丙二醛氧化修饰对白鲢肌原纤维蛋白 结构性质的影响
2020-04-02蒋祎人刘友明熊善柏
蒋祎人,李 涛,刘友明,,熊善柏
(1.华中农业大学食品科学技术学院,湖北 武汉 430070;2.国家大宗淡水鱼加工技术研发分中心(武汉),湖北 武汉 430070)
白鲢(Hypophthalmichthys molitrix)作为我国重要的养殖淡水鱼品种,是淡水鱼糜加工的主要鱼种。白鲢鱼肉中富含营养价值较高的不饱和脂肪酸,鱼糜漂洗过程中未能完全去除鱼肉中的脂肪,导致鱼糜在冻藏过程中易发生脂肪氧化,同时脂肪氧化过程中生成的自由基及次级产物均会引发蛋白质氧化,从而引起蛋白质的功能特性发生改变[1]。
丙二醛是脂肪氧化主要的次级产物之一,其含量可用来衡量脂肪氧化的程度。丙二醛不仅可以氧化蛋白质的侧链和多肽骨架,造成蛋白质空间结构的改变,从而改变其功能特性,还可以与氨基酸基团反应,生成席夫碱导致分子间聚合物的生成[2-5]。已研究表明,较为温和的氧化条件可适当提高植物蛋白的功能性质,但氧化过度会导致功能特性下降[6-7]。动物蛋白经过丙二醛氧化改性后,二级结构发生改变。一般来说,氧化程度增加会导致蛋白质的α-螺旋含量下降,无规卷曲含量上升[8]。 白鲢肌原纤维蛋白经丙二醛诱导氧化后蛋白的溶解性下降,而且氧化程度增加可使白鲢鱼糜流动性降低,无法进行挤压式3D打印[9]。适宜浓度的丙二醛可引起蛋白质发生氧化,产生共价交联,从而促进蛋白质热凝胶形成能力、凝胶强度和凝胶持水性增强[10]。目前从脂肪氧化和蛋白质氧化的角度阐述白鲢鱼糜品质变化规律的研究较少。研究氧化应激环境中白鲢肌原纤维蛋白的响应机制可有助于理解白鲢鱼糜在冻藏过程中品质变化的分子机理。本实验以低浓度的丙二醛溶液作为氧化体系,对白鲢肌原纤维蛋白进行氧化,通过分析氧化前后白鲢肌原纤维蛋白结构特性变化,探究脂质氧化次级产物中的小分子醛类对鱼肉主要蛋白质结构的影响规律,以期为鱼糜冻藏过程中品质变化的量化检测提供理论依据。
1 材料与方法
1.1 材料与试剂
新鲜白鲢,体质量2~3 kg/条,春季购于华中农业大学农贸市场,冰水保鲜快速运至实验室,清洗干净后去除头尾和内脏,取背部肌肉用于提取肌原纤维蛋白。
氯化钠、磷酸氢钠、磷酸氢二钠、尿素、三氯乙酸、 乙二胺四乙酸、 2 , 4 - 二硝基苯肼(2,4-dinitrophenylhydrazine,DNPH)、二硝基苯甲酸(5,5’-dithiobis(2-nitrobenzoic acid),DTNB)(均为分 析纯) 国药集团化学试剂有限公司;1,1,3,3-四甲氧基丙烷(1,1,3,3-tetramethoxypropane,TMP)、三羟甲基氨基甲烷(均为分析纯) 美国Sigma公司。
1.2 仪器与设备
K600食品料理仪 德国博朗电器公司;722N型可见分光光度计 上海仪电分析仪器有限公司;Avanti J-26XP冷冻离心机 美国贝克曼公司;FJ-200高速分散均质机 上海标本模型厂;F-4600荧光分光光度计 日本岛津公司;Bio-Rad电泳仪 美国博乐公司;J-1500圆二色谱仪 日本JASCO公司。
1.3 方法
1.3.1 肌原纤维蛋白的提取
参考Huang Jingjing等[11]的方法提取白鲢肌原纤维蛋白并略作改动。从新鲜宰杀的白鲢背部采集鱼肉,然后用4 倍体积的低盐磷酸缓冲液(20 mmol/L、0.05 mol/L NaCl、pH 7.5)漂洗3 次,得到的沉淀与4 倍体积的高盐磷酸缓冲液(20 mmol/L、0.45 mol/L NaCl、pH 7.5)混合均匀,4 ℃静提过夜(22 h)。将混合物进行离心(12 000 r/min、10 min、4 ℃),取上清液,加入10 倍体积的冷凝水,静置30 min后离心(10 000 r/min、10 min、4 ℃),除去上清液,得到的沉淀即为肌原纤维蛋白,于4 ℃存放备用。用福林-酚法测量蛋白浓度[12]。
1.3.2 丙二醛溶液的制备
参考Wang Lin等[13]的方法配制丙二醛溶液,并略作改动。将8.4 mL TMP溶于10 mL 5 mol/L盐酸中用纯水定容至50 mL后,将溶液置于40 ℃水浴锅中避光水浴振荡30 min,直至深黄色生成。然后用6 mol/L NaOH调溶液pH值为6.0,最后将丙二醛原储液用50 mmol/L磷酸盐缓冲液(pH 6.0)定容至250 mL。丙二醛浓度用紫外分光光度计测定:将丙二醛溶液稀释105倍后在波长267 nm(碱液,ε=31 500)处测定吸光度,从而计算浓度。将丙二醛储备液避光低温保存。
1.3.3 肌原纤维蛋白的氧化
向质量浓度为40 mg/mL的肌原纤维蛋白溶液中加入不同浓度的丙二醛(0、0.05、0.10、0.25、0.50、1.00 mmol/L),在25 ℃条件下避光反应24 h,得到不同氧化程度的肌原纤维蛋白样品。
1.3.4 羰基含量的测定
参考Mesquita等[14]的方法,略作改动。400 μL 10 mmol/L DNPH(含有0.5 mol/L H3PO4)加入400 μL蛋白质溶液(2 mg/mL)混合均匀。混合液室温反应10 min后,加入200 μL NaOH溶液(6 mol/L),继续在室温下反应10 min,于450 nm波长处测定吸光度,以消光系数22 308 L/(mol·cm)计算蛋白羰基衍生物含量。
1.3.5 二聚酪氨酸含量的测定
参考Davies等[15]的方法进行测定。测量条件:发射波长420 nm,激发波长325 nm,狭缝宽度均为10 nm。二聚酪氨酸含量用相对荧光值表示。
1.3.6 巯基含量的测定
参考Ellman[16]的方法进行测定。测定活性巯基含量时,向1 mL肌原纤维蛋白溶液(2~4 mg/mL)中加入9 mL 0.02 mol/L Tris-HCl缓冲溶液(含2%十二烷基硫酸钠(sodium dodecyl sulfate,SDS)、10 mmol/L EDTA,pH 6.8)和1 mL Ellman试剂(含0.1% DTNB、0.2 mol/L Tris-HCl,pH 6.8),混匀后于4 ℃恒温放置1 h,测其在412 nm波长处的吸光度。
测定总巯基含量时,向1 mL肌原纤维蛋白溶液(2~4 mg/mL)中加入9 mL 0.02 mol/L Tris-HCl缓冲溶液(含8 mol/L尿素、2% SDS、10 mmol/L EDTA,pH 6.8)和1 mL Ellman试剂混合均匀,40 ℃水浴25 min,测其在412 nm波长处的吸光度。
1.3.7 游离氨基含量的测定
参考冯岩等[17]的方法,略作修改。邻苯二甲醛(o-phthaldialdehyde,OPA)试剂的配制:准确称取40 mg OPA溶解于1 mL甲醇中,分别加入2.5 mL 20%的SDS,25 mL 0.1 mol/L四硼酸钠溶液,100 μL β-巯基乙醇,用蒸馏水定容到50 mL,此为OPA试剂,且使用前即时配制。取200 μL蛋白样品(2 mg/mL)加入4 mL OPA试剂,混匀后于35 ℃反应2 min,在340 nm波长处测定吸光度。以L-亮氨酸制作标准曲线,计算样品中游离氨基含量。
1.3.8 表面疏水性的测定
参考Yongsawatdigul等[18]的方法,略作修改。蛋白疏水性采用1-苯胺基-8-萘基磺酸盐(8-anilino-1-naphthalenesulphonic acid,ANS)荧光测定法。用缓冲液(0.6 mol/L NaCl、20 mmol/L Tris-HCl,pH 7.5)将不同氧化程度的肌原纤维蛋白稀释为0.05、0.1、0.15 mg/mL和0.2 mg/mL。取 4 mL稀释的蛋白溶液与 20 μL 8 mmol/L ANS溶液混匀,避光反应10 min后测量荧光强度。测量时激发波长和发射波长分别为390 nm和470 nm。以相对荧光强度对蛋白质浓度的斜率作为表面疏水性。
1.3.9 Ca2+-ATPase活性的测定
参考Benjakul等[19]的方法,略作修改。向3.5 mL肌原纤维蛋白溶液(2 mg/mL)中加入 0.3 mL Trismaleat溶液(0.5 mol/L,pH 7.0),0.5 mL CaCl2溶液(0.1 mol/L),25 ℃水浴恒温后,加入0.25 mL 20 mmol/L ATP溶液,在25 ℃反应10 min后用2.5 mL 15%三氯乙酸阻断反应。反应液离心后(6 5 0 0 r/m i n、5 min)得上清液。空白组中三氯乙酸和ATP添加顺序调换。采用钼酸铵法测定反应释放出无机磷含量, Ca2+-ATPase活性以1 mg蛋白在1 min内生成的无机磷含量 计,以μmol/(mg·min)表示。
1.3.10 溶解性的测定
取5 mL蛋白样品离心(8 000×g、10 min、4 ℃),采用福林-酚法[12]测定上清液的蛋白质浓度。溶解度表示为离心后的蛋白质浓度与离心前蛋白质浓度的百分比。
1.3.11 SDS-聚丙烯酰胺凝胶电泳(SDS-polyacrylamide gel electrophoresis,SDS-PAGE)
参考Xiong Youling等[20]的方法,略作修改。不同氧化程度的肌原纤维蛋白与含有(不含有)5% β-ME的样品缓冲液(4% SDS、10% β-ME、2%甘油、0.5%溴酚蓝,pH 6.8)混合,将蛋白质量浓度调至1.0 mg/mL。用10%分离胶,4%浓缩胶。电极缓冲液为Tris-甘氨酸缓冲液(含0.1% SDS,pH 8.3),用考马斯亮蓝R-250染色液(含0.15%考马斯亮蓝R-250、50%乙醇、10%冰醋酸)进行染色;用含50%乙醇、10%冰醋酸的洗脱液进行脱色。
1.3.12 圆二色谱测度
参考Wang Zhaoming等[21]的方法,略作修改。将肌原纤维蛋白溶液样品稀释至1 mg/mL,放入0.1 cm光径的石英样品池中,在远紫外区(波长范围190~250 nm)进行扫描,扫描速率为100 nm/min,测定温度为20 ℃。残基平均摩尔质量为110 g/mol,采用杨氏模量模拟计算α-螺旋、β-折叠、β-转角和无规卷曲二级结构相对含量。圆二色性用平均残基椭圆值θ表示,单位 为(deg·cm2)/dmol。
1.4 数据处理
采用Excel软件进行数据分析,Origin 8.0和Excel软件作图,运用SPSS 19.0统计软件进行相关性及显著性分析(P<0.05,显著水平)。
2 结果与分析
2.1 丙二醛对白鲢肌原纤维蛋白羰基含量的影响
如图1所示,经丙二醛氧化后的肌原纤维蛋白中羰基的含量显著高于未添加的对照组(P<0.05),说明丙二醛氧化导致肌原纤维蛋白的羰基含量增加。低浓度的丙二醛(0.05~0.50 mmol/L)浓度变化与肌原纤维蛋白羰基含量之间的量效关系不显著(P≥0.05),而高浓度丙二醛(1.00 mmol/L)使羰基含量显著增加(P<0.05),此时蛋白质氧化程度明显增加。羰基的主要产生途径是敏感性氨基酸侧链的直接氧化以及肽键的断裂[22],实验中丙二醛浓度在0.05~0.50 mmol/L范围内羰基含量的变化不显著,表明此浓度范围内蛋白质发生的氧化未大量攻击敏感性氨基酸侧链和肽键,属于轻度氧化。

图 1 丙二醛浓度对白鲢肌原纤维蛋白羰基含量的影响Fig. 1 Effect of MDA concentration on the carbonyl content of myofibrillar protein from silver carp
2.2 丙二醛对白鲢肌原纤维蛋白二聚酪氨酸含量的影响

图 2 丙二醛浓度对肌原纤维蛋白二聚酪氨酸含量的影响Fig. 2 Effect of MDA concentration on dityrosine content of myofibrillar protein from silver carp
由图2可知,在较低浓度丙二醛(≤0.25 mmol/L)氧化应激环境中,白鲢肌原纤维蛋白的二聚酪氨酸含量无显著性变化(P≥0.05),丙二醛浓度达0.50 mmol/L时,二聚酪氨酸含量显著增加(P<0.05)。这可能是在低浓度的丙二醛条件下,蛋白氧化程度较轻,蛋白空间结构比较完整和稳定,丙二醛不易攻击氨基酸侧链,产生的酪氨酰自由基和酪氨酸残基数量较少[23];当丙二醛浓度增加到0.50 mmol/L,氨基酸侧链遭到攻击的机会增多,酪氨酸单体被氧化的几率更大,迅速与其周围其他蛋白的酪氨酸残基产生了共价交联,因此形成了较多的酪氨酸二聚体[24]。
2.3 丙二醛对白鲢肌原纤维蛋白总巯基和活性巯基含量的影响
如图3 所示,总巯基含量在丙二醛浓度较低(≤0.25 mmol/L)时没有显著性变化,随着浓度进一步增加,总巯基含量开始下降。活性巯基的含量呈现先上升后下降的量效关系,丙二醛浓度为0.25 mmol/L时活性巯基的含量增加至1.02 μmol/g。蛋白质发生轻度氧化时,分子结构展开,构象发生变化,原本包埋在内部的巯基暴露出来使检测到的活性巯基含量上升,但丙二醛浓度高于0.25 mmol/L后总巯基和活性巯基含量均有所下降,可能是由于暴露的巯基基团遭到氧化攻击,形成了磺酸或二硫键等相关化合物[25]。活性巯基含量与ATPase活性、疏水相互作用和胶凝过程中二硫键的形成有密切关系,在鱼糜热凝胶形成过程中,肌原纤维蛋白氧化后活性巯基含量的上升可促进鱼糜凝胶强度的增加[26]。
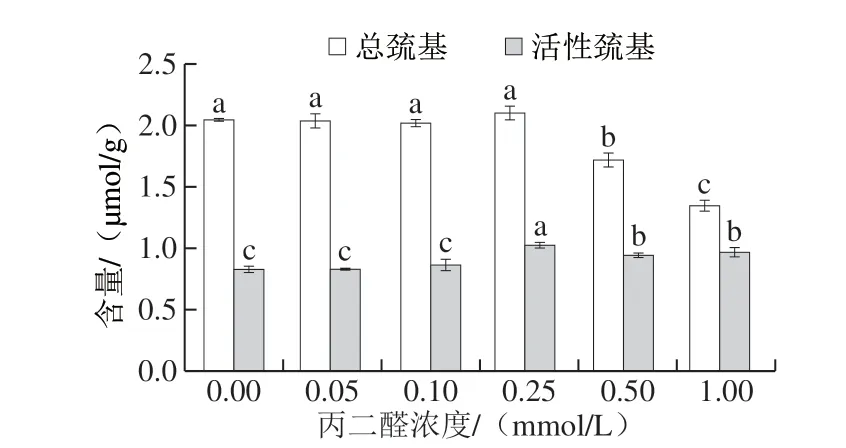
图 3 丙二醛浓度对肌原纤维蛋白总巯基和活性巯基含量的影响Fig. 3 Effect of MDA concentration on total thiol and active thiol contents of myofibrillar protein from silver carp
2.4 丙二醛对白鲢肌原纤维蛋白游离氨基含量的影响

图 4 丙二醛浓度对肌原纤维蛋白游离氨基含量的影响Fig. 4 Effect of MDA concentration on free amino group content of myofibrillar protein from silver carp
如图4所示,丙二醛浓度低于0.50 mmol/L时游离氨基含量无显著差异(P≥0.05),丙二醛浓度达到1.00 mmol/L时游离氨基显著降低(P<0.05)。蛋白发生氧化时,氨基酸侧链中的NH—或NH2基团可能会受到自由基攻击产生羰基衍生物,实验结果表明,丙二醛浓度较低时,氨基酸侧链上的氨基基团未受到大范围攻击,丙二醛浓度上升到1.00 mmol/L,游离氨基的减少可能是因为丙二醛分子中的羰基可直接与氨基酸侧链的氨基发生共价交联反应,进而降低了肌原纤维蛋白的游离氨基含量。
2.5 丙二醛对白鲢肌原纤维蛋白表面疏水性的影响
表面疏水性的变化反映了蛋白质构象的变化,蛋白质中非极性氨基酸暴露会导致表面疏水性增强[27]。如图5所示,随着丙二醛浓度的增加,肌原纤维蛋白的表面疏水性显著升高(P<0.05),表明丙二醛诱导的蛋白质氧化可以促进蛋白质分子结构的展开,蛋白质内部的大量非极性位点暴露于蛋白表面,导致表面疏水性的上升,这与活性巯基含量上升的结果一致(图3)。疏水基团的暴露是蛋白形成聚集体的前提,可促进蛋白质分子分子间的物理和化学交联,从而有利于鱼糜凝胶形成能力的增强[28-29]。
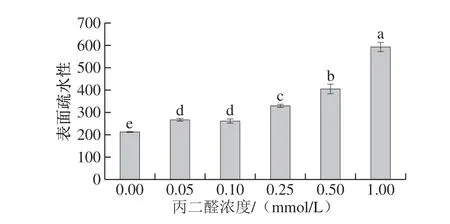
图 5 丙二醛浓度对肌原纤维蛋白表面疏水性的影响Fig. 5 Effect of MDA concentration on surface hydrophobicity of myofibrillar protein from silver carp
2.6 丙二醛对白鲢肌原纤维蛋白Ca2+-ATPase活性的影响

图 6 丙二醛浓度对肌原纤维蛋白Ca2+-ATPase活性的影响Fig. 6 Effect of malondialdehyde concentration on Ca2+-ATPase activity of myofibrillar protein from silver carp
如图6所示,随着丙二醛浓度的上升,Ca2+-ATPase活性呈先下降后上升的趋势。丙二醛浓度达到0.10 mmol/L 时,Ca2+-ATPase活性由0.056 7 μmol/(mg·min)降至0.042 5 μmol/(mg·min),丙二醛浓度增加到0.25 mmol/L, Ca2+-ATPase有所升高。肌球蛋白的头部S1区的2 个活性巯基SH1和SH2与Ca2+-ATPase的活性密切相关,适度的蛋白氧化可提高Ca2+-ATPase的活性[26,30]。当丙二醛浓度较低时(≤0.10 mmol/L),肌球蛋白头部活性位点的巯基基团遭到了氧化攻击,导致Ca2+-ATPase的活性下降;丙二醛浓度上升至0.25~1.00 mmol/L范围,Ca2+-ATPase的活性升高,推测可能是因为蛋白氧化造成了肌球蛋白头部S1区域的结构发生了显著变化,导致酶活上升,但具体原因还需进行进一步研究。
2.7 丙二醛对白鲢肌原纤维蛋白溶解度的影响
如图7所示,随着丙二醛浓度的上升,蛋白溶解度呈现下降的变化趋势,对照组的蛋白溶解度为94.76%,丙二醛浓度上升到1.00 mmol/L后,蛋白溶解度下降至47.75%。蛋白质-蛋白质和蛋白质-水之间的相互作用会影响蛋白质的溶解度[2]。蛋白溶解度的下降可能是因为随着丙二醛浓度增加,蛋白氧化程度增加,导致了表面疏水性的增加(图5),产生了聚集体,进而造成蛋白溶解度的降低。
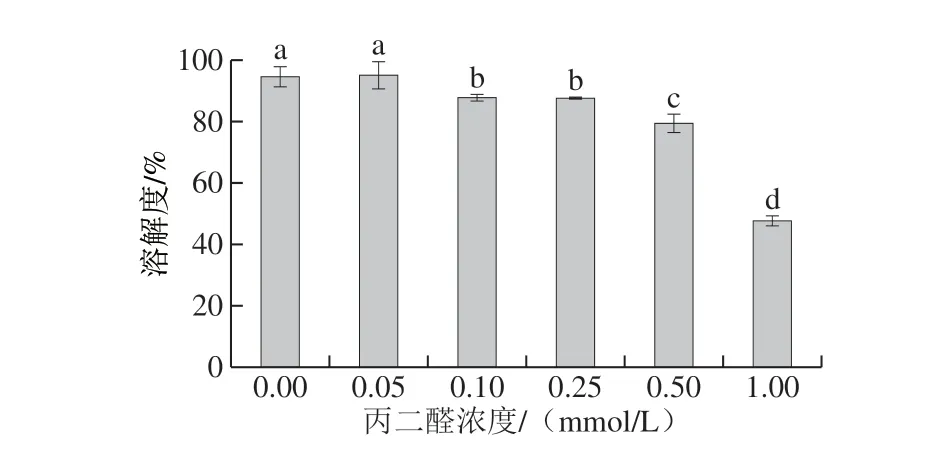
图 7 丙二醛浓度对肌原纤维蛋白溶解度的影响Fig. 7 Effect of MDA concentration on solubility of myofibrillar protein from silver carp
2.8 SDS-PAGE分析
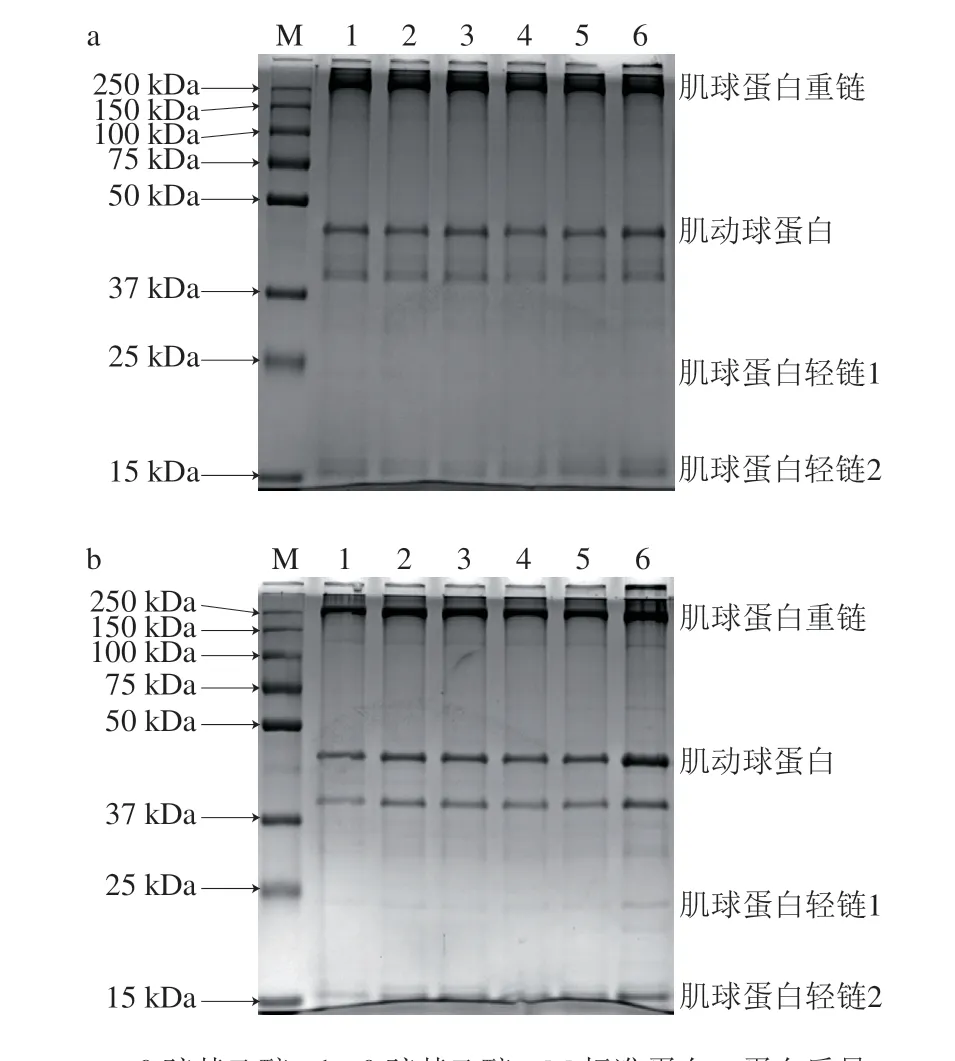
图 8 经丙二醛氧化修饰后的肌原纤维蛋白的SDS-PAGE图谱Fig. 8 SDS-PAGE pattern of silver carp myofibrillar protein after oxidative modification with MDA
如图8a所示,肌球蛋白重链是肌原纤维蛋白的主要片段[31],此外还有少量的肌动球蛋白和肌球蛋白轻链。随着丙二醛浓度的增加,在图8b中肌球蛋白重链条带强度没有明显降低且没有产生新的可见条带,这可能表示蛋白的氧化程度较轻,未能引起肌球蛋白重链的断裂。
由图8a可以看出,经过丙二醛氧化修饰后,肌球蛋白重链上方区域产生了少量高分子化合物,并有部分堆积在进样口无法进入浓缩胶,且随着丙二醛浓度的升高条带逐渐变深,表明蛋白质经氧化修饰会发生交联或聚集形成聚集体。β-巯基乙醇是一种还原剂,能够打断二硫键。从图8b可以看出,添加了β-巯基乙醇后,肌球蛋白重链上方区域的条带消失,表明巯基氧化形成了二硫键,且二硫键是主要的交联方式,但丙二醛浓度为1.00 mmol/L的还原性电泳图谱中进样口顶部条带并未完全消失,说明形成的聚合物有部分未被还原,存在无法被β-巯基乙醇打断的非二硫共价键,如酪氨酸残基之间的共价交联以及活性羰基-氨基等[32]。
2.9 丙二醛对白鲢肌原纤维蛋白二级结构的影响

图 9 丙二醛浓度对白鲢肌原纤维蛋白二级结构的影响Fig. 9 Effect of MDA concentration on secondary structure content of myofibrillar protein from silver carp
由图9可以看出,随着丙二醛浓度的增加,白鲢肌原纤维蛋白的α-螺旋和β-转角相对含量逐渐减少,无规卷曲相对含量略有下降,β-折叠相对含量逐渐增多。α-螺旋为位于多肽链内部的紧密、不含空腔的稳定结构,β-折叠和无规卷曲结构的紧密程度和构象稳定性弱于α-螺旋[33]。 蛋白质经过氧化后,α-螺旋逐渐解旋,且部分转化更为松散和舒展的β-折叠和无规卷曲结构,包埋于分子内部的疏水性残基暴露出来,活性巯基含量和表面疏水性上升(图3和图5),肌球蛋白分子间的疏水相互作用增强,引起蛋白质的聚合。
3 结 论
白鲢肌原纤维蛋白经过丙二醛氧化修饰后羰基含量显著上升,羰基和游离氨基含量在丙二醛浓度范围0.05~0.50 mmol/L内量效关系不明显,变化不显著,此浓度范围内酪氨酸单体未发生交联聚合,表明肌原纤维蛋白未发生过度氧化。丙二醛浓度低于0.25 mmol/L时,总巯基和活性巯基含量变化不明显,Ca2+-ATPase活性降低,表面疏水性升高;浓度达到0.25 mmol/L时,活性巯基含量还有Ca2+-ATPase活性显著上升,蛋白溶解度随着丙二醛浓度上升而下降。SDS-PAGE图谱显示,氧化使肌原纤维蛋白发生了交联或聚合,产生了分子质量大于250 kDa的聚集体,聚集体的主要交联方式为二硫键,但丙二醛浓度高于1 mmol/L时形成的聚集体中存在其他的交联方式。随着丙二醛浓度上升,蛋白α-螺旋相对含量减少,β-折叠相对含量增多,表明蛋白质结构逐渐伸展,原本包埋的活性巯基暴露出来。以丙二醛为代表的脂肪氧化次级产物在适宜的浓度范围内引起的肌原纤维蛋白氧化程度较为温和,能够促进蛋白质结构伸展,暴露更多活性位点,有助于蛋白质之间的交联,可能对鱼糜凝胶的形成有促进作用。
