菲律宾蛤仔GnRH基因沉默对其性激素合成酶基因表达的影响❋
2020-04-24姜珊珊苗晶晶刘力铷潘鲁青
姜珊珊, 苗晶晶, 刘力铷, 潘鲁青
(中国海洋大学水产学院水产动物环境生理学研究室,山东 青岛 266003)
促性腺激素释放激素(Gonadotropin-releasing hormone,GnRH)是一种神经肽类激素,是性激素合成与性腺发育的关键内分泌调控因子[1]。在鱼类和其它脊椎动物中,GnRH通过刺激垂体促性腺激素(Gonadotropin, GtH)的产生从而调节性腺类固醇激素的合成与释放,促进性腺发育成熟[2],然而目前对贝类GnRH的调控机制了解甚少。近年来,在几种贝类体内均发现了GnRH多肽的存在,GnRH基因在虾夷扇贝(Patinopectenyessoensis)和长牡蛎(Crassostreagigas)的神经组织中高度表达,且与脊椎动物GnRH基因相比具有较高的同源性和结构相似性[3-5]。此外,Rodet等在长牡蛎体内克隆到了GnRH受体基因,并发现该基因主要在雌雄性腺中表达[6]。Treen等研究发现GnRH多肽可刺激体外培养的虾夷扇贝精原细胞分裂[7]。目前在软体动物中尚未发现GtH同源基因的存在,Kah等[8]推测GnRH的作用方式可能随动物神经系统的进化而演变,在神经系统较为简单的无脊椎动物中,GnRH可能通过直接作用于性腺发挥其生殖调控作用,目前这一观点仍有待于进一步证实。
脊椎动物通过“GnRH-GTHs-性激素合成通路”调控性腺发育,性激素由孕烯醇酮通过Δ4和Δ5两种主要途径合成,这两条途径都涉及细胞色素P450c17(CYP17)以及3β-羟类固醇脱氢酶(3β-hydroxysteroid dehydrogenase,3β-HSD)和17β-羟基类固醇脱氢酶(17β-hydroxysteroid dehydrogenase, 17β-HSD)[9]。在双壳动物体内,性激素(雌二醇、睾酮和孕酮等)已经被广泛的鉴定和量化,许多研究通过对双壳贝类性腺组织中类固醇激素代谢、生化变化的研究,提出双壳贝类体内性激素的主要合成途径与脊椎动物类似,且类固醇合成酶对性激素的生物合成至关重要,其中,CYP17、3β-HSD和17β-HSD是关键的性激素合成酶[10-13]。脊椎动物中CYP17、3β-HSD和17β-HSD在转录水平由GnRH、GtH通过其受体激活。然而在软体动物中这3种酶基因表达调控的相关研究尚未见报道。本文在前期克隆了菲律宾蛤仔(Ruditapesphili-ppinarum)rp-GnRH全长cDNA序列的基础上,设计并筛选了特异性干扰GnRHmRNA表达的dsRNA,旨在通过RNAi技术,研究菲律宾蛤仔rp-GnRH基因沉默对性腺中性激素合成相关基因表达的影响,进而探讨GnRH在菲律宾蛤仔生殖内分泌调控中的作用。
1 材料与方法
1.1 dsRNA制备
采用体外合成法制备GnRH及绿色荧光蛋白(Green fluorescent protein)的dsRNA用于干扰实验。根据菲律宾蛤仔GnRH基因序列信息(GenBank KF891317)和RNAi引物设计原则设计了两组不同的引物,并分别在每组引物的正义链和反义链的5’末端添加T7序列(见表1),以蛤仔脏神经节cDNA为模板,PCR扩增特异序列。根据绿色荧光蛋白(EGFP-C1,Clontech)序列信息设计RNAi引物,用以从EGFP-C1中扩增特异序列。
用TIANgel Midi Purification Kit(TIANGEN)纯化PCR产物,按照T7 RiboMAXTM Express RNAi System(Promega)试剂盒说明书的方法,体外合成用于干扰GnRH表达的dsRNA及EGFP-dsRNA(阴性对照),并用不含RNase的DNaseⅠ(Takara)消化模板DNA。将获得的 dsRNA 以1%琼脂糖凝胶电泳分析其大小和纯度,用核酸蛋白仪(Ultrospec 2100)测定合成的dsRNA 浓度和OD值,OD260/OD280为1.84~2.16。

表1 RNAi dsRNA合成引物
1.2 GnRH-dsRNA筛选及不同时间作用下的干扰效率
菲律宾蛤仔于2017年11月采自青岛市红岛养殖场,壳长(3.4±0.3) cm,性腺发育处于休止期。实验前于实验室暂养一周,定期换水和投喂螺旋藻粉。将菲律宾蛤仔随机分为4组:空白对照组(PBS组)、阴性对照组(EGFP-dsRNA组)和2个GnRH干扰组(GnRH-ds1组、GnRH-ds2组),每组3个平行,每个平行注射3个个体。将体外转录合成的dsRNA 溶解于灭菌的 PBS 缓冲液中,用注射器从菲律宾蛤仔双壳铰合部注入血腔中,其中空白对照组每个个体注射50 μL灭菌的PBS,阴性对照组和2个GnRH干扰组每个个体分别注射50 μL(10 μg/50 μL)的EGFP-dsRNA、GnRH-ds1和GnRH-ds2。参考王晓安等[14],菲律宾蛤仔脏神经节位于后闭壳肌膜上,在注射后的0、24、48和72 h用镊子取脏神经节提取总RNA,用RT-qPCR法测定GnRH的表达水平,计算干扰率。
1.3 不同浓度GnRH-dsRNA的干扰率
挑选36只健康的蛤仔,随机分为4组,根据GnRH基因干扰链筛选结果,参考合浦珠母贝(Pinctadafucata)[15]目的基因dsRNA的浓度设定,设定空白对照组(注射50 μL灭菌的PBS)、阴性对照组(注射50 μL 20 μg/50 μLEGFP-dsRNA)、GnRH-ds1低浓度组(注射50 μL20 μg/50 μL GnRH-ds1)和GnRH-ds1高浓度组(注射 50 μL 30 μg/50 μLGnRH-ds1),于48 h后用镊子取脏神经节提取总RNA,用RT-qPCR法测定GnRH的表达水平,计算干扰率。
1.4 GnRH干扰对性腺发育相关基因的表达研究
将菲律宾蛤仔随机分为3组,包括空白对照组(PBS)、阴性对照组(EGFP-dsRNA)和GnRH-ds1组。空白对照组每个个体注射50 μL灭菌的PBS缓冲液,阴性对照组每个个体注射50 μLEGFP-dsRNA,RNAi组每个个体分别注射50μL筛选出的30 μg/50 μLGnRH-ds1。于48 h后取性腺组织提取总RNA,用RT-qPCR法测定性激素合成相关基因的表达。
1.5 基因表达的RT-qPCR分析
根据RNAiso Plus(TaKaRa,大连)说明书分别提取脏神经节、性腺总RNA。每一样品取1 μg总RNA,按照 PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa,大连)试剂盒说明书反转录合成cDNA,以cDNA为模版,进行实时荧光定量PCR反应用于检测rp-GnRH及性激素合成相关基因表达量, 所用引物见表2。以18s rRNA为内参基因,荧光定量数据分析采用 2-ΔΔCt 法。用SPSS 22.0软件进行单因素方差分析(One-Way ANOVA),采用Tukey’s多重比较检验差异显著性,以p<0.05作为差异显著标准。
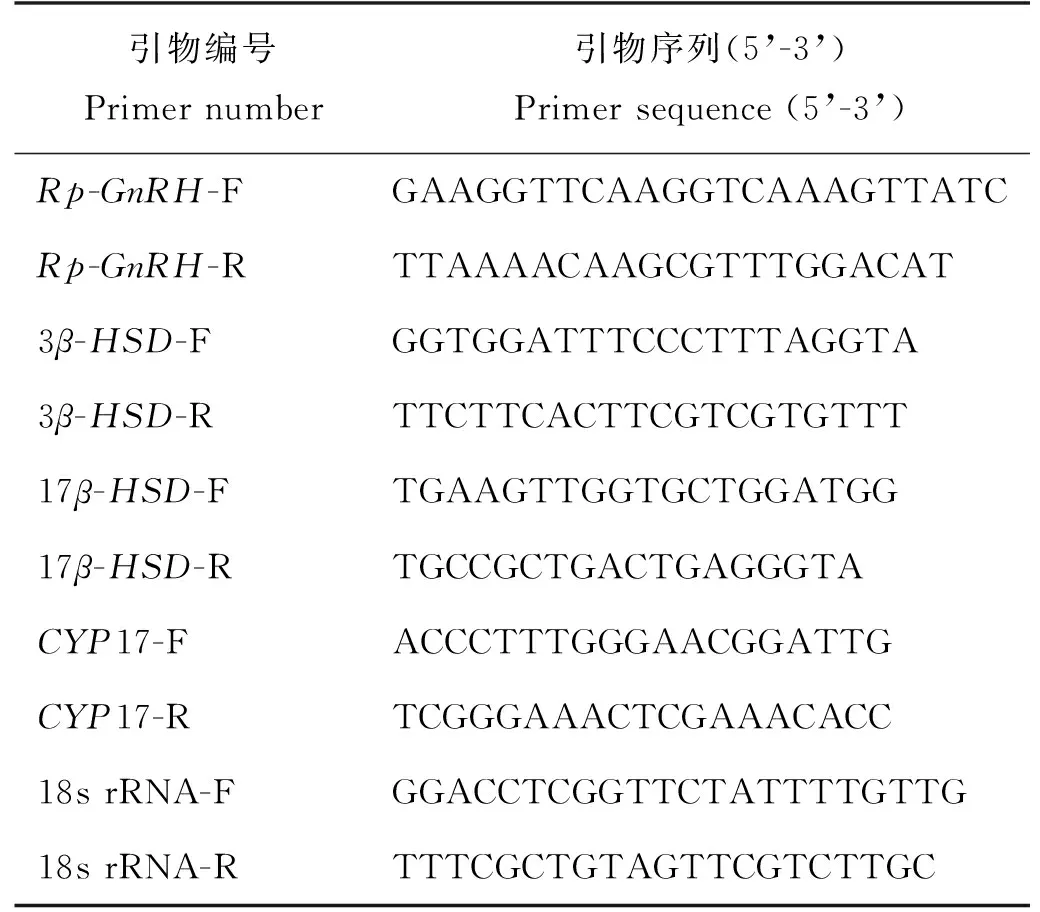
表2 荧光定量引物
2 实验结果
2.1 两种 GnRH-dsRNA干扰的效果检测
注射后的 0、24、48和72 h,提取脏神经节检测GnRH的相对表达量。结果表明, PBS组与阴性对照组(EGFP-dsRNA)相比,GnRH的表达量无显著性差异(p>0.05)。在注射后48 h,两个干扰组GnRH的表达量出现显著下降(p<0.05),其中GnRH-ds1注射组GnRH表达量干扰率为63.3%,GnRH-ds2注射组干扰效率为38.9%(见图1)。
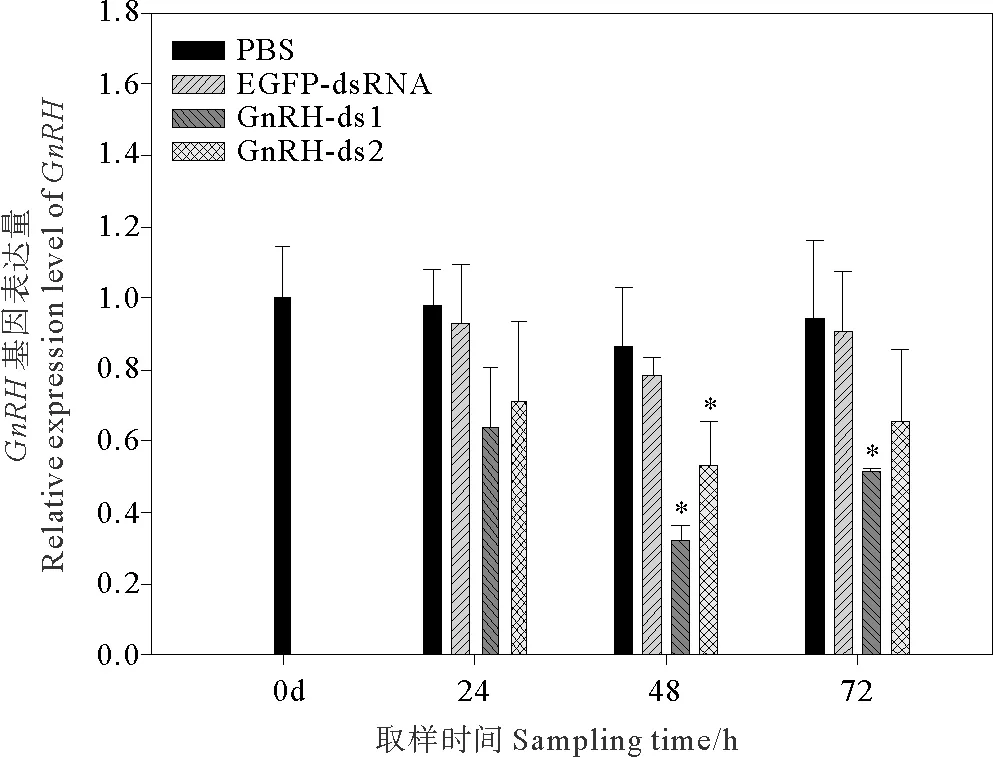
图1 dsRNA注射对菲律宾蛤仔脏神经节GnRH基因表达的影响
2.2 不同浓度GnRH-ds1的RNAi效果
根据2.1中GnRH的干扰效果,选择GnRH-ds1进行注射,设置浓度梯度为20 μg/50 μL和30 μg/50 μL,于48 h取样检测抑制效果。结果表明,注射后48 h,PBS组与阴性对照组相比,GnRH基因表达量无显著性差异(p>0.05)。高浓度(30 μg/50 μL)GnRH-ds1注射组的干扰率为73.3%,低浓度(20 μg/50 μL)注射组的干扰率为54.3%(见图2)。

图2 不同浓度dsRNA注射对菲律宾蛤仔脏神经节GnRH基因表达的影响
2.3 GnRH基因干扰后性激素合成基因表达量的变化
如图3所示,注射dsRNA 48 h后,GnRH干扰组的菲律宾蛤仔性腺中17β-HSD和CYP17的基因表达较空白对照组、阴性对照组显著性下降(p<0.05),而3β-HSD基因表达量没有出现显著变化(p>0.05)。与PBS对照组相比,GnRH干扰组中17β-HSD及CYP17基因的表达量分别下降了86.3%、84.9%;与阴性对照组相比,17β-HSD及CYP17基因表达量分别下降了78.6%、81.3%。
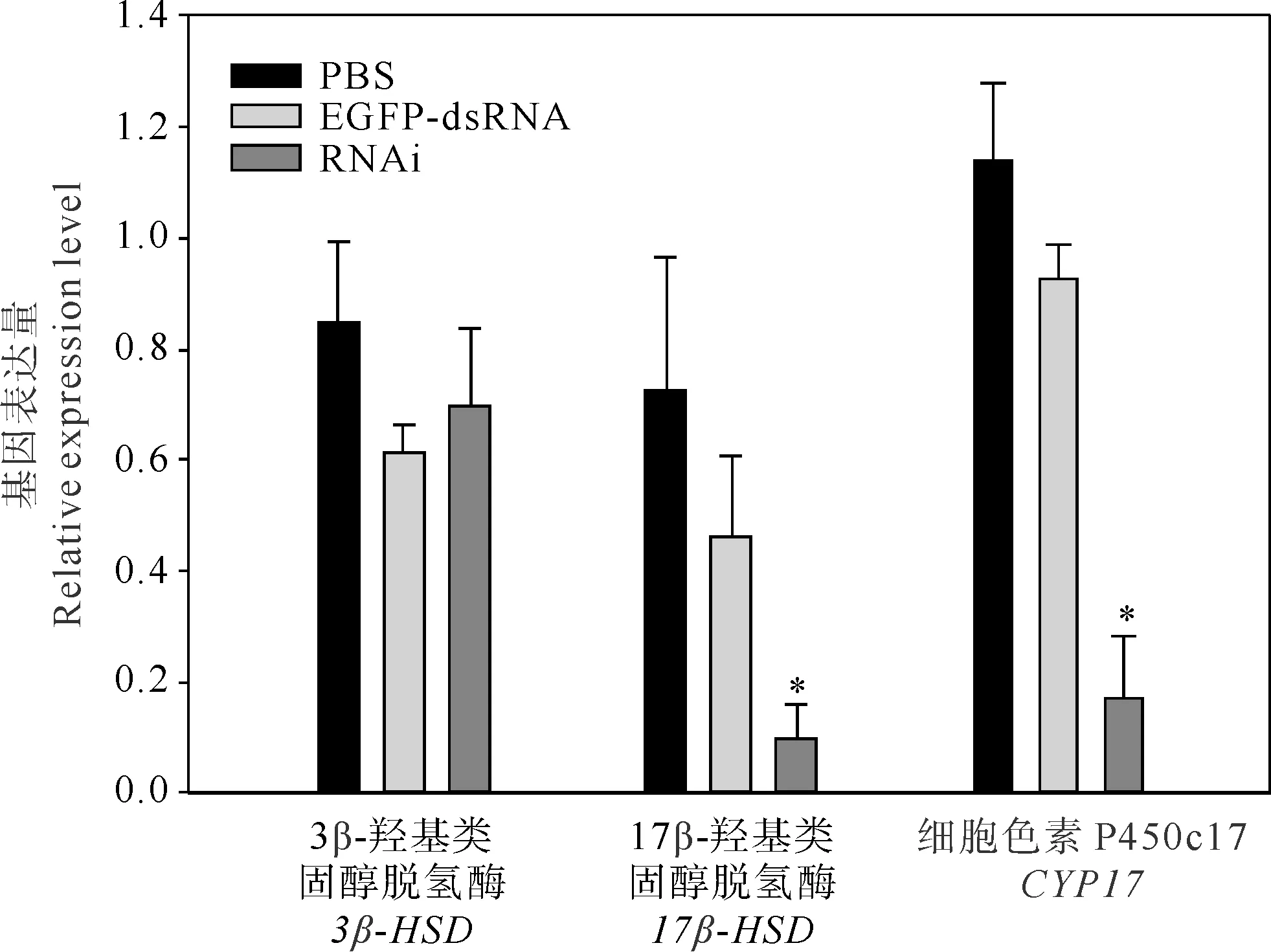
图3 不同处理组3β-HSD、17β-HSD和CYP17基因的相对表达量
3 讨论
目前,RNAi作为一种简单、高效、特异性沉默同源目标基因的敲减工具, 为动物基因功能的研究提供了新的思路和方法。Fabioux等体外合成了牡蛎性腺基因Oyvlg的RNA干扰链(Oyvlg-dsRNA),将其注射到牡蛎性腺中成功引起不育现象,首次证明双壳贝类体内注射dsRNA的有效性[16]。这项技术在日本合浦珠母贝、三角帆蚌、栉孔扇贝等双壳贝类中也均有应用报道[15,17-18]。RNAi在生物体内具有一定的时效性,多数研究表明RNAi持续的时间为2~4 d。对甲壳动物的研究发现在南美白对虾(Litopenaeusvannamei)和克氏原螯虾(Procambarusclarkii)中,注射目的基因dsRNA干扰链后48 h即可达到有效干扰效果[19-20]。Feng等探究栉孔扇贝Cf-dmrt-likemRNA基因功能时dsRNA注射后72 hCf-dmrt-like表达量显著性降低[18]。本研究结果显示,GnRH-ds1干扰链注射后48 h显著下降,在注射后72 h,GnRH基因的表达量开始回升(见图1)。在RNAi实验中,dsRNA处理后目的基因mRNA表达水平下降70%被认为干扰有效[21]。已有研究表明在RNAi实验中dsRNA的有效注射浓度有所区别:如李颖翔等设计的长牡蛎Toll样受体(Toll-like receptor)的dsRNA的有效注射浓度为100 μg/100 μL[22]。Fang等以合浦珠母贝为研究对象,设计了3种不同浓度的dsRNA用以抑制5个与贝壳矿化有关的基因的表达,结果显示高浓度组(160 μg/100 μL dsRNA)对目的基因的抑制率显著高于低浓度组(40 μg/100 μL或80 μg/100 μL)[15]。本研究结果显示向菲律宾蛤仔血腔中注射10 μg/50 μLGnRH-ds1和ds2即可抑制GnRH的表达(抑制率分别为63.3%和38.9%),但尚未达到有效抑制率,而注射30 μg/50 μLGnRH-ds1 48 h后GnRH的抑制率达73.31%, 说明脏神经节中GnRH的mRNA表达被有效抑制。由此可见,本实验中对GnRH基因的干扰效果也主要受到dsRNA注射浓度的影响有关,30 μg/50 μL为GnRH-ds1注射48 h的有效浓度。
脊椎动物中促性腺激素释放激素(GnRH)是性激素合成与性腺发育的关键神经内分泌调控因子。对硬骨鱼类的研究发现,体内注射GnRH短期内可诱导垂体GTHs合成,同时注射GnRH和多巴胺拮抗剂可促进多数种类的性腺发育成熟[23-24]。目前关于软体动物GnRH功能的研究较少:Lisa等研究发现人工合成的真蛸(Octopusvulgaris)GnRH多肽可促进离体培养的真蛸卵巢和精巢组织中孕酮、睾酮和雌二醇含量显著升高[25-26];Nakamura等采用人工合成的GnRH离体孵育虾夷扇贝(P.yessoensis)的精巢,发现可促进精原细胞分裂[27];Song等发现菲律宾蛤仔脏神经节rpGnRH基因表达在性腺发育过程中具有明显的变化规律,其相对表达量与血液中孕酮、睾酮的含量具有显著正相关性[5]。这些研究结果均体现了GnRH在软体动物的性腺发育与性激素合成中可能具有重要作用。脊椎动物中性激素合成关键酶CYP17、3β-HSD及17β-HSD等基因在转录水平受到GnRH和GtHs的调控[10],但是贝类中GnRH对性激素合成酶基因表达的影响尚未见报道。在本实验中,处于性腺发育休止期的菲律宾蛤仔的GnRH表达被抑制时,两种关键的性激素合成酶CYP17与17β-HSD的基因表达均显著下调,间接证实了双壳贝类中GnRH对其性激素合成的调控作用。另一方面,有研究表明,菲律宾蛤仔在性腺发育时期血淋巴中性激素水平(孕酮、睾酮和雌二醇)的含量及变化均存在雌雄差异[5],但是GnRH功能在软体动物中的性别差异研究尚未见报道。本实验中的受试菲律宾蛤仔处于性腺发育休止期,因此在本研究中未区分GnRH表达抑制对不同性别菲律宾蛤仔的影响,在今后的实验中将测试GnRH表达抑制对不同发育阶段、不同性别的菲律宾蛤仔性激素合成、主要性激素含量的影响,为查明GnRH在不同发育阶段和不同性别菲律宾蛤仔中的功能提供依据。
目前研究发现贝类体内的性激素合成途径与脊椎动物类似,主要由CYP17、3β和17β-HSDs等性激素合成关键酶介导完成[11-13]。Thitiphuree等研究表明CYP17、3β-HSD和17β-HSD在虾夷扇贝的外周和性腺组织中普遍表达,并且3β-HSD和17β-HSD基因的mRNA表达与性腺成熟度有关[28]。CYP17同时具有17α-羟化酶和17,20-裂解酶的两种活性,参与脊椎动物中孕烯醇酮和孕酮的17-羟化、催化17-羟孕酮转化为雄烯二酮和17-羟孕烯醇酮转化为去氢表雄酮(DHEA)。Mu等研究许氏平鲉发现,性腺中CYP17a1的表达与血清中睾酮浓度密切相关[29]。研究表明在斑点叉尾鮰(Ictaluruspunctatus)和日本鳗鲡(Anguillajaponica)卵泡发育过程中,CYP17的含量变化极为显著[30-31]。早在1973年,De等利用使用氚和氧化标记的前体,在紫贻贝中发现了CYP17酶活性,随后Carreau等利用同样的方法在乌贼(SepiaofficinalisL.)性腺中也发现CYP17酶活性[32]。Thitiphuree等在虾夷扇贝性腺中克隆到了一种CYP17的cDNA全长,与脊椎动物氨基酸序列相比具有较高的保守性,并且虾夷扇贝CYP17mRNA的表达水平在配子早期分化阶段较高,在性腺成熟中期、后期受到抑制,说明CYP17在贝类的性腺发育过程中具有重要的作用[28]。在脊椎动物中,17β-HSD主要功能是参与性激素的代谢,通过调节细胞内甾体类激素水平从而在生殖系统中发挥重要作用。它可以将脱氢表雄酮转化为雄烯二酮, 并能将雌酮转化为雌二醇。对贝类的研究表明,长牡蛎和虾夷扇贝的雌性性腺中均可检测到雌二醇和17β-HSD酶活性,且雌二醇的含量随性腺成熟升高,而排卵后17β-HSD的酶活性与早期性腺分化阶段相比明显降低[33]。Liu等通过实验证明栉孔扇贝中的17β-HSD 能有效地将17β-雌二醇转化为雌酮,并可催化睾酮向雄烯二酮的转化。此外,栉孔扇贝雌雄性腺17β-HSD的表达量随配子发育而增加,在生长期和成熟期雄性性腺17β-HSD的表达量显著高于雌性性腺[34]。Zhou等研究证实九孔鲍(Haliotisdiversicolorsupertexta)中17β-HSD可以催化雌酮转化为雌二醇,并且在成熟期性腺内17β-HSDmRNA表达量显著升高,在产卵后显著性下降[35]。这些研究表明17β-HSD在贝类性类固醇激素的合成中发挥重要作用,进而调节贝类性腺发育。本研究结果显示,当GnRH表达被抑制时,CYP17、17β-HSDmRNA的表达均显著下降,由此我们推测在性腺发育处于休止期的菲律宾蛤仔中,CYP17、17β-HSD基因表达可能在转录水平受到GnRH的调控。
3β-HSD作为动物体内大多数性激素生物合成的关键酶,在睾酮生物合成中起限速酶的作用。Thitiphuree等通过RACE技术获得了虾夷扇贝(Mizuhopectenyessoensis)性腺组织3β-HSD的cDNA全长,研究发现当扇贝性成熟出现大量生殖细胞时,3β-HSD的表达量最高[28],说明3β-HSD在贝类性腺发育中也可能具有重要作用。Lin等通过GnRH体外培养大鼠睾丸间质细胞发现,GnRH可促进细胞中3β-HSD的表达进而促进睾酮的分泌[36]。Chianese等研究发现蛙(Rana esculenta)体内内源性大麻素通过与大麻素受体(Cannabinoidreceptor,CB1)结合抑制下丘脑GnRH分泌及GtH的释放从而降低了蛙睾丸中3β-HSD的表达进而影响睾酮的生成[37]。本研究结果显示当GnRH被干扰时,菲律宾蛤仔性腺内3β-HSD的表达并没有发生显著变化。在脊椎动物中,3β-HSD可能受多种其他因素调节,如卵巢内ghrelin的表达参与抑制3β-HSD活性,降低性激素生成[38]。此外,3β-HSD主要表达于精巢间质细胞、精原细胞、颗粒细胞以及精子中参与睾酮的合成,其中精巢间质细胞是雄激素合成的主要场所,且仅在减数分裂时期发挥上述功能[39]。由此,我们推测菲律宾蛤仔雌雄性腺内3β-HSD的表达调控可能与其所处的性腺发育时期有关。
