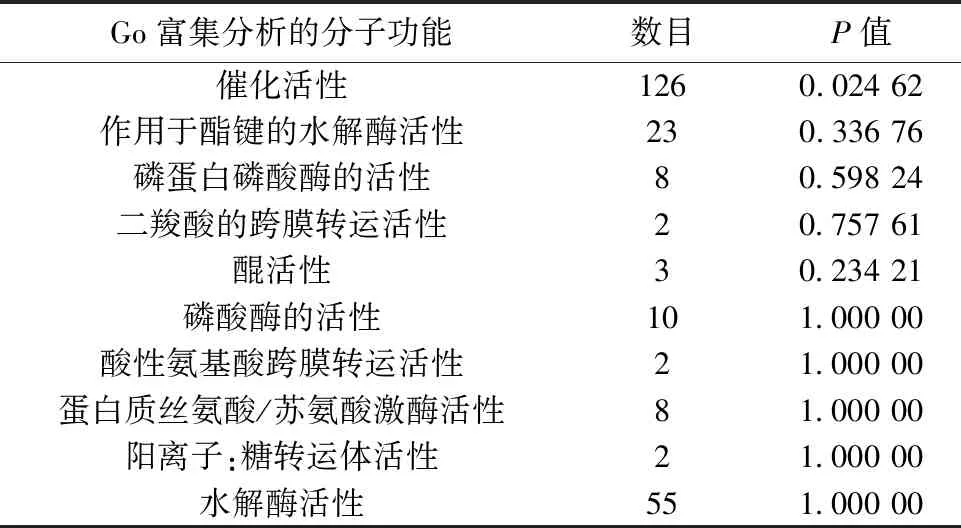
表3 差异基因涉及的分子功能
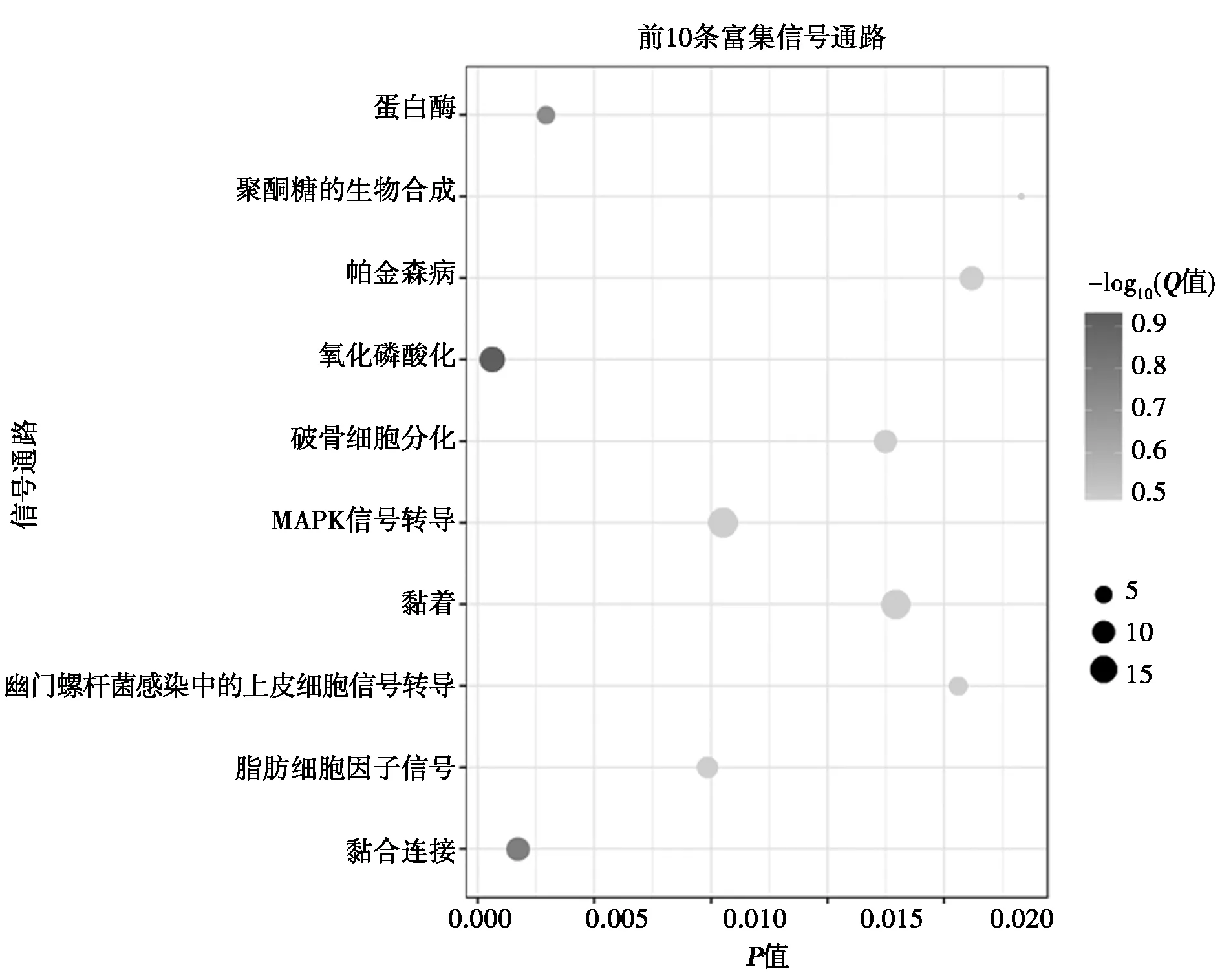
图1 信号通路富集
2.4差异基因编码蛋白的相互作用分析 蛋白质互作网络图中的线代表两个蛋白之间有相互作用关系,而线的数量记作度。对这些差异表达基因进行了进一步的筛选。模型组相比正常组出现差异表达的208个基因,用药组相比模型组发生差异表达的585个基因,取了一个交集。共获得了107个DEGs,这107个差异表达基因就是大鼠在慢性酒精中毒以后出现差异表达,而后又经过NBP治疗被反向调控的。提示NBP可能通过调控这107个基因对慢性酒精中毒损伤起到了一定的保护作用。
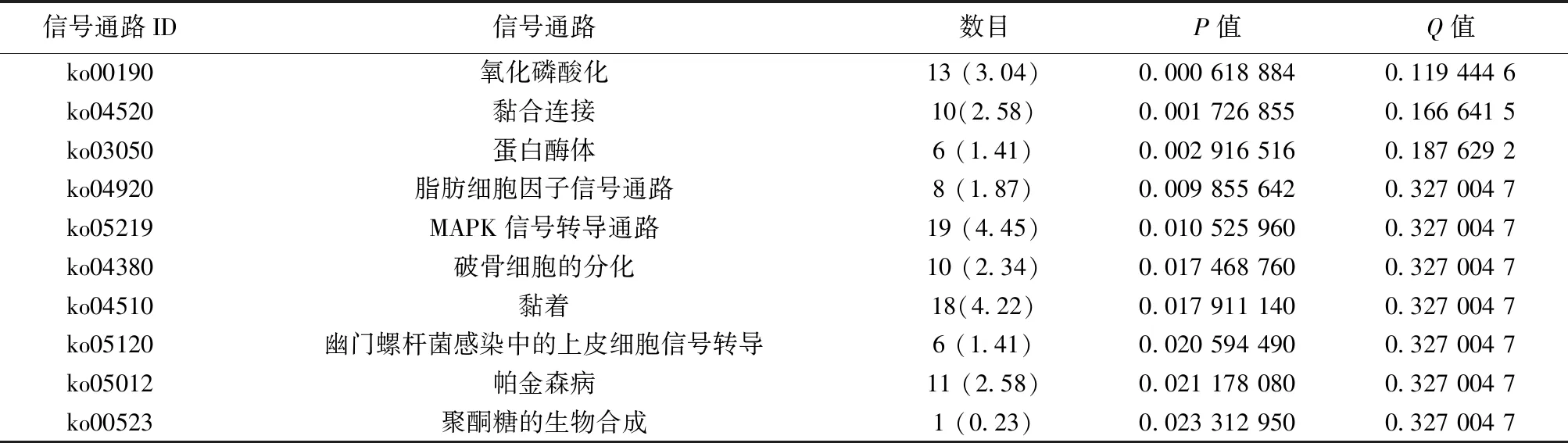
表4 差异基因涉及的信号通路富集分析〔n(%)〕

表5 富集程度最高的前10条通路中所参与的差异基因
随后在STRING数据库在线系统上读取了107个DEGs的基本信息,并运行相应的预测工具,结果显示仅102个基因能在数据库中搜到相对应的信息,在得到初步的蛋白互作图后,将孤立的蛋白除去,并将数据表格导出(见图2),再将其导入到Cytoscape软件中进行可视化操作及中心网络节点分析,再排除degree为1且分散的局部蛋白互作网络,才得到最终的中心蛋白互作网络图(见图3),同时借助MCODE插件分析出最重要的2个子网络(见图4)。其中RPSA、RPS9、RPS11、SRP19、sec61G、EIF3E、LARS、Ndufa4、Ndufb6、Ndufa2等是与周围蛋白相互作用最密集的前10个蛋白,属于该相互作用网络的中心蛋白(见表6),删除这些中心蛋白后,网络结构涣散。
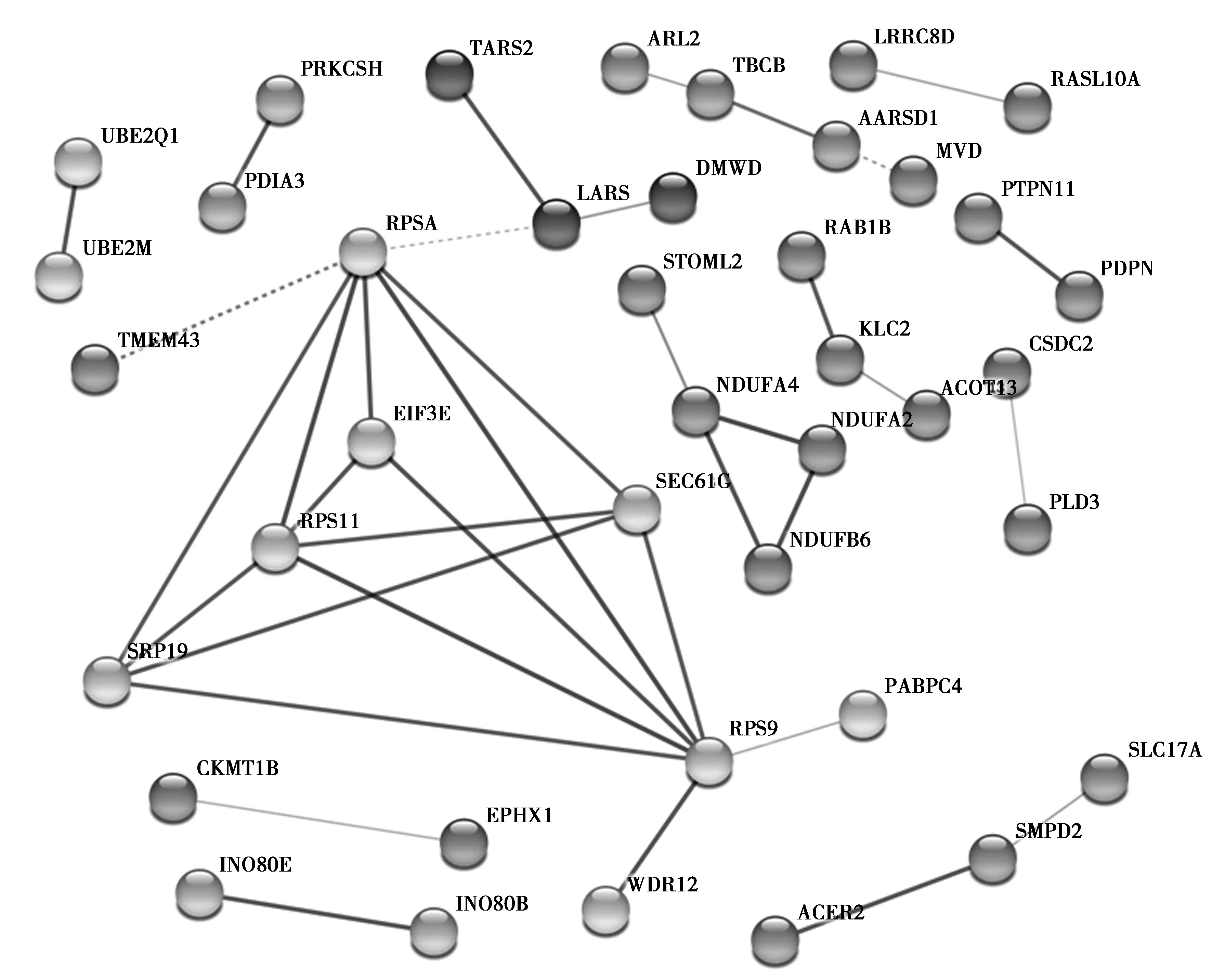
图2 差异基因编码蛋白质互作网络
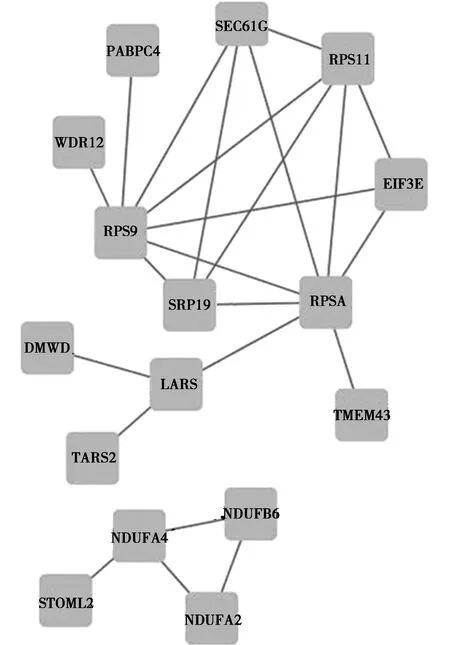
图3 中心蛋白互作网络
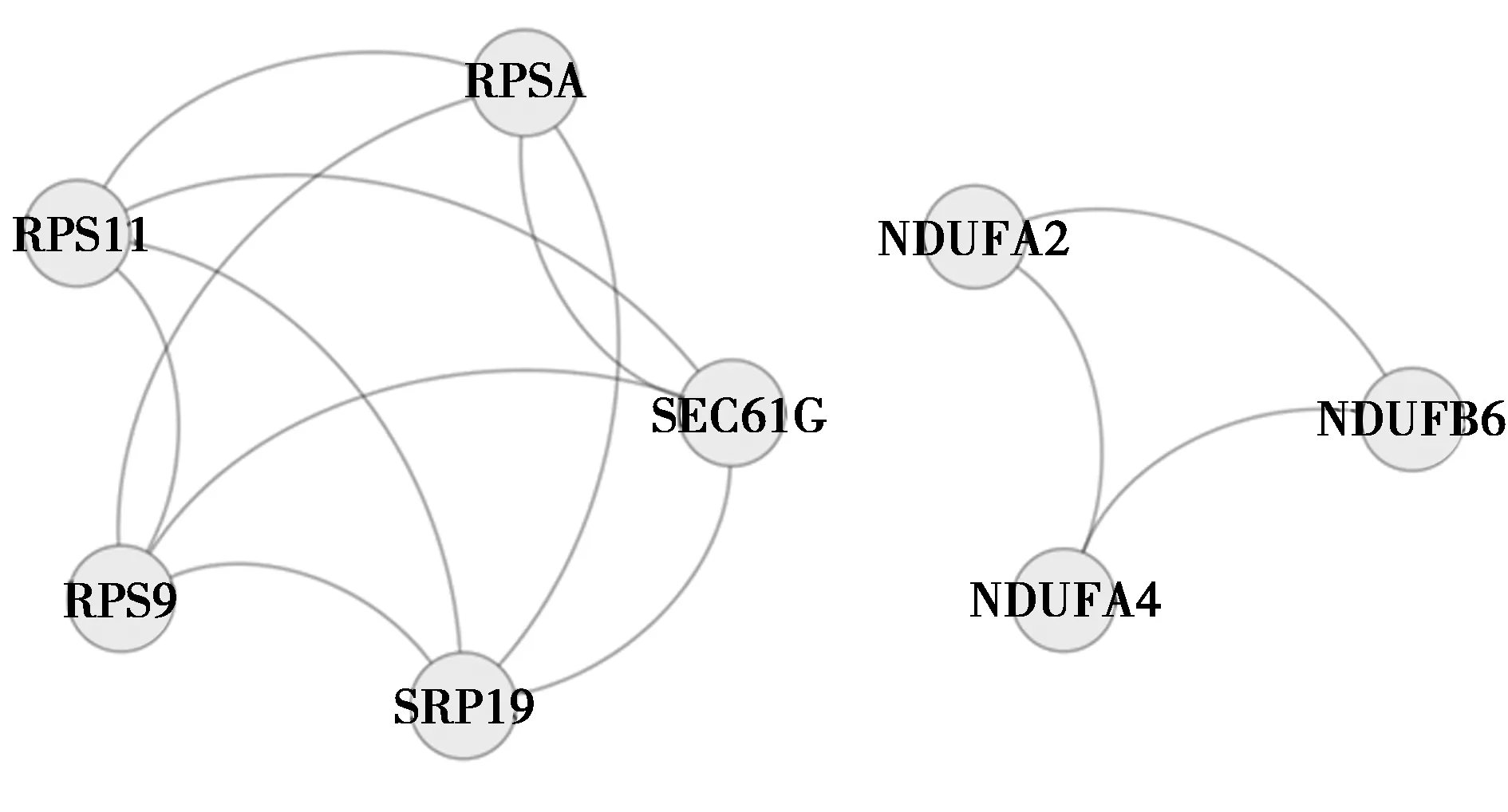
图4 最重要的2个子网络
2.5关键mRNA的相对表达量 对Sec61g、Ndufa4、Ndufa2等关键基因进行检测,慢性酒精中毒以后,这3个与内质网及线粒体相关的mRNA表达量显著降低,提示内质网及线粒体出现了一定的损伤。然而与模型组相比,用药组海马体内的Sec61g、Ndufa4、Ndufa2的相对表达量有较为显著提高(P<0.05或P<0.01),见表7。提示BNP可能对内质网应激及线粒体损伤的修复作用。使用GraphPad Prism对以上结果进行处理。
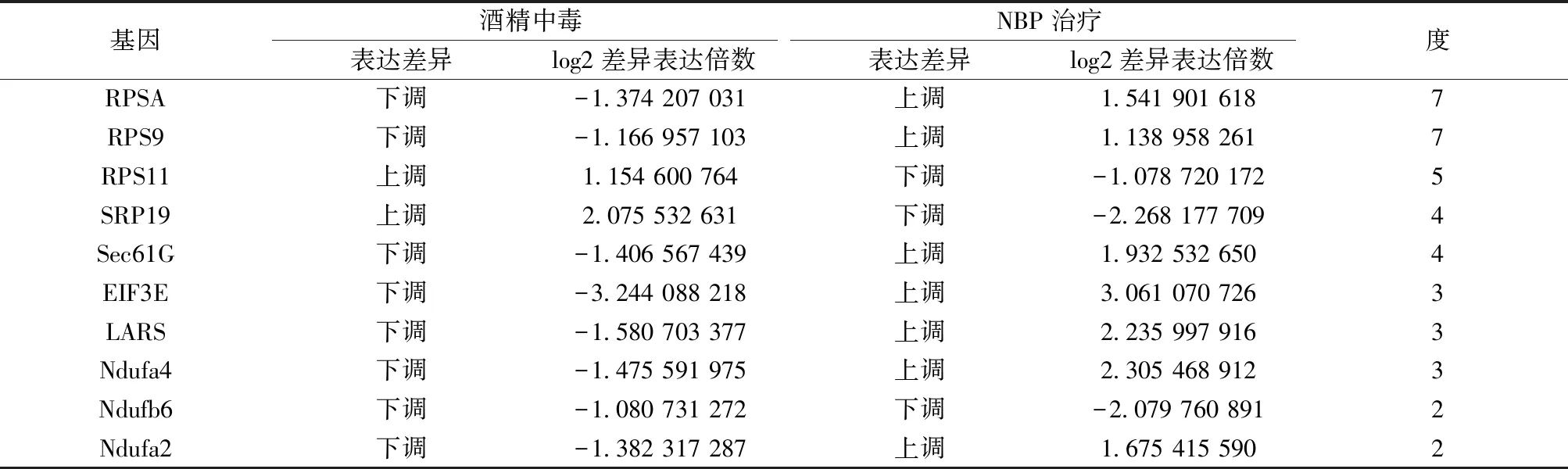
表6 PPI网络中心蛋白
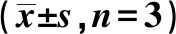
表7 Ndufa2、Ndufa4和Sec61g mRNA的 相对表达量
与正常组比较:1)P<0.05,2)P<0.01;与模型组比较:3)P<0.01
3 讨 论
易位通道是新生的肽链在共翻译转运时所经过的通道。由于N端带有正电荷,新生的肽链无法轻易地进入表面疏水的内质网膜,所以需要借助1个蛋白引导通道才能进入内质网,从而完成蛋白质的转运,这样的通道就是易位通道。
Sec61异源三聚体,包含α、β和γ 3个亚基。在真核生物中,Sec61异源三聚体就是易位通道的核心。它的主要作用是能够形成一个疏水通道,从而帮助新生的肽链穿过内质网膜进入内质网。Sec61复合体、内质网膜蛋白与核糖体共同的作用才能形成易位通道。通过Sec61和易位子相关蛋白(TRAP)结合,为分泌蛋白和膜蛋白转运蛋白的转运提供便利。然而慢性酒精中毒以后Sec61γ的表达下调可能影响与TRAP组成的复合体的活性导致蛋白质转运过程中未折叠或错误折叠的蛋白质滞留内质网或细胞质中,使得内质网分子伴侣基因表达激活,并引发内质网应激反应(UPR)〔13〕,损伤内质网的正常生理功能。 UPR 主要由3种内质网跨膜受体介导:肌醇需求酶(IRE)1、PKR样内质网激酶(PERK)和激活转录因子(ATF)6。一方面,UPR能够减少蛋白质的合成从而使内质网应激减轻,在有限程度上降低甚至消除蛋白质的折叠给内质网带来的负荷。另一方面,由于前者的调节是有限的,随着蛋白质的过度积累和钙稳态的进一步失衡,UPR将无法修复细胞损伤并引起细胞凋亡,即由内质网应激引起的细胞凋亡。另外,细胞还可启动内质网相关蛋白质降解途径内质网相关蛋白降解(ERAD)从而清除折叠错误的蛋白质〔14〕,然而持续的内质网应激可以通过 UPR 激活凋亡信号通路,这可能与阿尔茨海默病等神经退行性疾病的发生机制有关,并且已被实验证实〔15~18〕。当UPR不足以使细胞对抗应激时,CCAAT/增强子结合蛋白同源蛋白(CHOP)等内质网相关的促凋亡因子表达增加caspase12活化并激活下游caspase3,最终导致凋亡发生。且GO富集显示,差异表达基因主要与膜磷脂的代谢过程相关,提示内质网可能出现了损伤。在本实验中,慢性酒精中毒经过BNP治疗以后Sec61γ的表达又被反向调控到原始水平,可能在一定程度上修复了易位通道,促使新生肽链的转入和错误折叠的蛋白质的转出,减轻UPR,减少神经细胞的死亡,qPCR实验进一步验证了这种结果。
此外,内质网稳态的破坏,会阻碍钙离子的摄取和转运,进而影响线粒体的功能〔19,20〕。过度的内质网应激也可能涉及线粒体和细胞色素C的协同作用,从而激活caspase和细胞凋亡。而在本研究中,芯片数据显示慢性酒精中毒以后Cox6c表达上调,提示线粒体复合体Ⅰ的损伤可能与UPR有关。据Rizzuto等〔21〕的研究,内质网和线粒体之间存在很多密切关系,它们可以通过膜交换钙离子、脂质和糖化蛋白。内质网1,4,5-三磷酸肌醇(InsP3)/Ca2+通道的开放能影响到线粒体中钙离子的平衡,也是caspase3作用的靶标之一〔22,23〕。由此可见,慢性酒精中毒会导致内质网和线粒体的损害。
通过KEGG聚类分析显示,慢性酒精中毒可能与氧化磷酸化通路、阿尔茨海默病信号通路、帕金森病信号通路、亨廷顿病信号通路、D-谷氨酰胺和谷氨酸代谢通路相关。提示慢性酒精中毒损害可能是阿尔茨海默病、亨廷顿病、帕金森病等神经退行性疾病发病的重要因素之一〔24,25〕。慢性酒精中毒可能对海马的神经元造成损伤,进一步损害其学习记忆能力。通过上调Cox6c和Ndufb8,下调Ndufa2、Ndufa4、Ndufb6等的表达,从而抑制线粒体复合物Ⅰ的功能。不仅使机体产生的游离自由基增加,损伤神经细胞的正常功能,而且还能让线粒体中的电子传递链活性被抑制,导致三磷酸腺苷(ATP)产生减少和细胞损伤〔26〕。此外,Cox6c是细胞色素C氧化酶的亚基〔27〕,在本研究中异常上调,也提示着内质网应激可能涉及线粒体及细胞色素C的协同作用而使caspase 激活和细胞凋亡。然而经过BNP治疗以后,Ndufa4,Ndufa2又被上调到正常水平,qPCR实验进一步验证了这一结果。提示BNP对线粒体具有一定的保护作用,可在一定程度上,对慢性酒精中毒起到治疗作用。利用生物信息学的方法能有效地分析基因芯片数据,从而高效地获取生物内在信息〔28〕,本研究中差异表达基因涉及蛋白酶体通路、氧化磷酸化通路等通路,可能与内质网内积聚的未折叠或错误折叠的蛋白的降解及线粒体复合体的修复有关,可能通过这些通路对线粒体及内质网起到一定的保护作用。综上,通过生物信息学的分析发现BNP可以将由慢性酒精中毒造成差异表达的多个基因反向调控到正常水平,通过调控Sec61G、Ndufa4、Ndufa2等内质网或线粒体相关基因的表达,缓解了内质网应激,减轻线粒体损伤,在一定程度上保护了神经元细胞。BNP对慢性酒精中毒引起的海马体损伤具有一定的保护作用。
