乙烯信号参与水杨酸提高采后李果实抗冷性的研究
2020-04-01王雅楠张一冉韩育梅
王雅楠,张一冉,杨 杨,韩育梅
(内蒙古农业大学食品科学与工程学院,内蒙古呼和浩特 010018)
李子是具有重要经济价值的核果类果实,采后呼吸强度高,后熟速度快,常温贮藏条件下极易发生腐烂变质,难于长期贮藏[1]。低温贮藏可降低果实的呼吸作用而减少贮藏期间的物质消耗,推迟果实呼吸跃变,能有效延长果实的贮藏期[2]。但李子又属于冷敏型果实,低温适应性差,贮藏温度低于7 ℃时便会发生冷害,表现出果肉褐变、风味劣变、腐烂程度加剧等症状致使果实品质降低甚至丧失商品价值[3-4]。冷害现象严重制约了低温贮藏在李果实采后保鲜中的应用,冷害防治在李果实保鲜贮藏实践中至关重要。
水杨酸(salicylic acid,SA)是高等植物体内普遍存在的一种结构简单的酚类物质,具有信号分子的作用,其激活植物系统获得性抗性的功能已被熟知,近年来又发现SA可以增强植物对低温的耐受性,如果实的抗冷能力[5-6]。LUO等[7]研究发现SA能够抑制低温贮藏期间李果实的冷害发生,延缓内部果肉褐变、表皮坏死凹陷、风味劣变等冷害症状。但其引发抗冷效应的关键机制及其与其他抗冷信号分子之间的关系尚不清晰,探明这些问题对于阐释SA增加采后果实低温耐受性的机制十分必要。乙烯是调控呼吸跃变型果实采后成熟衰老的重要植物激素,在果实采后抗逆信号网络中占有重要的地位,陈鑫瑶等[8]研究发现乙烯受体抑制剂1-甲基环丙烯(1-methylcyclopropene,1-MCP)处理加剧了(2±1) ℃下采后番茄果实的冷害症状。此外一些研究也发现乙烯可能参与了SA所调节的信号过程,李翠丹等[9]研究结果发现乙烯参与了SA诱导采后番茄果实的抗病过程,LIU等[10]报道了拟南芥的抗冷诱导过程中SA和乙烯存在互作关系,但在采后果实的冷防御信号诱导过程中SA和乙烯信号之间的关系还有待加以明确。
因此,本试验以中熟期“布朗”李果实为材料,分别对其采用1.0 mmol/L SA、1.0 μL/L 1-MCP和1.0 mmol/L SA结合1.0 μL/L 1-MCP处理,以蒸馏水处理作为对照,通过研究其对冷害和活性氧代谢的影响,探讨SA诱导采后李果实抗冷能力产生的机制以及该过程与乙烯信号的关系。
1 材料与方法
1.1 材料与仪器
供试李子 采自于呼和浩特市土左旗果园,品种为“布朗”,成熟度中等,果实颜色紫红色着色面积占2/3,香气明显,手感较软。采后立即运回实验室,选择形状、大小一致,成熟度、色泽基本相同,无病虫害和机械损伤的果实进行试验;SA 天津市风船化学试剂科技有限公司;1-MCP 陕西秦丰农化有限公司;硫代巴比妥酸、愈创木酚 国药集团化学试剂有限公司;过氧化氢 天津市永晟精化工有限公司;其他均为国产试剂分析纯。
TGL-16M高速冷冻离心机 上海卢湘离心机仪器有限公司;T6新世纪型紫外分光光度计 北京普析通用仪器有限公司;DDSJ-318电导率仪 上海仪电科学仪器股份有限公司;电热恒温水浴锅 北京长安科学仪器厂。
1.2 实验方法
1.2.1 材料处理 将果实随机等分为四组,SA处理组:用1.0 mmol/L SA浸泡果实30 min;1-MCP处理组:用蒸馏水浸泡30 min后,晾干将果实进行1.0 μL/L 1-MCP熏蒸处理30 min;SA+1-MCP处理组:用1.0 mmol/L SA浸泡果实30 min,再用1.0 μL/L 1-MCP熏蒸处理30 min;CK组:用蒸馏水浸泡30 min为空白对照。将处理后的李果实自然晾干贮藏于4 ℃,85%~90%相对湿度冷库。每组处理 180个果,每6 d记录冷害发生情况并取样测定各种生理指标,每组每次取样30 个果实。将果肉切成约 0.5 cm×0.5 cm×0.5 cm 的小块,立即用液氮速冻,放入-80 ℃超低温冰箱储存并用于相关指标的测定。
1.2.2 指标测定
1.2.2.1 冷害发生率和冷害指数测定 参考陈鑫瑶[8]的方法,以单个果实冷害程度达1级及以上计为冷害果,统计冷害果占总果数的百分率。按照下列公式计算果实的冷害发生率:

参考LUO[7]、陈鑫瑶[8]的方法,通过沿着赤道轴将每个果实切成两半并通过褐变程度评估果实冷害的症状。根据果面的褐变发生面积分为5级:0级—无冷害;1级—果肉褐变占果面面积的0~25%;2级—果肉褐变占果面面积的26%~50%;3级—果肉褐变占果面面积的51%~75%;4级—果肉褐变占果面面积的76%~100%。按照下列公式计算果实的冷害指数:

1.2.2.2 过氧化物酶(POD)和多酚氧化酶(PPO)活性的测定 POD活性测定:参考曹建康等[11]方法并稍作修改,提取体系为0.1 mol/L、pH6.6的柠檬酸—柠檬酸钠缓冲液(内含 4%(w/v)PVPP,1%(v/v)Triton X-100)。反应体系是25 mmol/L愈创木酚溶液。在反应体系中加入0.5 mL酶提取液,均匀混合后,再加入200 μL 0.5 mol/L H2O2溶液迅速混合启动反应,测定5 min内470 nm波长下吸光度值的变化。以每克鲜重样品每分钟吸光度变化值增加1为1个过氧化物酶活单位,结果以U/g FW表示。
PPO活性测定:参考曹建康等[11]的方法,提取体系为0.1 mol/L、pH=5.5 的乙酸—乙酸钠提取缓冲液(内含 1 mmol/L 聚乙二醇6000,4%(w/v)PVPP,1%(v/v)Triton X-100)。反应体系是50 mmol/L、pH=5.5的乙酸—乙酸钠缓冲液和50 mmol/L 的邻苯二酚溶液,最后加入 100 μL 酶提取液混合启动反应,测定5 min内420 nm波长下吸光度值的变化。以每克鲜重样品每分钟吸光度变化值增加1为1个多酚氧化酶活单位,结果以U/g FW表示。
1.2.2.3 细胞膜渗透率和丙二醛(MDA)含量测定 细胞膜渗透率:参考曹建康等[11]的方法。
丙二醛含量:采用硫代巴比妥酸方法,略有改进。提取体系为100 g/L TCA溶液[7,12]。反应体系为酶提取液、0.6%(w/v)TBA(由100 g/L TCA配成),混合后在95 ℃水浴20 min,迅速冷却并以10000×g离心10 min。再分别测定上清液在450、532和600 nm波长处的吸光度值,并计算MDA的含量。结果以nmol/g FW表示。
1.2.2.4 超氧化物歧化酶(SOD)和过氧化氢酶(CAT)活性的测定 SOD活性测定:参考丁波等[13]的方法。提取体系为0.1 mol/L、pH=7.8 的磷酸提取缓冲液(内含 5 mmol/L 二硫苏糖醇和 5%(w/v)PVP)。反应体系是50 mmol/L、pH=7.8 磷酸缓冲液(内含13 mmol/L甲硫氨酸、75 μmol/L氮蓝四唑、10 μmol/L EDTA-Na2和2 μmol/L核黄素),在反应体系中加入0.1 mL酶提取液,光照25 min后,在560 nm波长比色,以每分钟每克鲜重样品的反应体系对氮蓝四唑光化还原的抑制为50%为一个SOD 活性单位(U)。结果以U/g FW表示。
CAT活性测定:参考曹建康等[11]方法并加以修改。提取体系为0.2 mol/L、pH7.9的磷酸钠缓冲液(内含1%(v/v)Triton X-100)。反应体系是酶提取液、0.2 mol/L、pH7.9的磷酸钠缓冲液、蒸馏水,均匀混合后,将混合液置于30 ℃下恒温反应30 min,再加入300 μL 0.1 mol/L H2O2溶液迅速混合启动反应,测定样品在波长240 nm处每隔30 s吸光度值的变化。以每克鲜重样品每分钟吸光度变化值减少0.01为1个过氧化氢酶活单位,结果以U/g FW表示。
1.3 数据处理
以上试验测定重复三次,各项指标测定值均取三个平行值进行数据分析。采用Excel 2007的STDEV与SPSS 20.0的ANOVA进行统计处理。所有图表采用Orgin9.4绘制。
2 结果与分析
2.1 不同处理对李果实低温贮藏期间冷害发生率和冷害指数的影响
由图1A可知,李果实低温贮藏第6 d各处理组均不同程度出现冷害症状,对照组、SA处理组、1-MCP处理组和SA结合1-MCP处理组果实冷害发生率依次为10.00%、3.33%、13.33%和6.67%,冷害指数依次为:0.025、0.008、0.042和0.017,1-MCP处理组冷害发生率和冷害指数均显著高于对照组(P<0.05),SA处理和SA结合1-MCP处理组冷害发生率和冷害指数均显著低于对照组(P<0.05);随着贮藏时间的延长各处理组果实冷害发生率冷害指数均逐渐增加,到第24 d时1-MCP处理的冷害发生率为83.33%,冷害指数为0.733;SA处理组冷害发生率为50%,冷害指数为0.400;SA结合1-MCP处理组冷害发生率为63.33%,冷害指数为0.500。与对照组相比,SA处理和SA结合1-MCP处理均显著降低了贮藏第24 d时果实冷害的发生(P<0.05),其中SA处理组的冷害发生速度低于SA结合1-MCP处理组。贮藏第30 d时,各处理组果实均发生冷害现象,各处理组果实的冷害指数在贮藏结束后均达到最大,SA处理组和SA结合1-MCP处理组的冷害指数显著低于对照组(P<0.05),分别为0.917和0.967,1-MCP处理和对照组的冷害指数最高为1。这表明1-MCP处理加重了4 ℃条件下果实冷害的发生,SA和SA结合1-MCP处理均显著抑制了果实冷害的发生,但SA结合1-MCP的抑制能力低于SA单独处理。(图1A、1B)。
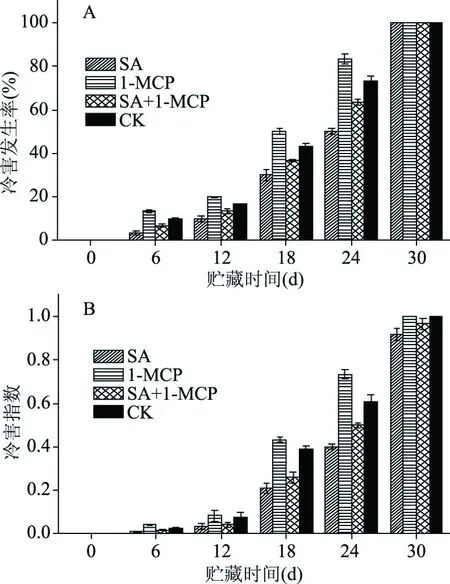
图1 不同处理对李果实4 ℃贮藏期间冷害发生率和冷害指数的影响Fig.1 Effects of different treatments on the chilling injury rate and chilling injury index of plum fruit during storage at 4 ℃
2.2 不同处理对李果实低温贮藏期间POD和PPO活性的影响
褐变是李果实冷害的典型现象,与POD和PPO所引发的酚类物质的氧化有关。由图2A可以看出,SA处理组果实POD活性高峰出现于处理后第24 d,对照李果实POD活性高峰出现于第12 d,SA处理推迟了果实POD活性上升,抑制李果实POD活性,减轻果肉褐变程度;1-MCP处理组果实POD活性于处理后第6 d达到活性最大值并显著(P<0.05)高于对照和其他处理组,随后POD活性迅速下降后上升,果肉褐变症状最先出现,在整个贮藏期间1-MCP处理组果实果肉褐变程度最为严重;SA结合1-MCP处理果实POD活性于处理后第6 d达到活性最大值,但POD活性又迅速下降,在贮藏12~18 d期间低于对照组,抑制李果实果肉褐变冷害症状。PPO活性在SA、1-MCP处理和对照果实中表现出类似的模式(图2B),综上所述,SA处理推迟了果实POD、PPO活性上升,减轻果肉褐变程度;1-MCP处理组果实POD、PPO活性于贮藏前期达到活性最大值,果肉褐变症状最先出现,随着贮藏时间的延长果肉褐变严重,从而导致贮藏后期李果实细胞坏死严重;而SA结合1-MCP处理果实在贮藏(第12~18 d)中期抑制POD、PPO等促褐变酶的活性,延缓果肉褐变等冷害现象。
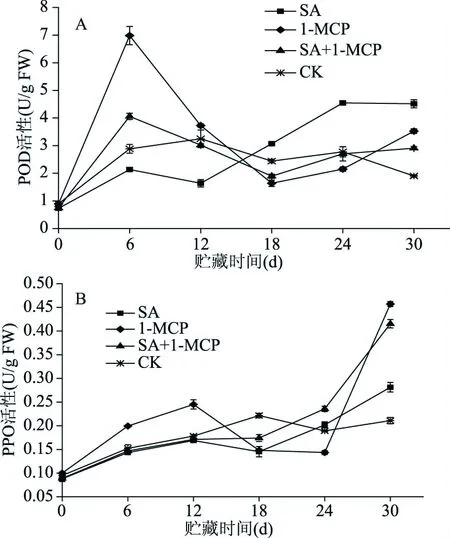
图2 不同处理对李果实4 ℃贮藏期间POD和PPO活性的影响Fig.2 Effects of different treatments on POD and PPO activities of plum fruit during storage at 4 ℃
2.3 不同处理对李果实低温贮藏期间相对电导率和MDA含量的影响

图3 不同处理对李果实4 ℃贮藏期间细胞膜渗透率和MDA含量的影响Fig.3 Effects of different treatments on cell membrane permeability and MDA content of plum fruit during storage at 4 ℃
细胞渗透率是衡量细胞膜完整性的重要指标之一,反映细胞膜损伤的程度和组织器官衰老程度。由图3A可知,对照组果实在贮藏0~24 d期间细胞膜渗透率平均上升速率为0.97%/d,而SA处理组果实相对电导率平均上升速率为0.91%/d,SA显著抑制(P<0.05)果实贮藏期间细胞膜渗透率的增加。1-MCP处理在贮藏前期第6 d达到细胞膜渗透率最大值71.49%,显著高于其他三组果实(P<0.05),并一直保持较高的水平,SA结合1-MCP处理的相对电导率变化趋势与SA处理组果实变化一致,该处理显著抑制了贮藏(第18~24 d)相对电导率的升高(P<0.05)。MDA是膜脂过氧化代谢产物,其含量大小反映细胞膜破坏的程度。图3B结果显示,SA处理组果实MDA含量在整个贮藏期间显著低于对照组(P<0.05);1-MCP处理果实MDA在贮藏前期升高较快,第6 d和第24 d时显著(P<0.05)高于对照组;SA结合1-MCP处理组在贮藏12~18 d期间MDA含量显著(P<0.05)低于对照组。SA处理对整个贮藏期李果实MDA含量的增加有抑制作用,而SA结合1-MCP处理在贮藏(第12~18 d)中期对MDA含量增加有抑制作用。
2.4 不同处理对李果实低温贮藏期间SOD和CAT活性的影响
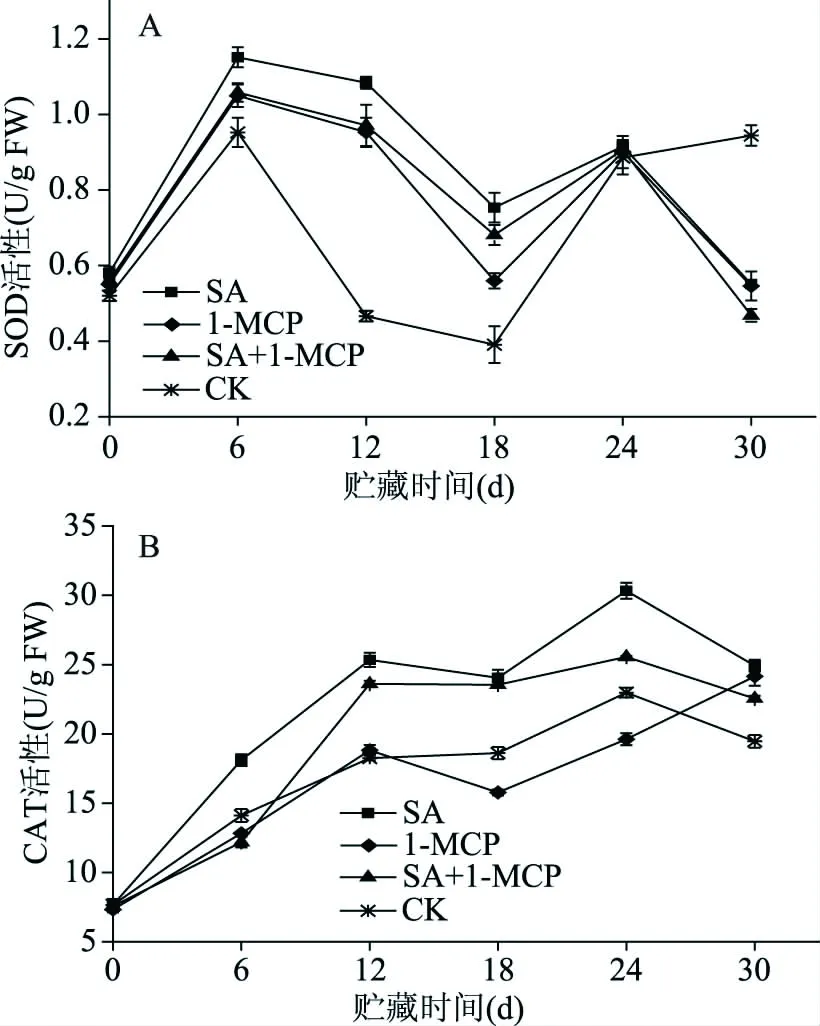
图4 不同处理对李果实4 ℃贮藏期间SOD和CAT活性的影响Fig.4 Effects of different treatments on SOD and CAT activities of plum fruit during storage at 4 ℃
SOD和CAT是生物细胞代谢酶系的重要成员,与活性氧清除密切相关。由图4A可知,随着贮藏时间的延长SOD活性总体呈先上升后下降的变化趋势。三组处理SOD活性在低温贮藏6~18 d期间均显著(P<0.05)高于对照组,其中SA处理组果实在此期间一直保持最高的SOD活性,并在第6 d达到了整个贮藏期SOD活性的最大值1.15 U/g FW,是对照组第6 dSOD活性的1.21倍。由图4B可知,在整个贮藏期间,各处理和对照组果实CAT活性总体呈上升的趋势,其中SA处理组活性上升的速度最快,并于处理后第24 d达到CAT活性最大值(30.33 U/g FW),显著(P<0.05)高于对照组和其他两组处理;1-MCP处理果实在贮藏18~24 d期间CAT活性显著(P<0.05)低于对照组。SA结合1-MCP处理组果实在贮藏12~30 d 期间一直保持较高的CAT活性,且始终显著高于对照(P<0.05)。上述结果表明,SA处理和SA结合1-MCP处理可提高SOD、CAT活性,1-MCP处理抑制了贮藏第18~24 d的CAT的活性。SA结合1-MCP处理组果实在低温贮藏期间也保持了较高的SOD、CAT活性但整体水平低于SA处理组。
3 结论与讨论

果实对低温等逆境环境的耐受性提高并非关于单一信号分子作用的结果,而是依靠一套信号调节网络实现的。在SA提高植物对生物和非生物胁迫适应能力的研究中,乙烯信号作用一直备受关注。李翠丹[9]、S. Sugano[37]、LIU等[10]在抗病、低温胁迫领域的研究表明SA和乙烯存在互作关系。本试验研究结果显示:与空白对照相比,乙烯受体竞争性抑制剂1-MCP 处理李果实随着低温贮藏时间的延长冷害症状加剧,果实POD、PPO活性于贮藏前期达到活性最大值,果肉褐变症状最先出现,从而导致贮藏后期李果实细胞坏死严重,对减轻李果实冷害没有任何明显作用,反而加剧果实冷害症状,这有可能是因为1-MCP抑制乙烯信号分子的传导与表达,进而加重李果实的冷害现象,这与陈鑫瑶[8]在番茄果实上的研究结果相似。而1-MCP处理只在贮藏前、中期提高SOD活性,对CAT活性提高起到抑制的作用,推测1-MCP处理李果实无法利用CAT将有毒性的H2O2分解为无毒的H2O和O2,进而导致李果实体内积累大量的H2O2,可能直接或间接地导致细胞膜脂质过氧化损害,细胞膜结构和功能受损严重,使贮藏前期细胞渗透率和MDA含量处在较高水平,导致果肉褐变严重,最后加剧李果实冷害的现象。因此,本试验说明阻断乙烯信号降低了李果实的抗冷能力。
SA结合1-MCP处理李果实在4 ℃低温贮藏期间冷害发生率和冷害指数均显著(P<0.05)低于对照组和1-MCP处理组,但显著(P<0.05)高于SA处理组,其处理对李果实抑制冷害现象具有一定的效果。SA结合1-MCP处理组在贮藏中期抑制POD、PPO等促褐变酶的活性,延缓果肉褐变等冷害现象,同时,在低温贮藏期间保持较高的SOD、CAT等活性氧清除酶活性,在贮藏前、中期对细胞渗透率和MDA含量增加有显著抑制作用(P<0.05),其抑制冷害症状的效果仅次于SA处理组,有效减少活性氧的积累,降低细胞膜质的氧化伤害,延缓李果实冷害的发生。上述试验结果说明,阻断乙烯信号一定程度上削弱了SA对李果实抗冷性的诱导效应,但同时SA对果实抗冷信号的调节机制中可能也存在不依赖于乙烯信号的通路。李翠丹[9]在研究SA诱导番茄果实抗病机制的结果中发现番茄果实乙烯信号阻断时削弱了SA诱抗效果,但SA对NO的诱导作用未受到明显影响,因此,推测不依赖于乙烯信号途径可能与NO有关。在SA诱导李果实产生抗冷能力的过程中是否存在相似的机制还有待于进一步研究。
综上所述,适宜浓度SA处理提高了采后中熟期“布朗”李果实的抗冷能力,该作用与处理维持高水平ROS代谢能力和抑制促褐变活性有关。乙烯信号参与了这一过程,但同时SA对李果实抗冷能力诱导不完全依赖于乙烯信号的传递。
