顶复门原虫γ-氨基丁酸代谢旁路研究进展
2020-04-02胡俊菁戚南山吴彩艳吕敏娜林栩慧于林增张健騑廖申权孙铭飞
胡俊菁,戚南山,吴彩艳,李 娟,吕敏娜,林栩慧,于林增,张健騑,廖申权,孙铭飞
(广东省农业科学院动物卫生研究所 广东省畜禽疫病防治研究重点实验室 农业农村部兽用药物与诊断技术广东科学观测实验站,广州 510640)
顶复门原虫是一大类细胞内寄生原虫,包括疟原虫(Plasmodiumspp.)、刚地弓形虫(Toxoplasma gondii)、隐孢子虫(Cryptosporidiumspp.)、巴贝斯虫(Babesiaspp.)和艾美耳球虫(Eimeriaspp.)等多种重要病原体[1]。能量代谢是寄生原虫生存所必须,顶复门原虫在不同生活阶段其寄生环境有所不同,获取能量底物的种类、数量及能量代谢方式也存在差异[2]。胞内寄生原虫可以利用多种途径获得能量,其内生发育阶段主要处于低氧或无氧环境,糖酵解(glycolysis)是获取能量的主要途径之一,其次是利用多个途径的代谢产物进入三羧酸循环(tricarboxylic acid cycle,TCA)产生ATP[3-5]。其中γ-氨基丁酸代谢旁路(γ-aminobutyric acid shunt,GABA shunt)作为TCA循环的补充途径,为弓形虫和疟原虫的生长、发育和繁殖提供能量储备及部分碳骨架,具有重要的生物学意义[6-8]。本文对顶复门原虫γ-氨基丁酸代谢途径及其生物学作用进行综述。
1 γ-氨基丁酸代谢旁路的研究
1.1 γ-氨基丁酸代谢旁路 γ-氨基丁酸是一种谷氨酸衍生的氨基酸,存在于动物、植物和微生物等各种生物中,可通过γ-氨基丁酸代谢旁路代谢为琥珀酸进入TCA循环,也可转化为α-酮戊二酸进入TCA循环[9-10]。典型的γ-氨基丁酸代谢旁路是由谷氨酸脱羧酶(glutamate decarboxylase,GAD)、γ-氨基丁酸转氨酶(GABA transaminase,GABA-T)和琥珀酸半醛脱氢酶(succinic semialdehyde dehydrogenase,SSDH)三种重要酶组成。其代谢过程为:谷氨酸在GAD催化下生成γ-氨基丁酸(γ-aminobutyric acid,GABA);γ-氨基丁酸在GABA-T催化下,与丙酮酸发生转氨作用生成琥珀酸半醛和丙氨酸;琥珀酸半醛在SSDH催化下氧化生成琥珀酸进入TCA循环。虫体利用底物谷氨酸生成琥珀酸的代谢路径构成TCA循环的一个侧支,被称为γ-氨基丁酸代谢旁路。
1.2 γ-氨基丁酸代谢旁路的生理功能 γ-氨基丁酸是一种广泛存在于原核和真核生物的四碳非蛋白质氨基酸。γ-氨基丁酸通过γ-氨基丁酸代谢旁路参与大多数生物的重要代谢过程,在不同生物体中发挥不同的生理作用。血吸虫具有发达的神经系统,含有大量的神经递质,比如γ-氨基丁酸可作为神经系统的一种重要的抑制性神经递质[11]。寄生原虫靶向γ-氨基丁酸活性的化学治疗药物通过结合神经细胞或肌肉细胞的谷氨酸门控氯离子通道而发挥作用,而宿主的GABA受体仅存在于中枢神经系统,该系统有血脑屏障保护,因此此类治疗药物对哺乳动物宿主相对安全[12-13]。近年来,研究发现GABA广泛存在于细菌、真菌、植物及哺乳动物中,因病原生物中GABA受体存在的位置与哺乳动物宿主存在显著的差异,针对GABA的药物被推荐用于治疗感染性疾病[14]。γ-氨基丁酸参与微生物氮源和碳源的代谢,以及腐胺和精胺等多胺的代谢,并能够对多种逆境胁迫产生应答,具有重要的生理功能[15]。植物中γ-氨基丁酸代谢旁路参与调节植物体内pH值、植物生长发育、碳/氮源营养平衡、逆境胁迫响应和信号传递等,植物γ-氨基丁酸在多种逆境条件(缺氧、冷害、机械刺激、干旱、盐胁迫)大量积累,发挥多种生物学作用[16-17]。对顶复门原虫如弓形虫和疟原虫的研究发现,γ-氨基丁酸代谢旁路在虫体入侵宿主细胞及有性生殖阶段发挥重要的生物学作用[6-7,18]。
2 顶复门原虫γ-氨基丁酸代谢旁路
顶复门原虫可以利用谷氨酸作为碳源,为虫体提供能量或中间代谢产物。最近的研究表明,顶复门弓形虫具有完整的γ-氨基丁酸代谢旁路,谷氨酸经代谢转化为琥珀酸进入TCA循环[7,9];顶复门疟原虫具有部分的γ-氨基丁酸代谢,因缺乏琥珀酸半醛脱氢酶而不具备完整的γ-氨基丁酸代谢旁路,谷氨酸经代谢转化为α-酮戊二酸进入TCA循环[19-22]。
2.1 弓形虫的γ-氨基丁酸代谢旁路 研究表明弓形虫具有完整的TCA循环,TCA循环和β-氧化是弓形虫速殖子阶段获取能量的主要方式之一[7,9]。构建弓形虫琥珀酸合成酶缺失株,发现缺失株速殖子的生长受到一定抑制。但谷氨酸代谢对速殖子的生长发育具有同样重要的作用,推测γ-氨基丁酸代谢旁路主要为速殖子的快速繁殖、胞外溢出及入侵新的宿主细胞提供能量。研究进一步证实,这一时期弓形虫显著的减少用于形成膜脂的脂肪酸合成,减少对能量的消耗,以维持虫体的增殖与入侵[23]。
弓形虫利用γ-氨基丁酸代谢旁路将谷氨酸转为γ-氨基丁酸,再经转氨酶(SSDH)作用生成琥珀酸半醛。SSDH还可以利用丙酮酸或α-酮戊二酸作为底物进行转氨作用(图1)[1]。Blume等[24]发现弓形虫的葡萄糖转运蛋白缺失株在谷氨酸的存在条件下,可恢复滑行运动能力,证实虫体可以利用谷氨酸代谢适应葡萄糖饥饿。抑制TCA循环可抑制速殖子的复制;而缺乏γ-氨基丁酸代谢旁路的虫体则表现出生长衰退,因营养不足而无法维持活力的现象,表明GABA可作为短期的能量储备[7,25]。TCA循环对于弓形虫在宿主细胞内的复制是必要的,而γ-氨基丁酸代谢旁路则是能量的储备,为弓形虫的发育繁殖提供必需的能量需求。
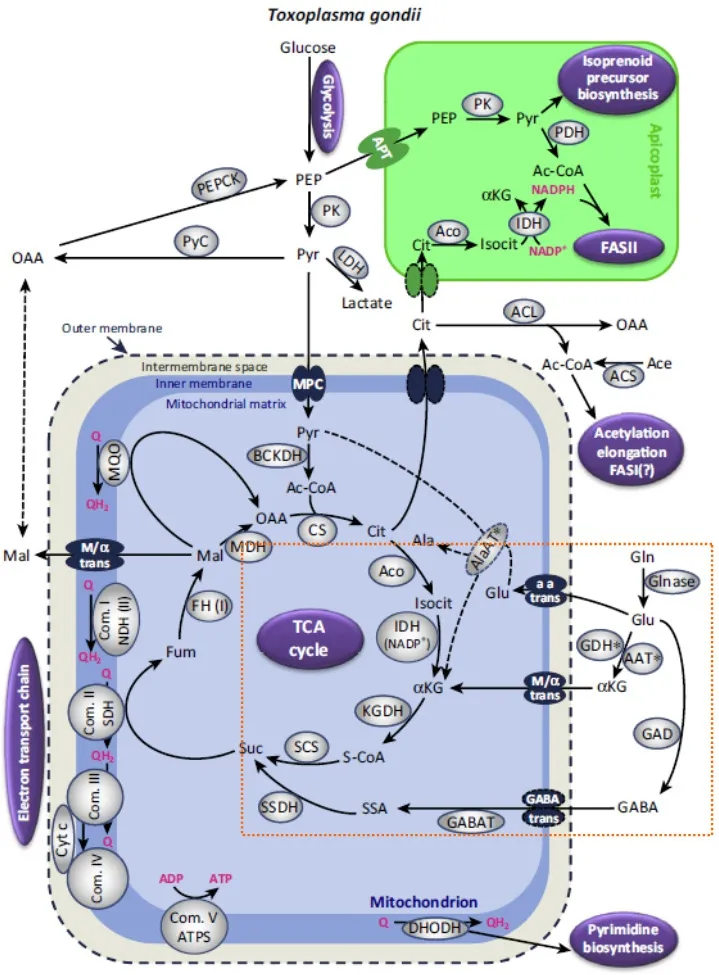
图1 弓形虫γ-氨基丁酸代谢旁路模式图[1]Fig.1 The schematic representation of GABA shunt in Toxoplasma gondii[1]
2.2 疟原虫的γ-氨基丁酸代谢旁路 疟原虫无性生殖期主要利用糖酵解途径获取能量,将葡萄糖降解为乳酸,仅有少量的丙酮酸经支链酮酸脱氢酶(branched-chain Ketoacid Dehydrogenase,BCKDH)转化为乙酰辅酶A进入TCA循环[19]。丙酮酸进入TCA循环受谷氨酸代谢生成的α-酮戊二酸调控。因疟原虫缺失的琥珀酸半醛脱氢酶使得其γ-氨基丁酸代谢与弓形虫的不同,γ-氨基丁酸转化为α-酮戊二酸直接参与TCA循环(图2)[19]。研究表明γ-氨基丁酸在疟原虫的有氧代谢中起到重要作用,可调控TCA循环。
疟原虫在配子生殖阶段的能量获得方式发生改变,以有氧代谢的TCA循环作为其获取能量的主要方式[26-27]。此时,糖酵解途径生成的丙酮酸与乙酰辅酶A是TCA循环的主要底物,这一现象表明在蚊子体内有性生殖阶段需要大量的ATP以维持其发育。研究证实配子生殖期参与TCA循环的代谢酶上调,同时抑制这些代谢酶的活性可影响疟原虫的生长发育[18-19,28-29]。虽然TCA循环在无性繁殖期并不是必须的,但有性繁殖期TCA循环发挥重要的生物学作用,抑制BCKDH活性可显著增强γ-氨基丁酸代谢通量。
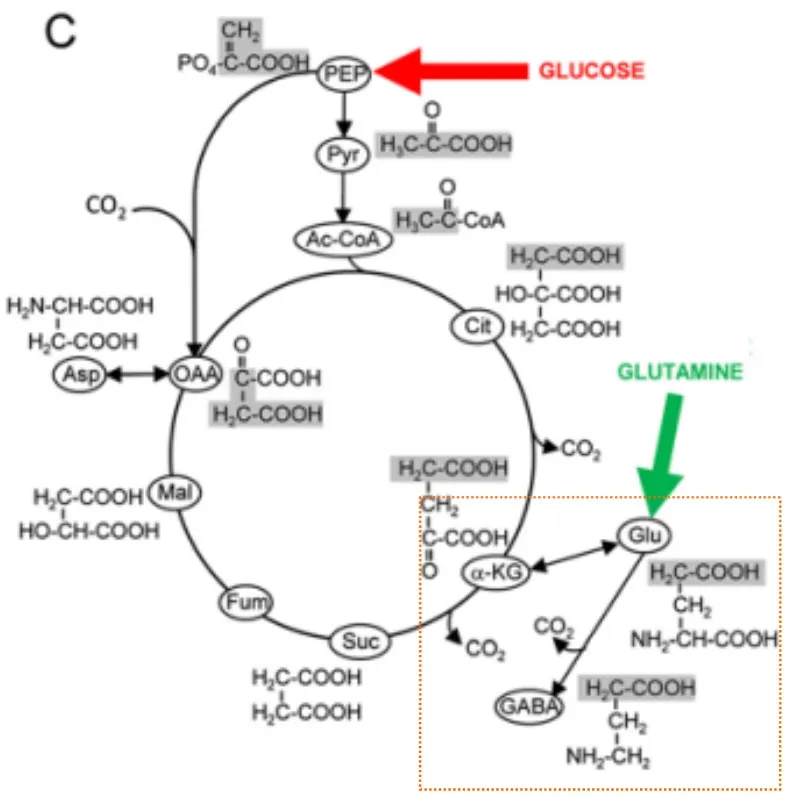
图2 疟原虫γ-氨基丁酸代谢模式图[19]Fig.2 The schematic representation of GABA metabolism in Plasmodium falciparum[19]
2.3 其他原虫的γ-氨基丁酸代谢旁路 近年来,研究表明γ-氨基丁酸代谢广泛存在于原生动物中。枯氏锥虫(Trypanosoma cruzi)的生活史复杂,包括脊椎动物、哺乳动物宿主和昆虫载体寄生阶段。因哺乳动物胞内与昆虫肠道的内环境具有较大差异,虫体进化了一套能量代谢方式以适应内环境变化。在葡萄糖存在情况下,上鞭毛体利用糖酵解获得能量;而当葡萄糖缺失情况下,虫体利用氨基酸代谢获得能量[30-31]。此外,氨基酸代谢在枯氏锥虫生长发育过程中起着多种作用,包括合成蛋白质和提供能量[32]。研究发现枯氏锥虫在能量底物缺乏时,可以利用谷氨酸、谷氨酰胺及脯氨酸产生ATP,同时谷氨酸可以由谷氨酰胺、脯氨酸或组氨酸转化提供,并且谷氨酰胺的吸收利用受谷氨酰胺合成酶的调控[33-34],而γ-氨基丁酸的吸收利用是通过至今为止发现的唯一一个Na+依赖氨基酸转运载体实现的[35]。枯氏锥虫的γ-氨基丁酸代谢类似疟原虫,γ-氨基丁酸转化为α-酮戊二酸的形式参与TCA循环,产生ATP。因此,在虫体处于能量底物缺乏时,γ-氨基丁酸代谢对于维持虫体的致病力及能量供应具有重要意义。
3 展望
近年来,γ-氨基丁酸代谢旁路在寄生虫,尤其是顶复门原虫中的生物学作用研究受到越来越多的关注。在弓形虫和疟原虫中的研究取得了突破性进展,γ-氨基丁酸代谢旁路不仅为虫体提供能量储备,并且对虫体的生长繁殖、致病力及适应不同寄生环境等起到重要作用,推测可从该旁路中的关键酶为生物靶点着手,寻找防治顶复门原虫病的新方法。研究表明,该代谢途径中的关键酶GAD是潜在的杀虫、杀螨药物靶标,L-苹果酸是其抑制剂;GABA-T是哺乳动物中枢神经系统中的重要药物靶点,用对羟基苯甲醛抑制GABA-T活性对治疗癫痫病表现出了良好的效果,香草醛和原儿茶酸等均可有效抑制GABA-T的活性[36]。野黄芩苷元是SSDH的有效抑制剂。现有的GAD、GABA-T和SSDH有效抑制剂的实验数据为证明它们在顶复门原虫上是否有成为药物靶标的可能性提供了有效参考。目前对顶复门其他寄生原虫γ-氨基丁酸代谢的研究仅限于在基因组数据中,发现和缓艾美耳球虫(E. mitis)和犬新孢子虫(Neospora caninum)γ-氨基丁酸转运载体基因序列[37],而对于γ-氨基丁酸代谢在其他原虫的代谢途径、生理意义、调控机制等仍不清楚,仍需要在生化、分子和细胞水平上对顶复门原虫GABA代谢途径进行深入研究,为寄生性原虫疾病的防治奠定一定的理论基础。
